1. Introduction
Nowadays, it is widely accepted that incorporation of functional ingredients in aquafeeds represents an efficient farming tool that combines the desired immune modulatory properties, without neglecting the correct nutritional provisions required for fish normal physiological needs. In this context, it is of utmost importance to unveil the nutraceutical potential of tryptophan in the quest for the definition of new strategies to empower fish in the mitigation of the non-ideal production conditions. Recent works have put into evidence the immune tolerance role of tryptophan dietary supplementation in fish, displaying its potential in the self-restrain mechanisms that compose the immune and stress responses. Machado et al. [
1] showed that European seabass (
Dicentrarchus labrax) fed days a tryptophan supplemented diet for 14 and then submitted to an inflammatory insult presented a decreased cellular response and dropped plasma nitric oxide and bactericidal activity, compared to fish fed a control diet. In another study, Machado et al. [
2] showed that dietary tryptophan clearly displayed a role in the attenuation of the immune response, once again inhibiting the inflammatory mechanisms and, to some degree, decreasing seabass disease resistance to
Photobacterium damselae subsp.
piscicida (
Phdp). In fact, Machado et al.’s [
3] in vitro observations pointed to a more direct role of tryptophan in the limitation of self-damage in inflammatory scenarios by the modulation of pathways associated to the assemblage of anti-inflammatory machinery and induction of macrophages polarization towards an anti-inflammatory phenotype. Moreover, a scenario of dietary tryptophan deficiency may equally represent an handicap for the European seabass, since it may lead to a rise in plasma cortisol levels in response to infection, which may explain the observed compromised immune cell response and disease resistance to
Phdp [
2].
Tryptophan has also recognized roles in the neuroendocrine system and, therefore, works as a player in the neuroendocrine–immune axis. Lepage et al. [
4,
5] worked on the topic and observed that rainbow trout (
Oncorhynchus mykiss) fed for 1 week on a diet supplemented with tryptophan (up to 4× above the control diet) undergo different outcomes, according to their stress state, with serotonin (5-HT) acting either as a trigger or inhibitor of the hypothalamic–pituitary–interregnal axis. Taking the fact that fish showed elevated plasma cortisol levels in response to stress into account, tryptophan-supplemented feed led to a significant reduction in the stress-induced elevation of plasma cortisol. In contrast, tryptophan-supplemented feed offered to undisturbed fish resulted in a slight elevation of plasma cortisol levels. In accordance, in an attempt to explore the links between the immune and neuroendocrine responses, Azeredo et al. [
6] observed that an increase in dietary tryptophan (i.e., 4× more tryptophan than the control diet) was beneficial in chronically stressed (i.e., high stocking density) Senegalese sole (
Solea senegalensis) fed for 38 days. However, undisturbed individuals showed a compromised immune status and lower disease resistance in the latter study.
The outcomes of dietary tryptophan trust on its role as a precursor of the compounds responsible for the modulation of stress coping mechanisms and antioxidant and immune mechanisms [
7]. In macrophages, tryptophan catabolism is mediated by indoleamine 2,3-dioxygenase (IDO), which catalyzes the first and rate-limiting step of the tryptophan catabolism along the kynurenine pathway. In fact, IDO directly relies on the availability of tryptophan, and its transcription is induced by inflammatory stimuli, such as interferon-γ and cytokines [
8,
9]. Afterwards, tryptophan catabolic pathway is responsible for exerting anti-microbial effects by tryptophan depletion from the extracellular environment, thus reducing its availability to microorganisms. Moreover, during the kynurenine pathway, the formation of specific metabolites, such as 3-hydroxykynurenine, 3-hydroxyanthranilic acid, and quinolinic acid, regulate T-cell function, and the same metabolites set up a protector system, acting locally to the removal of superoxide radicals modulating oxidative status, thus creating conditions that favor immune suppression and tolerance [
8,
10]. Similarly, since both immune and stress responses share common pathways, effector organs, and signalling molecules [
11], leucocytes are also sensitive to a wide repertoire of other neuroendocrine effectors, such as glucocorticoids. Consequently, important stress signalling molecules, such as cortisol, can have profound effects on the fish immune response by influencing the balanced successive secretion of pro- and anti-inflammatory cytokines [
12,
13,
14]. Actually, tryptophan works as precursor of relevant functional molecules at the neuroendocrine level. The 5-HT is produced from tryptophan by the rate-limiting enzyme tryptophan hydroxylase [
15]. This monoamine, which can simultaneously function as a neurotransmitter in the central nervous system and a paracrine or endocrine signal in the gut and blood [
16,
17], is able to either induce or inhibit the production of adrenocorticotropic hormone (ACTH) in the pituitary. ACTH is then responsible for the increase or decrease of interrenal cortisol synthesis [
4].
Considering all previous studies, it could be inferred that the consumption needs and physiological fates of tryptophan seems to be modulated according to both the physiological status of the individuals (e.g., stress response) and the progress of inflammation. Therefore, and taking into account the bi-directional and dependent relationship of both neuro-endocrine and immune responses, dietary tryptophan supplementation may represent a tool in the mitigation of stressful conditions during current farming practices, consequently avoiding unfavorable stress-inducing immune suppressor effects upon an infection episode or an inflammatory inducer. Hence, the present work intends to study the ability of dietary tryptophan supplementation to modulate the immune response to an acute inflammatory stimulus in European seabass juveniles reared under stressful conditions (i.e., high stocking density).
3. Discussion
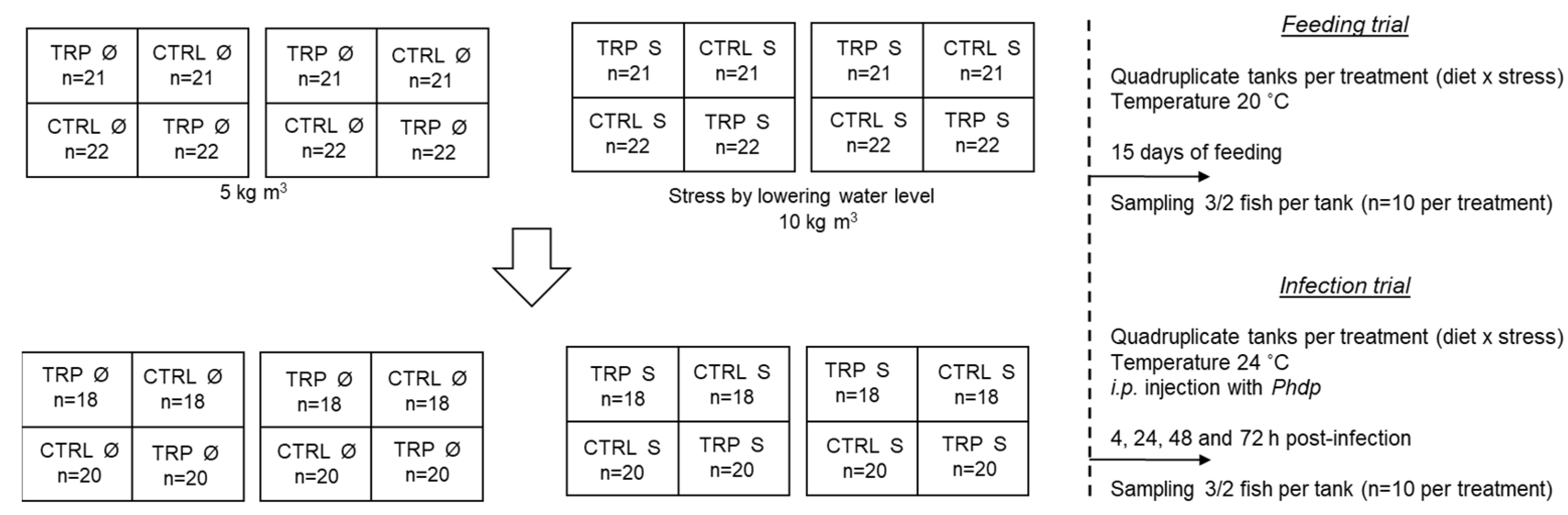
Aiming to understand the outcome of dietary tryptophan supplementation in an interactive scenario of stressful rearing conditions and an activation of inflammatory mechanisms, European seabass juveniles reared at high stocking density and fed a diet supplemented with tryptophan for 15 days were infected with Phdp.
Primarily, stress conditions alone, irrespective of dietary treatment and infection, did not significantly compromise the fish health status, with the exception of a decrease of haematocrit and
mc2r mRNA expression and an increase of the haemoglobin concentration and corpuscular indexes. The melanocortin 2 receptor (
mc2r) codes an ACTH receptor that regulates synthesis and release of glucocorticoids in response to ACTH release and, therefore, is essential for the fish stress response. However, the observed down-regulation of the gene, in response to stress, could be the result of a feedback response to the adaptation to prolonged stress [
18]. In fact, cortisol levels tended to be higher in stressed European seabass, compared to undisturbed groups at the sampling time 0 h, i.e., after 15 days exposure to high-density conditions. Therefore, as suggested by Agulleiro et al. [
18], chronic dietary cortisol may directly result in a decreased MC2R expression as an upstream control mechanism of cortisol release.
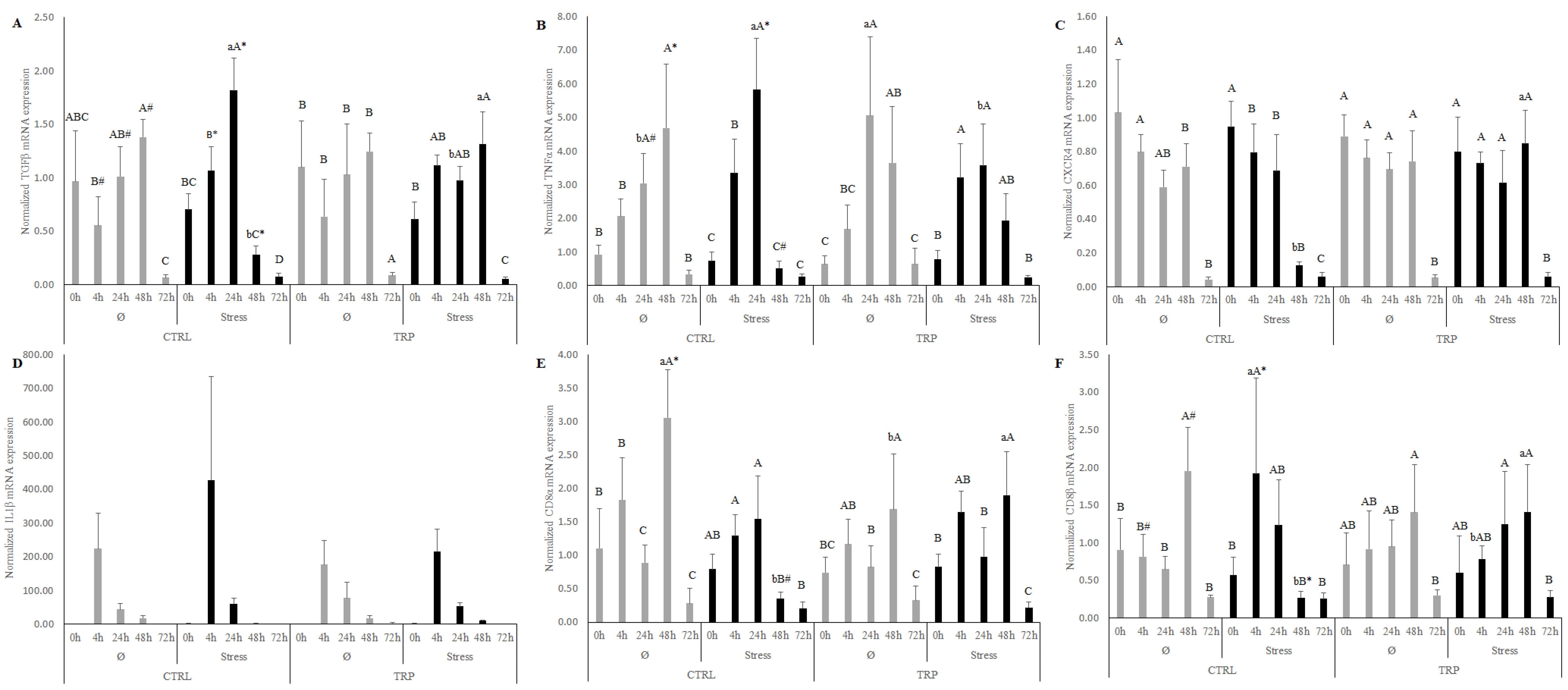
Differently, immune stimulation itself induced a clear inflammatory response, as could be expected, that was transversally characterized by a promptly enhanced neutrophil and monocyte response (4 h), followed by an increase of the concentration of total leucocytes found in the peritoneal cavity (from 24 to 48 h). In fact, plasma cortisol levels, as well as the expression of the pro-inflammatory cytokine il1β, were found to be augmented by inflammation right after infection (i.e., 4 and 24 h). Later, the inflammatory process was characterized by the up-regulation of numerous inflammatory genes at 48 h, such as the regulatory cytokine
tgfβ, the acute-phase cytokine
tnfα, both T-cell surface glycoproteins
cd8α and −
β, and T-cell receptors
cd3zeta and
tcrα. The observed modulation of the peripheral and local response to the infection with
Phdp is in line with several previous reports [
2,
19,
20,
21,
22] that characterize the acute response to inflammation as a fast neutrophilia and monocytosis within the first 24 h of infection, together with the up-regulation of pro-inflammatory genes at the systemic level. Interestingly, intestinal oxidative stress appeared diminished, as gathered by the decreased antioxidant enzymes superoxide dismutase (SOD), catalase (CAT), and total glutathione (tGSH) activity. This lower activity of antioxidant enzymes at the gut level might be the reflection of a shift in resource allocation to the inflammatory focus [
23], the peritoneal cavity, supported by a sturdy response at the peripheral (blood and head kidney genes) and local (peritoneal leucocyte) levels.
Nonetheless, it is worth mentioning that, in the present study, stressful rearing conditions seemed to exacerbate the inflammatory response. Despite the lower haemoglobin, haemoglobin corpuscular indexes, and activity of the antioxidant enzymes SOD, CAT, and tGSH in the gut, compared to non-stressed individuals, stressed fish showed a higher activity of several inflammatory indicators. Contrarily to what would be expected in chronically stressed animals (15 days exposure to high stocking density conditions), no clear innate immune suppression, due to the rise of cortisol [
7] levels, seems to have taken place. In fact, stressed seabass clearly displayed a profile resembling the one normally found in acute stressed fish [
24]. A significantly higher concentration of peripheral total leucocytes and neutrophils was found in stressed fish at the early stage of inflammation (4 and 24 h). In addition, key inflammatory genes, such as
il1β and
tgfβ (at 4 h),
tnfα (at 24 h) and
cd8α, and
-β (at 48 h), were found to be highly expressed in stressed European seabass, compared to non-stressed groups. This points to a probable enhancement of the inflammatory response by stress, both at the leucocyte and transcriptional levels. Nonetheless, stressed fish presented a significantly lower expression of
tnfα,
cd8α,
cd3zeta, and
tcrα than the undisturbed fish at a late sampling point (48 h). This may, in contrast, suggest a certain degree of immune-suppressing effect of stress, mostly on T-cell surface glycoproteins receptors. In fact, some authors have observed not only a down-regulation of the lymphocyte surface molecules essential for their activation, but also a restrain in their tissue distribution, while the same was not observed in phagocytic cells (neutrophils and monocytes/macrophages) [
25,
26]. The disparity of the observed results could be the outcome of several factors, such as the type and severity of the stressor used [
24,
27], despite the prolonged time of exposure applied in the present study (15 days).
The main goal of the present work was to assess whether dietary tryptophan supplementation during stressful conditions could be of advantage in the event of an infection.
Exclusively from tryptophan dietary supplementation, apart from a decreased plasma bactericidal activity, no further changes were observed. Such an inability of dietary tryptophan, per se, to induce significant changes in fish immune mechanisms was previously demonstrated in Persian sturgeon (
Acipenser persicus) by Hoseini et al. [
28] and in seabass [
1,
2] after a 15 days feeding period. Indeed, most of the attributed immune-related effects of dietary tryptophan were described as dependent on immune stimulation and associated with the induction of the IDO-related kynurenine pathway [
1,
2,
8,
9,
28]. In agreement, a small number of immunosuppressive effects were observed in fish fed a higher tryptophan dose in response to inflammation. Decreases of MCV, gut SOD activity, and down-regulation of the pro-inflammatory cytokine
il1β and T-cell glycoprotein
cd8α in the head kidney were observed. Nonetheless,
tnfα expression was found to be up-regulated at 24 h in response to infection. In a previous study, dietary tryptophan offered 26% above the requirement level led to a weaker peripheral and local leucocyte response, to a down-regulation of key immune-related genes, and to a concomitant decrease in disease resistance to bacterial infection (
Phdp) [
2]. Engelsma et al. [
29] also described a similar effect in common carp (
Cyprinus carpio). Despite being shy, the outcome of the inflammatory response observed in the present study is in agreement with the premise that tryptophan may prime immune suppression by promoting immune-tolerance signals during inflammation [
2,
3,
29,
30]. Upon inflammatory induction, tryptophan catabolism in macrophages through the kynurenine pathway contributes to a reduction of its availability for microorganisms consumption, deviates superoxide radicals for the modulation of cell oxidative status, and regulates T-cell function, thus creating conditions that favor immune suppression and tolerance [
8,
10]. In addition, tryptophan supplementation may contribute to immune tolerance by promoting macrophage differentiation towards the M2/healing phenotype in an inflammatory scenario [
3]. Nonetheless, no modulation of either kynurenine-related genes, such as the
afmid and
ido2, was observed in response to tryptophan supplementation in an inflammatory scenario.
In contrast, considering dietary tryptophan supplementation during stressful rearing conditions and its outcome in the inflammatory mechanisms, the present study showed an important reduction of plasma cortisol levels at 4 h of infection, compared to CTRL-fed fish. In fact, despite non-significance, tryptophan seemed to induce a reduction in basal cortisol levels in response to stress, even prior to the inflammatory stimulus (0 h). This is in agreement with the previous reports that point to the ability of tryptophan to counteract stress-induced cortisol increase [
4,
31,
32] and concomitantly avoid the cortisol immune suppressor effect [
2,
7]. This hypothesis is further supported by the up-regulation of several immune-related genes at 48 h following infection. At that sampling point, increased mRNA expressions of
tgfβ¸
cxcr4,
cd8α, and
–β,
cd3zeta and
tcrα were observed, highlighting a less restrained response. TGFβ is a multifunctional cytokine produced by leucocytes, with key roles in several cellular process, while CXCR4 is a membrane protein present in leucocytes that binds to the CXC chemokines family. Moreover, contrarily with the previously observed immune-suppressing effect at 48 h of stress, the tryptophan spare seems to revert stress-induced T-cell suppression by increasing the gene expression of the T-cell surface glycoproteins and receptors (
cd8α and
–β,
cd3zeta,
tcrα). However, it is described that only about 10% of total L-tryptophan is consumed through the neuro-endocrine routes [
33]. Indeed, in the present experimental conditions, an up-regulation of
ido2 and
afmid was visible at 48 h, pointing to the involvement of the tryptophan catabolic pathway in macrophages mediated by IDO. IDO2 is the first and a rate-limiting enzyme of tryptophan catabolism in macrophages, followed by the enzyme arylformamidase (AFMID), which is responsible for kynurenine formation. Through this route, tryptophan consumption sets up a protection system, mostly characterized by its immune suppressive effect and role during the immune response [
3,
8,
10]. Nonetheless, the down-regulation of
tnfα,
cd8β, and
tgfβ at earlier sampling points was perceived.
To conclude, and despite of the main goal of the present study, the multiplicity of factors such as stress, inflammation, and tryptophan, per se, as well as their interactions, cannot be ignored. To begin with, stressful rearing conditions alone did not significantly compromise seabass health indicators, apart from the expectation of a decreased MC2R expression, possibly as a control mechanism of the prolonged cortisol release. In contrast, immune stimulation itself induced, as expected, a clear inflammatory response at the peripheral (blood and head kidney genes) and local (peritoneal leucocyte) levels. However, no clear immune suppression was perceived when chronically stressed fish (15 days under high stocking density conditions) were submitted to an acute inflammation. Contrarily, stress seemed to exacerbate the inflammatory response at early sampling points (up to 24 h). Nonetheless, stressed fish presented some immune-supressing effects, mostly on T-cell surface glycoproteins receptors at a late sampling point (48 h).
Finally, while tryptophan alone failed to modulate seabass immune indicators, a small number of immunosuppressive effects were observed in response to inflammation. Such results are in agreement with the premise that tryptophan may prime immune suppression by promoting immune-tolerance signals during inflammation. Nonetheless, when stress is a present factor, dietary tryptophan supplementation seems to improve the imposed inflammatory response by mitigating stress effects, since a clear reduction of plasma cortisol levels was promptly observed in TRP-fed fish. This ability of tryptophan to counteract stress-induced cortisol increase is further supported by the up-regulation of several immune-related genes at 48 h of infection and by a reversal of the previously observed stress-induced T-cell suppression. Finally, the involvement of the tryptophan catabolic pathway in macrophages mediated by IDO was confirmed by the up-regulation of genes involved in the kynurenine pathway. Data from the present study could be useful for the development of novel prophylactic protocols during vaccination by intraperitoneal injection in the European seabass, since fish are crowded prior to i.p. injection, and an improved immune response during inflammation could enhance vaccination efficiency.


 ,
, 






{kind=link}
{kind=link}
{kind=link}