
Host- and Age-Dependent Transcriptional Changes in Mycobacterium tuberculosis Cell Envelope Biosynthesis Genes after Exposure to Human Alveolar Lining Fluid

 , ,
, ,
Abstract
:1. Introduction
2. Results
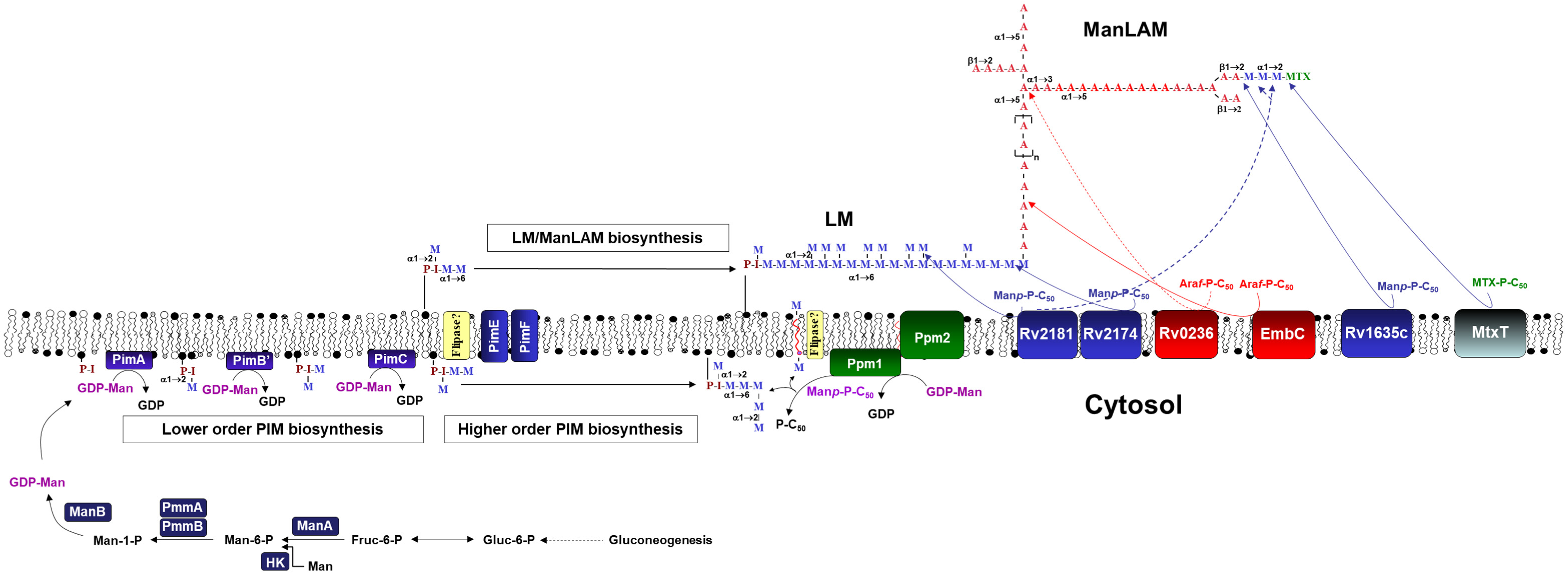
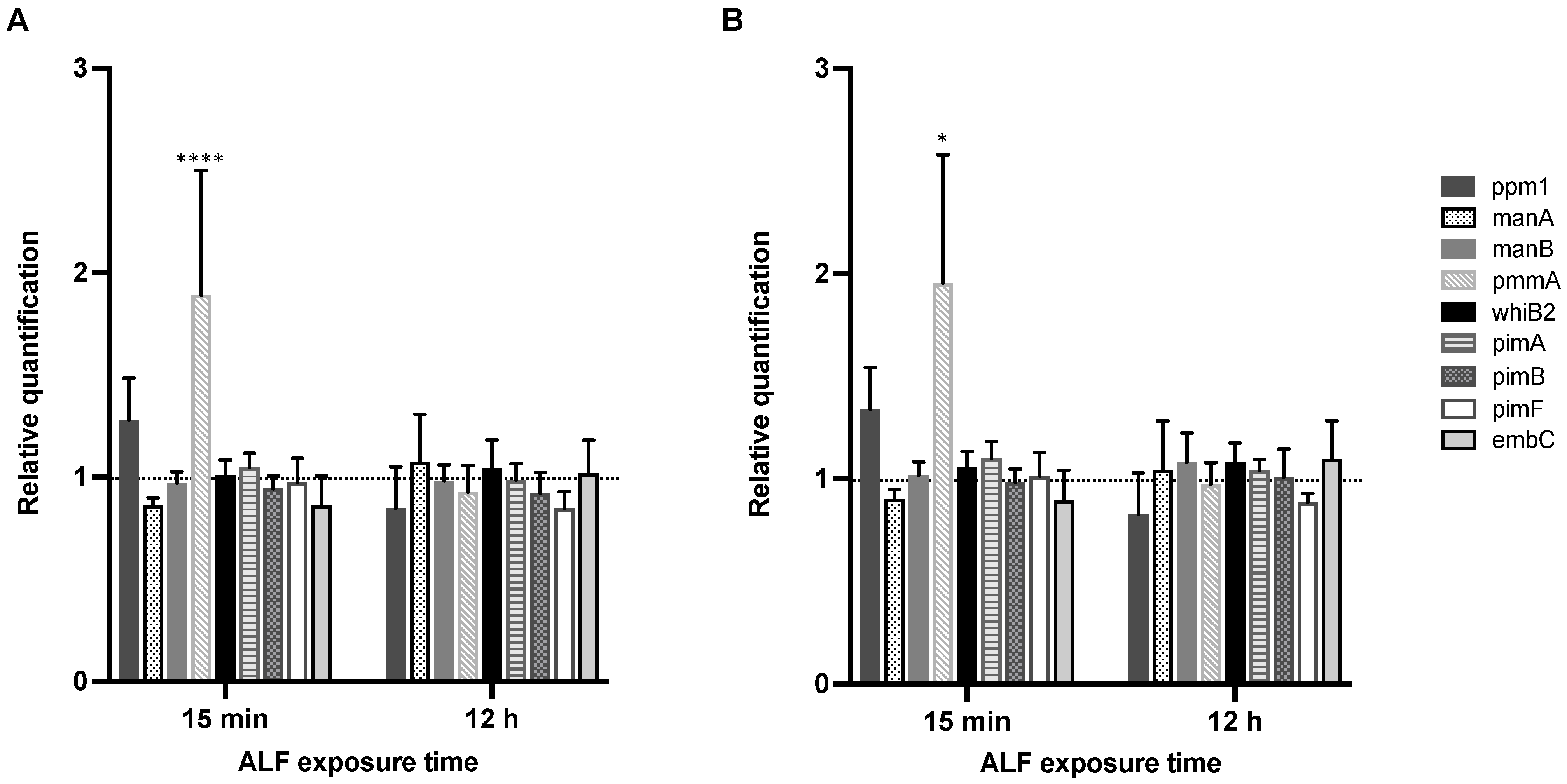
2.1. Expression of Cell Envelope PIMs/LM/ManLAM Biosynthesis Genes in M.tb after Exposure to Human ALF
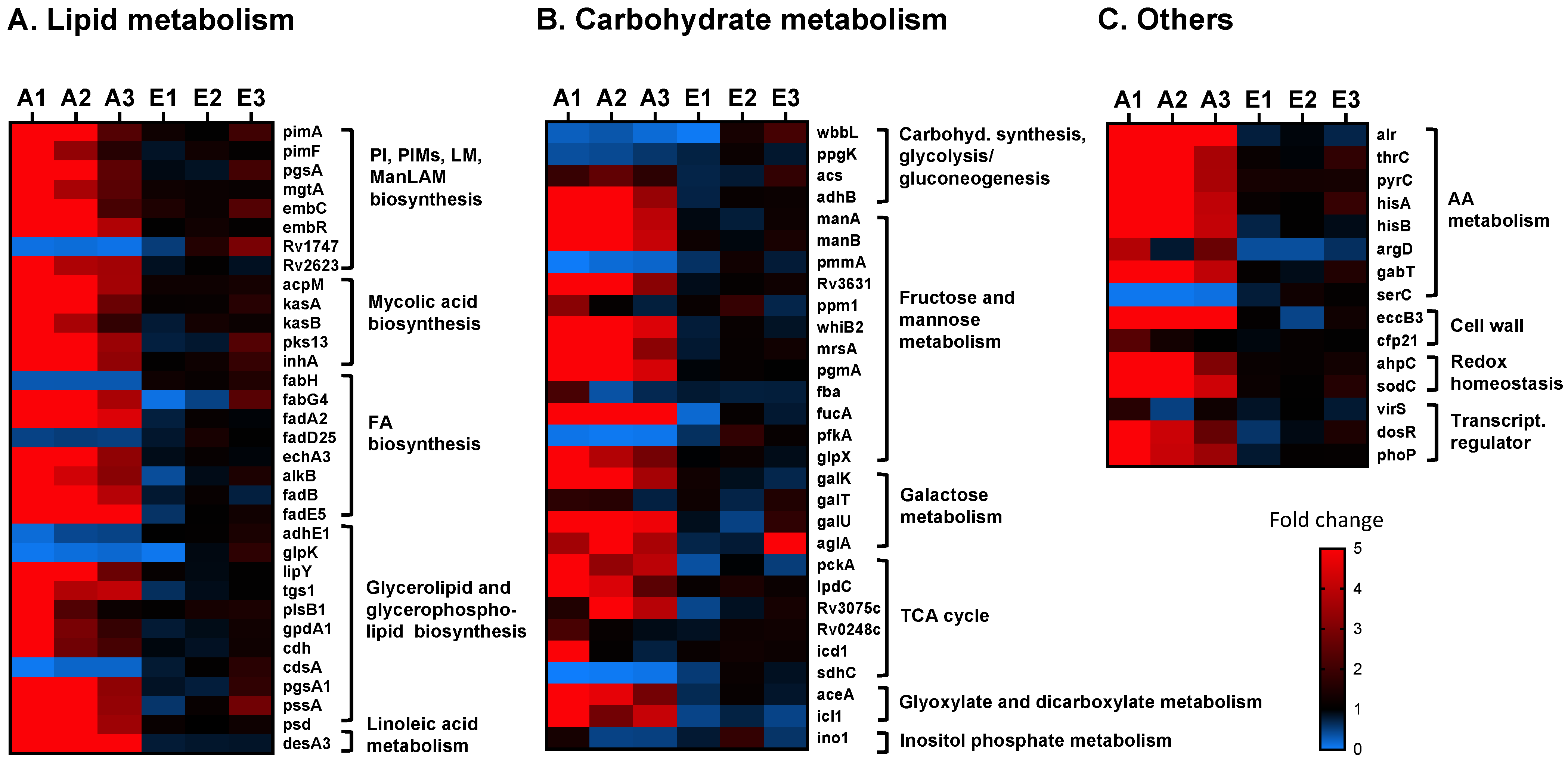
2.2. Effects of A- vs. E-ALF Exposure on the Expression of Cell Envelope Biosynthesis Genes in M.tb
3. Discussion
4. Materials and Methods
4.1. Human Subjects and Ethics Statement
4.2. Collection of BALF and ALF
4.3. Bacterial Cultures and ALF Exposure
4.4. RNA Extraction and cDNA Synthesis
4.5. qPCR Analysis of Targeted Genes
4.6. High-Throughput Multiplex qPCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Hogan, A.B.; Jewell, B.L.; Sherrard-Smith, E.; Vesga, J.F.; Watson, O.J.; Whittaker, C.; Hamlet, A.; Smith, J.A.; Winskill, P.; Verity, R.; et al. Potential impact of the COVID-19 pandemic on HIV, tuberculosis, and malaria in low-income and middle-income countries: A modelling study. Lancet Glob. Health 2020, 8, e1132–e1141. [Google Scholar] [CrossRef]
- Cilloni, L.; Fu, H.; Vesga, J.F.; Dowdy, D.; Pretorius, C.; Ahmedov, S.; Nair, S.A.; Mosneaga, A.; Masini, E.; Sahu, S.; et al. The potential impact of the COVID-19 pandemic on the tuberculosis epidemic a modelling analysis. EClinicalMedicine 2020, 28, 100603. [Google Scholar] [CrossRef]
- dos Santos, F.L.; Souza, L.L.L.; Bruce, A.T.I.; Crispim, J.D.; Arroyo, L.H.; Ramos, A.C.V.; Berra, T.Z.; Alves, Y.M.; Scholze, A.R.; da Costa, F.B.P.; et al. Patients’ perceptions regarding multidrug-resistant tuberculosis and barriers to seeking care in a priority city in Brazil during COVID-19 pandemic: A qualitative study. PLoS ONE 2021, 16, e0249822. [Google Scholar] [CrossRef]
- Marwah, V.; Peter, D.K.; Kumar, T.A.; Bhati, G.; Kumar, A. Multidrug-resistant tuberculosis in COVID-19: Double trouble. Med. J. Armed Forces India 2021, 77, S479–S482. [Google Scholar] [CrossRef]
- Tiberi, S.; Vjecha, M.J.; Zumla, A.; Galvin, J.; Migliori, G.B.; Zumla, A. Accelerating development of new shorter TB treatment regimens in anticipation of a resurgence of multi-drug resistant TB due to the COVID-19 pandemic. Int. J. Infect. Dis. 2021, 113, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Kalscheuer, R.; Palacios, A.; Anso, I.; Cifuente, J.; Anguita, J.; Jacobs, W.R., Jr.; Guerin, M.E.; Prados-Rosales, R. The Mycobacterium tuberculosis capsule: A cell structure with key implications in pathogenesis. Biochem. J. 2019, 476, 1995–2016. [Google Scholar] [CrossRef]
- Garcia-Vilanova, A.; Chan, J.; Torrelles, J.B. Underestimated manipulative roles of Mycobacterium tuberculosis cell envelope glycolipids during infection. Front. Immunol. 2019, 10, 2909. [Google Scholar] [CrossRef] [PubMed]
- Gago, G.; Diacovich, L.; Gramajo, H. Lipid metabolism and its implication in mycobacteria-host interaction. Curr. Opin. Microbiol. 2018, 41, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Dulberger, C.L.; Rubin, E.J.; Boutte, C.C. The mycobacterial cell envelope—A moving target. Nat. Rev. Microbiol 2020, 18, 47–59. [Google Scholar] [CrossRef]
- Vincent, A.T.; Nyongesa, S.; Morneau, I.; Reed, M.B.; Tocheva, E.I.; Veyrier, F.J. The mycobacterial cell envelope: A relict from the past or the result of recent evolution? Front. Microbiol. 2018, 9, 2341. [Google Scholar] [CrossRef] [Green Version]
- Chiaradia, L.; Lefebvre, C.; Parra, J.; Marcoux, J.; Burlet-Schiltz, O.; Etienne, G.; Tropis, M.; Daffe, M. Dissecting the mycobacterial cell envelope and defining the composition of the native mycomembrane. Sci. Rep. 2017, 7, 12807. [Google Scholar] [PubMed] [Green Version]
- Nicod, L.P. Pulmonary defence mechanisms. Respiration 1999, 66, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, P.S.; Bals, R. Series introduction: Innate host defense of the respiratory epithelium. J. Leukoc. Biol. 2004, 75, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.C. Alveolar type I cells: Molecular phenotype and development. Annu. Rev. Physiol. 2003, 65, 669–695. [Google Scholar] [CrossRef]
- Mason, R.J. Biology of alveolar type II cells. Respirology 2006, 11, S12–S15. [Google Scholar]
- Torrelles, J.B.; Schlesinger, L.S. Integrating lung physiology, immunology, and tuberculosis. Trends Microbiol. 2017, 25, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Liu, H.; Ge, B. Innate immunity in tuberculosis: Host defense vs. pathogen evasion. Cell Mol. Immunol. 2017, 14, 963–975. [Google Scholar] [CrossRef]
- Arcos, J.; Sasindran, S.J.; Fujiwara, N.; Turner, J.; Schlesinger, L.S.; Torrelles, J.B. Human lung hydrolases delineate Mycobacterium tuberculosis-macrophage interactions and the capacity to control infection. J. Immunol. 2011, 187, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Arcos, J.; Sasindran, S.J.; Moliva, J.I.; Scordo, J.M.; Sidiki, S.; Guo, H.; Venigalla, P.; Kelley, H.V.; Lin, G.; Diangelo, L.; et al. Mycobacterium tuberculosis cell wall released fragments by the action of the human lung mucosa modulate macrophages to control infection in an IL-10-dependent manner. Mucosal Immunol. 2017, 10, 1248–1258. [Google Scholar]
- Arcos, J.; Diangelo, L.E.; Scordo, J.M.; Sasindran, S.J.; Moliva, J.I.; Turner, J.; Torrelles, J.B. Lung mucosa lining fluid modification of Mycobacterium tuberculosis to reprogram human neutrophil killing mechanisms. J. Infect. Dis. 2015, 212, 948–958. [Google Scholar]
- Moliva, J.I.; Duncan, M.A.; Olmo-Fontanez, A.; Akhter, A.; Arnett, E.; Scordo, J.M.; Ault, R.; Sasindran, S.J.; Azad, A.K.; Montoya, M.J.; et al. The lung mucosa environment in the elderly increases host susceptibility to Mycobacterium tuberculosis infection. J. Infect. Dis. 2019, 220, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Scordo, J.M.; Arcos, J.; Kelley, H.V.; Diangelo, L.; Sasindran, S.J.; Youngmin, E.; Wewers, M.D.; Wang, S.H.; Balada-Llasat, J.M.; Torrelles, J.B. Mycobacterium tuberculosis cell wall fragments released upon bacterial contact with the human lung mucosa alter the neutrophil response to infection. Front. Immunol. 2017, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Moliva, J.I.; Rajaram, M.V.; Sidiki, S.; Sasindran, S.J.; Guirado, E.; Pan, X.J.; Wang, S.H.; Ross, P., Jr.; Lafuse, W.P.; Schlesinger, L.S.; et al. Molecular composition of the alveolar lining fluid in the aging lung. Age 2014, 36, 9633. [Google Scholar] [CrossRef] [Green Version]
- Olmo-Fontánez, A.M.; Scordo, J.M.; Garcia-Vilanova, A.; Maselli, D.J.; Peters, J.I.; Restrepo, B.I.; Clemens, D.L.; Turner, J.; Schlesinger, L.S.; Torrelles, J.B. Human alveolar lining fluid from the elderly promotes Mycobacterium tuberculosis growth in alveolar epithelial cells and bacterial translocation into the cytosol. bioRxiv, 2021; in press. [Google Scholar]
- Keiser, T.L.; Azad, A.K.; Guirado, E.; Bonacci, R.; Schlesinger, L.S. Comparative transcriptional study of the putative mannose donor biosynthesis genes in virulent Mycobacterium tuberculosis and attenuated Mycobacterium bovis BCG strains. Infect. Immun. 2011, 79, 4668–4673. [Google Scholar] [CrossRef] [Green Version]
- Guirado, E.; Mbawuike, U.; Keiser, T.L.; Arcos, J.; Azad, A.K.; Wang, S.H.; Schlesinger, L.S. Characterization of host and microbial determinants in individuals with latent tuberculosis infection using a human granuloma model. mBio 2015, 6, e02537-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papavinasasundaram, K.G.; Chan, B.; Chung, J.H.; Colston, M.J.; Davis, E.O.; Av-Gay, Y. Deletion of the Mycobacterium tuberculosis pknH gene confers a higher bacillary load during the chronic phase of infection in BALB/c mice. J. Bacteriol. 2005, 187, 5751–5760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treumann, A.; Xidong, F.; McDonnell, L.; Derrick, P.J.; Ashcroft, A.E.; Chatterjee, D.; Homans, S.W. 5-Methylthiopentose: A new substituent on lipoarabinomannan in Mycobacterium tuberculosis. J. Mol. Biol. 2002, 316, 89–100. [Google Scholar] [CrossRef]
- Turnbull, W.B.; Shimizu, K.H.; Chatterjee, D.; Homans, S.W.; Treumann, A. Identification of the 5-methylthiopentosyl substituent in Mycobacterium tuberculosis lipoarabinomannan. Angew. Chem. Int. Ed. 2004, 43, 3918–3922. [Google Scholar] [CrossRef]
- Angala, S.K.; McNeil, M.R.; Shi, L.; Joe, M.; Pham, H.; Zuberogoitia, S.; Nigou, J.; Boot, C.M.; Lowary, T.L.; Gilleron, M.; et al. Biosynthesis of the methylthioxylose capping motif of lipoarabinomannan in Mycobacterium tuberculosis. ACS Chem. Biol. 2017, 12, 682–691. [Google Scholar] [CrossRef]
- McCarthy, T.R.; Torrelles, J.B.; MacFarlane, A.S.; Katawczik, M.; Kutzbach, B.; Desjardin, L.E.; Clegg, S.; Goldberg, J.B.; Schlesinger, L.S. Overexpression of Mycobacterium tuberculosis manB, a phosphomannomutase that increases phosphatidylinositol mannoside biosynthesis in Mycobacterium smegmatis and mycobacterial association with human macrophages. Mol. Microbiol. 2005, 58, 774–790. [Google Scholar] [CrossRef]
- Rana, A.K.; Singh, A.; Gurcha, S.S.; Cox, L.R.; Bhatt, A.; Besra, G.S. Ppm1-encoded polyprenyl monophosphomannose synthase activity is essential for lipoglycan synthesis and survival in mycobacteria. PLoS ONE 2012, 7, e48211. [Google Scholar] [CrossRef]
- Gurcha, S.S.; Baulard, A.R.; Kremer, L.; Locht, C.; Moody, D.B.; Muhlecker, W.; Costello, C.E.; Crick, D.C.; Brennan, P.J.; Besra, G.S. Ppm1, a novel polyprenol monophosphomannose synthase from Mycobacterium tuberculosis. Biochem. J. 2002, 365 Pt 2, 441–450. [Google Scholar] [CrossRef]
- Lv, X.; Ma, C.Y.; Yan, Z.F.; Deng, R.; Ai, X.F.; Su, T.; Xiang, X.H.; Li, W. The Mycobacterium tuberculosis protein Rv2387 is involved in cell wall remodeling and susceptibility to acidic conditions. Biochem. Bioph. Res. 2018, 503, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Stokas, H.; Rhodes, H.L.; Purdy, G.E. Modulation of the M. tuberculosis cell envelope between replicating and non-replicating persistent bacteria. Tuberculosis 2020, 125, 102007. [Google Scholar] [CrossRef]
- Howard, N.C.; Marin, N.D.; Ahmed, M.; Rosa, B.A.; Martin, J.; Bambouskova, M.; Sergushichev, A.; Loginicheva, E.; Kurepina, N.; Rangel-Moreno, J.; et al. Mycobacterium tuberculosis carrying a rifampicin drug resistance mutation reprograms macrophage metabolism through cell wall lipid changes. Nat. Microbiol. 2018, 3, 1099–1108. [Google Scholar] [CrossRef]
- Bacon, J.; Alderwick, L.J.; Allnutt, J.A.; Gabasova, E.; Watson, R.; Hatch, K.A.; Clark, S.O.; Jeeves, R.E.; Marriott, A.; Rayner, E.; et al. Non-replicating Mycobacterium tuberculosis elicits a reduced infectivity profile with corresponding modifications to the cell wall and extracellular matrix. PLoS ONE 2014, 9, e87329. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.P.S.; Guan, X.L. Metabolic versatility of Mycobacterium tuberculosis during infection and dormancy. Metabolites 2021, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sinha, T.; Carlson, T.K.; Keiser, T.L.; Torrelles, J.B.; Schlesinger, L.S. Changes in the major cell envelope components of Mycobacterium tuberculosis during in vitro growth. Glycobiology 2013, 23, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Spurgeon, S.L.; Jones, R.C.; Ramakrishnan, R. High throughput gene expression measurement with real time PCR in a microfluidic dynamic array. PLoS ONE 2008, 3, e1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, K.Y.; de Carvalho, L.P.; Bryk, R.; Ehrt, S.; Marrero, J.; Park, S.W.; Schnappinger, D.; Venugopal, A.; Nathan, C. Central carbon metabolism in Mycobacterium tuberculosis: An unexpected frontier. Trends Microbiol. 2011, 19, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Glass, L.N.; Swapna, G.; Chavadi, S.S.; Tufariello, J.M.; Mi, K.; Drumm, J.E.; Lam, T.T.; Zhu, G.; Zhan, C.; Vilcheze, C.; et al. Mycobacterium tuberculosis universal stress protein Rv2623 interacts with the putative ATP binding cassette (ABC) transporter Rv1747 to regulate mycobacterial growth. PLoS Pathog. 2017, 13, e1006515. [Google Scholar] [CrossRef]
- Phong, W.Y.; Lin, W.; Rao, S.P.; Dick, T.; Alonso, S.; Pethe, K. Characterization of phosphofructokinase activity in Mycobacterium tuberculosis reveals that a functional glycolytic carbon flow is necessary to limit the accumulation of toxic metabolic intermediates under hypoxia. PLoS ONE 2013, 8, e56037. [Google Scholar] [CrossRef]
- Konyarikova, Z.; Savkova, K.; Kozmon, S.; Mikusova, K. Biosynthesis of galactan in Mycobacterium tuberculosis as a viable TB drug target? Antibiotics 2020, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, T.; Weinrick, B.; Vilcheze, C.; Berney, M.; Tufariello, J.; Cook, G.M.; Jacobs, W.R., Jr. Succinate dehydrogenase is the regulator of respiration in Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, e1004510. [Google Scholar] [CrossRef] [PubMed]
- Movahedzadeh, F.; Smith, D.A.; Norman, R.A.; Dinadayala, P.; Murray-Rust, J.; Russell, D.G.; Kendall, S.L.; Rison, S.C.; McAlister, M.S.; Bancroft, G.J.; et al. The Mycobacterium tuberculosis ino1 gene is essential for growth and virulence. Mol. Microbiol. 2004, 51, 1003–1014. [Google Scholar] [CrossRef]
- Haufroid, M.; Wouters, J. Targeting the serine pathway: A promising approach against tuberculosis? Pharmaceuticals 2019, 12, 66. [Google Scholar] [CrossRef] [Green Version]
- Famelis, N.; Rivera-Calzada, A.; Degliesposti, G.; Wingender, M.; Mietrach, N.; Skehel, J.M.; Fernandez-Leiro, R.; Bottcher, B.; Schlosser, A.; Llorca, O.; et al. Architecture of the mycobacterial type VII secretion system. Nature 2019, 576, 321–325. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Z.; Fu, R.; Zhang, Y.; Chen, L.; Huang, L.; Li, J.; Shi, C.; Fan, X. A DNA vaccine expressing CFP21 and MPT64 fusion protein enhances BCG-induced protective immunity against Mycobacterium tuberculosis infection in mice. Med. Microbiol. Immunol. 2011, 200, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Renier, W.; Bourdin, A.; Rubbo, P.A.; Peries, M.; Dedieu, L.; Bendriss, S.; Kremer, L.; Canaan, S.; Terru, D.; Godreuil, S.; et al. B cells response directed against Cut4 and CFP21 lipolytic enzymes in active and latent tuberculosis infections. PLoS ONE 2018, 13, e0196470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.F.; Shin, J.; Manimekalai, M.S.S.; Saw, W.G.; Yin, Z.; Bhushan, S.; Kumar, A.; Ragunathan, P.; Gruber, G. AhpC of the mycobacterial antioxidant defense system and its interaction with its reducing partner Thioredoxin-C. Sci. Rep. 2017, 7, 5159. [Google Scholar] [CrossRef]
- Liao, D.; Fan, Q.; Bao, L. The role of superoxide dismutase in the survival of Mycobacterium tuberculosis in macrophages. Jpn. J. Infect. Dis. 2013, 66, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Singh, A.; Tyagi, A.K. Deciphering the genes involved in pathogenesis of Mycobacterium tuberculosis. Tuberculosis 2005, 85, 325–335. [Google Scholar] [CrossRef]
- Turner, J.; Torrelles, J.B. Mannose-capped lipoarabinomannan in Mycobacterium tuberculosis pathogenesis. Pathog. Dis. 2018, 76, fty026. [Google Scholar] [CrossRef] [Green Version]
- Torrelles, J.B.; Schlesinger, L.S. Diversity in Mycobacterium tuberculosis mannosylated cell wall determinants impacts adaptation to the host. Tuberculosis 2010, 90, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Fratti, R.A.; Backer, J.M.; Gruenberg, J.; Corvera, S.; Deretic, V. Role of phosphatidylinositol 3-kinase and Rab5 effectors in phagosomal biogenesis and mycobacterial phagosome maturation arrest. J. Cell Biol. 2001, 154, 631–644. [Google Scholar] [CrossRef] [Green Version]
- Rao, N.; Meena, L.S. Biosynthesis and virulent behavior of lipids produced by Mycobacterium tuberculosis: LAM and cord factor: An overview. Biotechnol. Res. Int. 2011, 2011, 274693. [Google Scholar]
- Kang, P.B.; Azad, A.K.; Torrelles, J.B.; Kaufman, T.M.; Beharka, A.; Tibesar, E.; DesJardin, L.E.; Schlesinger, L.S. The human macrophage mannose receptor directs Mycobacterium tuberculosis lipoarabinomannan-mediated phagosome biogenesis. J. Exp. Med. 2005, 202, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Wietzerbin-Falszpan, J.; Das, B.C.; Gros, C.; Petit, J.F.; Lederer, E. The amino acids of the cell wall of Mycobacterium tuberculosis var. bovis, strain BCG. Presence of a poly (L-glutamic acid). Eur. J. Biochem. 1973, 32, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Alderwick, L.J.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The mycobacterial cell wall—Peptidoglycan and arabinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, a021113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cava, F.; de Pedro, M.A.; Lam, H.; Davis, B.M.; Waldor, M.K. Distinct pathways for modification of the bacterial cell wall by non-canonical D-amino acids. EMBO J. 2011, 30, 3442–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitra, A.; Munshi, T.; Healy, J.; Martin, L.T.; Vollmer, W.; Keep, N.H.; Bhakta, S. Cell wall peptidoglycan in Mycobacterium tuberculosis: An Achilles’ heel for the TB-causing pathogen. FEMS Microbiol. Rev. 2019, 43, 548–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Heredia, A.; Pohane, A.A.; Melzer, E.S.; Carr, C.R.; Fiolek, T.J.; Rundell, S.R.; Lim, H.C.; Wagner, J.C.; Morita, Y.S.; Swarts, B.M.; et al. Peptidoglycan precursor synthesis along the sidewall of pole-growing mycobacteria. Elife 2018, 7, e37243. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; De Buck, J. Cell wall proteome analysis of Mycobacterium smegmatis strain MC2 155. BMC Microbiol. 2010, 10, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehra, S.; Foreman, T.; Didier, P.J.; Ahsan, M.H.; Hudock, T.A.; Kissee, R.; Golden, N.A.; Gautam, U.; Johnson, A.-M.; Alvarez, X.; et al. The DosR Regulon Modulates Adaptive Immunity and Is Essential forMycobacterium tuberculosisPersistence. Am. J. Respir. Crit. Care Med. 2015, 191, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Ryndak, M.; Wang, S.; Smith, I. PhoP, a key player in Mycobacterium tuberculosis virulence. Trends Microbiol. 2008, 16, 528–534. [Google Scholar] [CrossRef]
- Broset, E.; Martin, C.; Gonzalo-Asensio, J. Evolutionary landscape of the Mycobacterium tuberculosis complex from the viewpoint of PhoPR: Implications for virulence regulation and application to vaccine development. mBio 2015, 6, e01289-15. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Crossman, D.K.; Mai, D.; Guidry, L.; Voskuil, M.I.; Renfrow, M.B.; Steyn, A.J. Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response. PLoS Pathog. 2009, 5, e1000545. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.B.; Gagneux, S.; DeRiemer, K.; Small, P.M.; Barry, C.E. The W-Beijing lineage of Mycobacterium tuberculosis overproduces triglycerides and has the DosR dormancy regulon constitutively upregulated. J. Bacteriol. 2007, 189, 2583–2589. [Google Scholar] [CrossRef] [Green Version]
- Scordo, J.M.; Olmo-Fontanez, A.M.; Kelley, H.V.; Sidiki, S.; Arcos, J.; Akhter, A.; Wewers, M.D.; Torrelles, J.B. The human lung mucosa drives differential Mycobacterium tuberculosis infection outcome in the alveolar epithelium. Mucosal Immunol. 2019, 12, 795–804. [Google Scholar] [CrossRef]
- Shaku, M.; Ealand, C.; Kana, B.D. Cell surface biosynthesis and remodeling pathways in mycobacteria reveal new drug targets. Front. Cell Infect. Microbiol. 2020, 10, 603382. [Google Scholar] [CrossRef]
- Drumm, J.E.; Mi, K.; Bilder, P.; Sun, M.; Lim, J.; Bielefeldt-Ohmann, H.; Basaraba, R.; So, M.; Zhu, G.; Tufariello, J.M.; et al. Mycobacterium tuberculosis universal stress protein Rv2623 regulates bacillary growth by ATP-Binding: Requirement for establishing chronic persistent infection. PLoS Pathog. 2009, 5, e1000460. [Google Scholar] [CrossRef]
- Curry, J.M.; Whalan, R.; Hunt, D.M.; Gohil, K.; Strom, M.; Rickman, L.; Colston, M.J.; Smerdon, S.J.; Buxton, R.S. An ABC transporter containing a forkhead-associated domain interacts with a serine-threonine protein kinase and is required for growth of Mycobacterium tuberculosis in mice. Infect. Immun. 2005, 73, 4471–4477. [Google Scholar] [CrossRef] [Green Version]
- Allué-Guardia, A.; Garcia, J.I.; Torrelles, J.B. Evolution of drug-resistant Mycobacterium tuberculosis strains and their adaptation to the human lung environment. Front. Microbiol. 2021, 12, 612675. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- You, F.M.; Huo, N.; Gu, Y.Q.; Luo, M.C.; Ma, Y.; Hane, D.; Lazo, G.R.; Dvorak, J.; Anderson, O.D. BatchPrimer3: A high throughput web application for PCR and sequencing primer design. BMC Bioinform. 2008, 9, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E., 3rd; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef]
- Camus, J.C.; Pryor, M.J.; Medigue, C.; Cole, S.T. Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv. Microbiology 2002, 148 Pt 10, 2967–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lew, J.M.; Kapopoulou, A.; Jones, L.M.; Cole, S.T. TubercuList—10 years after. Tuberculosis 2011, 91, 1–7. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locus Tag H37Rv | Gene Name | Product and Function | Primer Sequence (5′ to 3′) | Tm (°C) | Ref. |
|---|---|---|---|---|---|
| Rv0667 | rpoB | Reference gene | rpoB-F: CCTGGAAGAGGTGCTCTACG rpoB-R: GGGAAGTCACCCATGAACAC | 60 60 | [26] [26] |
| Rv2703 | sigA | Reference gene | sigA-F: CTCGGTTCGCGCCTACCTCA sigA-R: GCGCTCGCTAAGCTCGGTCA | 68 68 | [28] [28] |
| Rv3255c | manA | mannose-6-phosphate isomerase/GDP-Man biosynthesis: formation of Man-6-P | manA-F: GTTCACCACCTGGATTACCG manA-R: AACCCTCGGTGCATAACAAG | 60 60 | [26] [27] |
| Rv3264c | manBa | D-alpha-D-mannose-1-phosphate guanylyltransferase/GDP-Man biosynthesis: formation of GDP-Man | manB-F: ACATCGCCGTTAAACACCAT manB-R: GTTCCTCACCCATCTGCTGT | 60 60 | [27] [27] |
| Rv3257c | pmmAb | phosphomannose mutase/GDP-Man biosynthesis: formation of Man-1-P | pmmA-F: GATCACGTTGTGGATGATGG pmmA-R: GTGGATCTGCAGGCCTATGT | 60 60 | [26] [27] |
| Rv3308 | pmmB | phosphomannose mutase/GDP-Man biosynthesis: formation of Man-1-P | pmmB-F: ATACAGATCACGGCGTCACA pmmB-R: CGCTGGATATAACGGTCGAT | 60 60 | [27] [27] |
| Rv2051c | ppm1 | Polyprenol-monophosphomannose synthase/PPM biosynthesis | pmm1-F: TGGTTGAAGTCGATCCTTCC pmm1-R: GCGAACAAGACCAGGCATATG | 60 63 | [26] [26] |
| Rv3260c | whiB2 | Transcript. regulatory protein | whiB2-F: CCATTCGAGGAACCTCTGC whiB2-R: CAGGGCGTACTCCAGACACT | 61 60 | [26] [26] |
| Rv2610c | pimA | alpha-(1-2)-phosphatidylinositol mannosyl-transferase/PIM biosynthesis (1st step) | pimA-F: CCGCACTGCCTGATTACTTT pimA-R: CGGCTCGTGTAGATGCAGTA | 60 60 | [27] [27] |
| Rv2188c | pimB | alpha-(1-6)-phosphatidylinositol mannosyl- transferase/PIM biosynthesis (2nd step) | pimB-F: CTCGGTGGTCAAGGTACTCG pimB-R: GTGGTCACCTTTGGGAACAT | 61 60 | [27] [27] |
| Rv1500 | pimF | glycosyltransferase/LM/ ManLAM biosynthesis | pimF-F: CGCCGACGTAGTATTTGGTT pimF-R: TGCGTACATAGTCGGCTGTC | 60 60 | [27] [27] |
| Rv3793 | embC | Arabynosyl-tranferase/ManLAM biosynthesis | embC-F: ATCACCGAGCTGCTGATG embC-R: TGCGAGTCACCGTTCCTA | 58 59 | [28] [28] |
| Genes | 15 min vs. 12 h (rpoB) | 15 min vs. 12 h (sigA) | Statistical Significance |
|---|---|---|---|
| ppm1 | −0.4330 | −0.5129 | ns |
| manA | 0.2125 | 0.142 | ns |
| manB | 0.01161 | 0.0622 | ns |
| pmmA | −0.9634 | −0.9820 | *** |
| whiB2 | 0.03380 | 0.0271 | ns |
| pimA | −0.06219 | −0.0576 | ns |
| pimB | −0.02300 | 0.0218 | ns |
| pimF | −0.1283 | −0.129 | ns |
| embC | 0.1572 | 0.2012 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allué-Guardia, A.; Garcia-Vilanova, A.; Olmo-Fontánez, A.M.; Peters, J.; Maselli, D.J.; Wang, Y.; Turner, J.; Schlesinger, L.S.; Torrelles, J.B. Host- and Age-Dependent Transcriptional Changes in Mycobacterium tuberculosis Cell Envelope Biosynthesis Genes after Exposure to Human Alveolar Lining Fluid. Int. J. Mol. Sci. 2022, 23, 983. https://doi.org/10.3390/ijms23020983
Allué-Guardia A, Garcia-Vilanova A, Olmo-Fontánez AM, Peters J, Maselli DJ, Wang Y, Turner J, Schlesinger LS, Torrelles JB. Host- and Age-Dependent Transcriptional Changes in Mycobacterium tuberculosis Cell Envelope Biosynthesis Genes after Exposure to Human Alveolar Lining Fluid. International Journal of Molecular Sciences. 2022; 23(2):983. https://doi.org/10.3390/ijms23020983
Chicago/Turabian StyleAllué-Guardia, Anna, Andreu Garcia-Vilanova, Angélica M. Olmo-Fontánez, Jay Peters, Diego J. Maselli, Yufeng Wang, Joanne Turner, Larry S. Schlesinger, and Jordi B. Torrelles. 2022. "Host- and Age-Dependent Transcriptional Changes in Mycobacterium tuberculosis Cell Envelope Biosynthesis Genes after Exposure to Human Alveolar Lining Fluid" International Journal of Molecular Sciences 23, no. 2: 983. https://doi.org/10.3390/ijms23020983