
Title IgA Nephropathy and Oral Bacterial Species Related to Dental Caries and Periodontitis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Historical Findings
3. Nasopharynx-Associated Lymphoid Tissue (NALT) and Gut-Associated Lymphoid Tissue (GALT)
4. NALT and IgA Nephropathy
5. GALT and IgA Nephropathy
6. IgA Nephropathy and Cnm-(+) S. mutans: Clinical Evidence
7. IgA Nephropathy and Cnm-(+) S. mutans: Animal Models
8. IgA and Periodontal Bacteria
9. IgA and Periodontal Bacteria: Animal Models
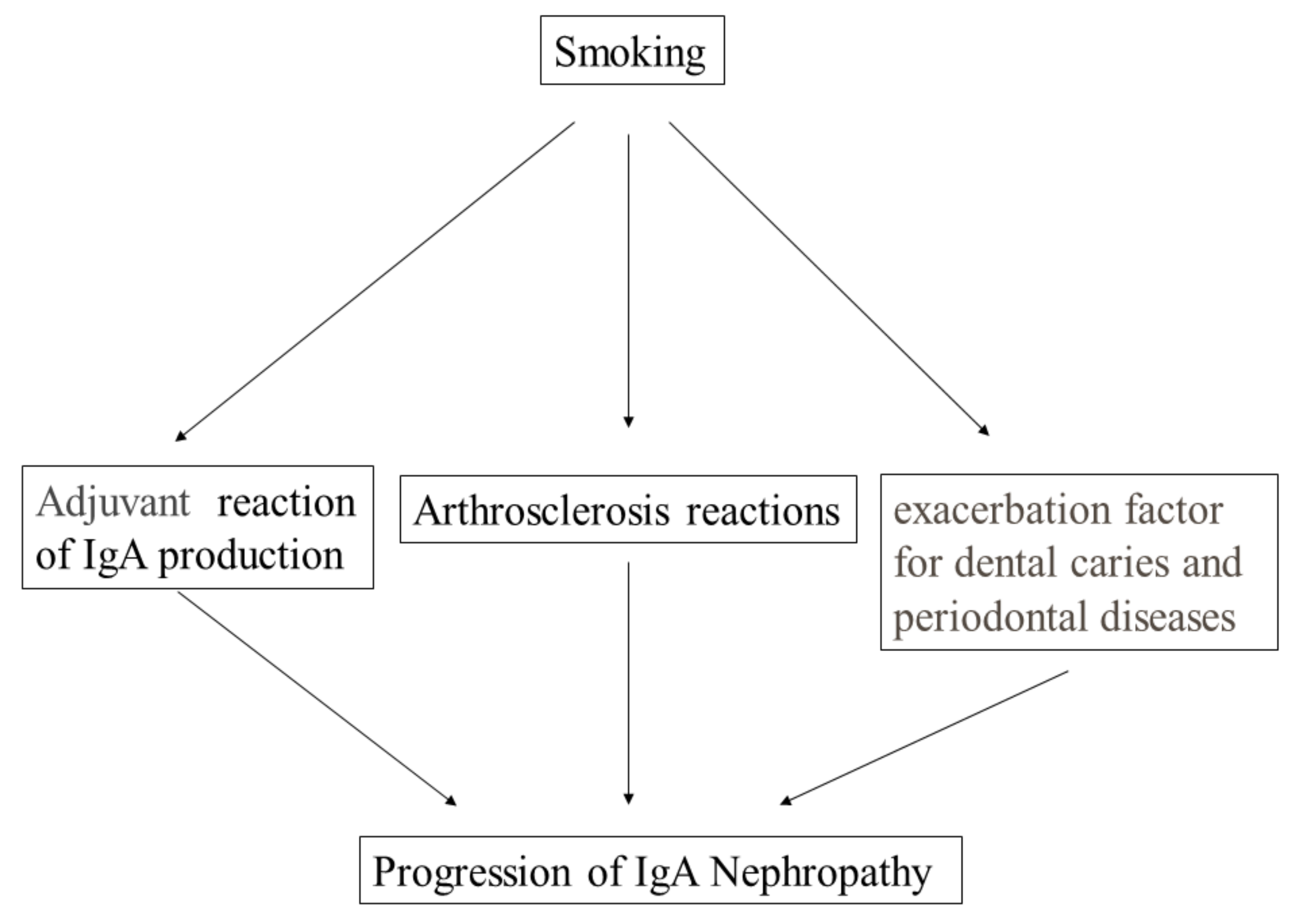
10. IgA and Cigarette Smoking
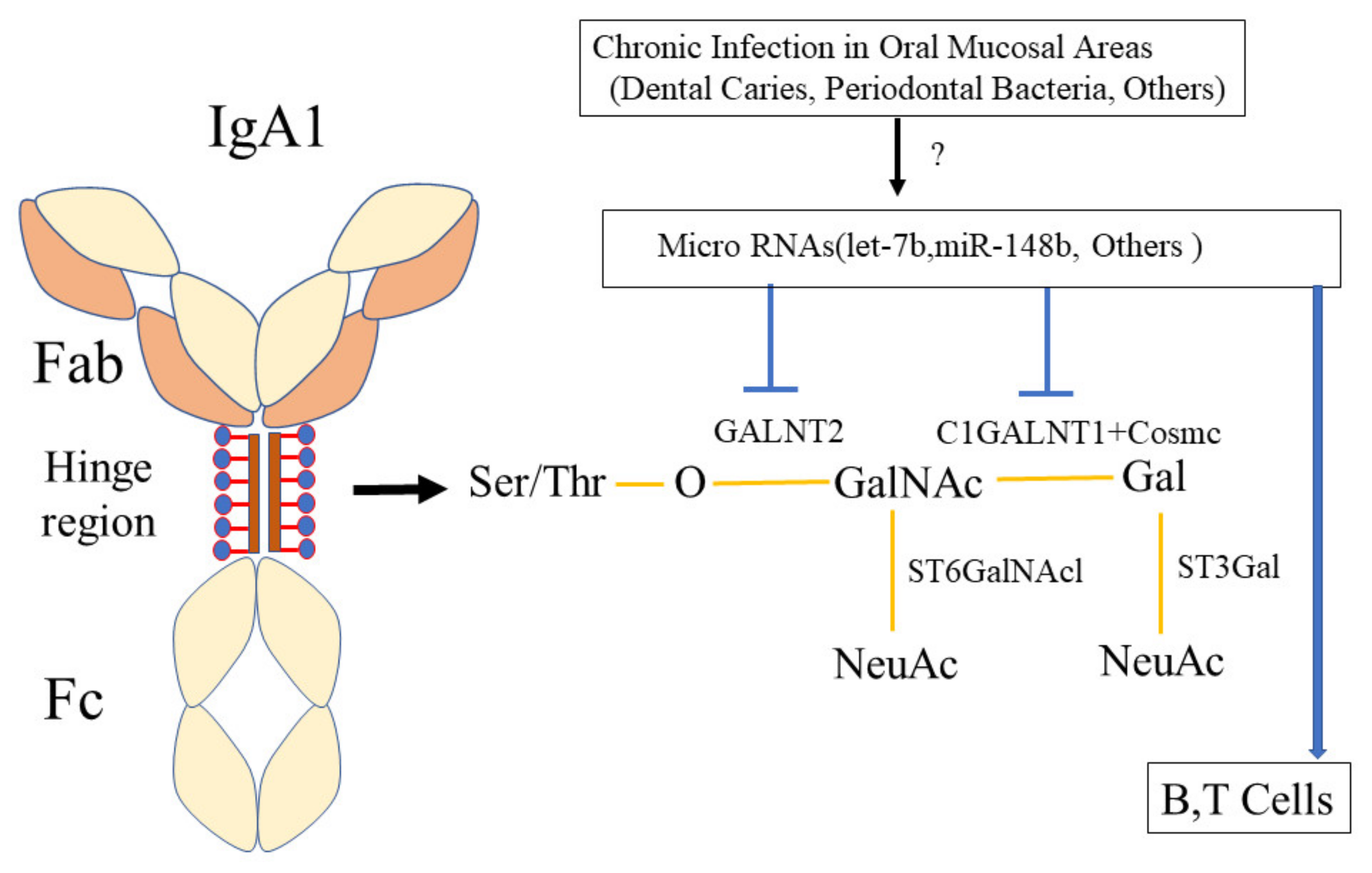
11. Hypothesis of the Mechanism by Which Oral Infection Induces Aberrantly Glycosylated IgA1, Which May Induce IgA Nephropathy
12. Gene Influence on The Relationship between IgA and Infection
13. Conclusions Regarding Oral Bacteria and IgA Nephropathy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wyatt, R.J.; Julian, B.A. IgA Nephropathy. N. Engl. J. Med. 2013, 368, 2402–2414. [Google Scholar] [CrossRef] [Green Version]
- Lai, K.N.; Tang, S.C.; Schena, F.P.; Novak, J.; Tomino, Y.; Fogo, A.B.; Glassock, R.J. IgA nephropathy. Nat. Rev. Dis. Prim. 2016, 2, 16001. [Google Scholar] [CrossRef]
- Selvaskandan, H.; Cheung, C.K.; Muto, M.; Barratt, J. New strategies and perspectives on managing IgA nephropathy. Clin. Exp. Nephrol. 2019, 23, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Maixnerova, D.; Tesar, V. Emerging modes of treatment of IgA nephropathy. Int. J. Mol. Sci. 2020, 21, 9064. [Google Scholar] [CrossRef]
- Sallustio, F.; Curci, C.; Di Leo, V.; Gallone, A.; Pesce, F.; Gesualdo, L. A New vision of IgA nephropathy: The missing link. Int. J. Mol. Sci. 2019, 21, 189. [Google Scholar] [CrossRef] [Green Version]
- Nagasawa, Y.; Yamamoto, R.; Shoji, T.; Shinzawa, M.; Hasuike, Y.; Nagatoya, K.; Yamauchi, A.; Hayashi, T.; Kuragano, T.; Moriyama, T.; et al. Serum uric acid level predicts progression of IgA nephropathy in females but not in males. PLoS ONE 2016, 11, e0160828. [Google Scholar] [CrossRef]
- Yamamoto, R.; Nagasawa, Y.; Shoji, T.; Iwatani, H.; Hamano, T.; Kawada, N.; Inoue, K.; Uehata, T.; Kaneko, T.; Okada, N.; et al. Cigarette smoking and progression of IgA nephropathy. Am. J. Kidney Dis. 2010, 56, 313–324. [Google Scholar] [CrossRef]
- Hotta, O.; Miyazaki, M.; Furuta, T.; Tomioka, S.; Chiba, S.; Horigome, I.; Abe, K.; Taguma, Y. Tonsillectomy and steroid pulse therapy significantly impact on clinical remission in patients with IgA nephropathy. Am. J. Kidney Dis. 2001, 38, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Yoshimura, M.; Miyazaki, Y.; Okamoto, H.; Kimura, K.; Hirano, K.; Matsushima, M.; Utsunomiya, Y.; Ogura, M.; Yokoo, T.; et al. A multicenter randomized controlled trial of tonsillectomy combined with steroid pulse therapy in patients with immunoglobulin A nephropathy. Nephrol. Dial. Transplant. 2014, 29, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Matsuzaki, K.; Yasuda, T.; Nishikawa, M.; Yasuda, Y.; Koike, K.; Maruyama, S.; Yokoo, T.; Matsuo, S.; Kawamura, T.; et al. Association between tonsillectomy and outcomes in patients with immunoglobulin a nephropathy. JAMA Netw. Open 2019, 2, e194772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, K.; Hokamura, K.; Taniguchi, N.; Wada, K.; Kudo, C.; Nomura, R.; Kojima, A.; Naka, S.; Muranaka, Y.; Thura, M.; et al. The collagen-binding protein of Streptococcus mutans is involved in haemorrhagic stroke. Nat. Commun. 2011, 2, 485. [Google Scholar] [CrossRef] [Green Version]
- Kojima, A.; Nakano, K.; Wada, K.; Takahashi, H.; Katayama, K.; Yoneda, M.; Higurashi, T.; Nomura, R.; Hokamura, K.; Muranaka, Y.; et al. Infection of specific strains of Streptococcus mutans, oral bacteria, confers a risk of ulcerative colitis. Sci. Rep. 2012, 2, 332. [Google Scholar] [CrossRef] [Green Version]
- Naka, S.; Nomura, R.; Takashima, Y.; Okawa, R.; Ooshima, T.; Nakano, K. A specific Streptococcus mutans strain aggravates non-alcoholic fatty liver disease. Oral Dis. 2014, 20, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Naka, S.; Hatakeyama, R.; Takashima, Y.; Matsumoto-Nakano, M.; Nomura, R.; Nakano, K. Contributions of Streptococcus mutans Cnm and PA antigens to aggravation of non-alcoholic steatohepatitis in mice. Sci. Rep. 2016, 6, 36886. [Google Scholar] [CrossRef] [PubMed]
- Tonomura, S.; Ihara, M.; Kawano, T.; Tanaka, T.; Okuno, Y.; Saito, S.; Friedland, R.P.; Kuriyama, N.; Nomura, R.; Watanabe, Y.; et al. Intracerebral hemorrhage and deep microbleeds associated with cnm-positive Streptococcus mutans; a hospital cohort study. Sci. Rep. 2016, 6, 20074. [Google Scholar] [CrossRef]
- Watanabe, I.; Kuriyama, N.; Miyatani, F.; Nomura, R.; Naka, S.; Nakano, K.; Ihara, M.; Iwai, K.; Matsui, D.; Ozaki, E.; et al. Oral cnm-positive streptococcus mutans expressing collagen binding activity is a risk factor for cerebral microbleeds and cognitive impairment. Sci. Rep. 2016, 6, 38561. [Google Scholar] [CrossRef] [Green Version]
- Satoskar, A.A.; Parikh, S.V.; Nadasdy, T. Epidemiology, pathogenesis, treatment and outcomes of infection-associated glomerulonephritis. Nat. Rev. Nephrol. 2020, 16, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, J.; Pedreañez, A. Acute post-streptococcal glomerulonephritis: Analysis of the pathogenesis. Int. Rev. Immunol. 2021, 40, 381–400. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakatomi, Y.; Sato, H.; Tsukada, H.; Arakawa, M. Haemophilus parainfluenzae antigen and antibody in renal biopsy samples and serum of patients with IgA nephropathy. Lancet 1994, 343, 12–16. [Google Scholar] [CrossRef]
- Suzuki, S.; Fujieda, S.; Sunaga, H.; Yamamoto, C.; Kimura, H.; Gejyo, F. Synthesis of immunoglobulins against Haemophilus parainfluenzae by tonsillar lymphocytes from patients with IgA nephropathy. Nephrol. Dial. Transplant. 2000, 15, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, C.; Suzuki, S.; Kimura, H.; Yoshida, H.; Gejyo, F. Experimental nephropathy induced by Haemophilus parainfluenzae antigens. Nephron 2002, 90, 320–327. [Google Scholar] [CrossRef]
- Day, N.K.; Geiger, H.; McLean, R.; Resnick, J.; Michael, A.; Good, R.A. The association of respiratory infection, recurrent hematuria, and focal glomerulonephritis with activation of the complement system in the cold. J. Clin. Investig. 1973, 52, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Forsström, J.; Viander, M.; Lehtonen, A.; Ekfors, T. Yersinia enterocolitica infection complicated by glomerulonephritis. Scand. J. Infect. Dis. 1977, 9, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Tomino, Y.; Endoh, M.; Nomoto, Y.; Sakai, H. Immunoglobulin A1 in IgA Nephropathy. N. Engl. J. Med. 1981, 305, 1159–1160. [Google Scholar] [CrossRef]
- Tomino, Y.; Sakai, H.; Miura, M.; Endoh, M.; Nomoto, Y. Detection of polymeric IgA in glomeruli from patients with IgA nephropathy. Clin. Exp. Immunol. 1982, 49, 419–425. [Google Scholar]
- Monteiro, R.C.; Halbwachs-Mecarelli, L.; Roque-Barreira, M.C.; Noel, L.-H.; Berger, J.; Lesavre, P. Charge and size of mesangial IgA in IgA nephropathy. Kidney Int. 1985, 28, 666–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, S.J.; Feehally, J. The pathogeneic role of immunoglobin APolymers in immunoglobulin a nephropathy. Nephron 1993, 65, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Tomino, Y. The mucosa-bone-marrow axis in IgA Nephropathy. Contrib. Nephrol. 2007, 157, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Pabst, R.; Russell, M.W.; Brandtzaeg, P. Tissue distribution of lymphocytes and plasma cells and the role of the gut. Trends Immunol. 2008, 29, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.R.; Efroni, S. The Immune System Computes the State of the Body: Crowd Wisdom, Machine Learning, and Immune Cell Reference Repertoires Help Manage Inflammation. Front. Immunol. 2019, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Brandtzaeg, P.; Johansen, F.-E. Mucosal B cells: Phenotypic characteristics, transcriptional regulation, and homing properties. Immunol. Rev. 2005, 206, 32–63. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.C.; Bailey, E.M.; Brenchley, P.E.; Buck, K.S.; Barratt, J.; Feehally, J. Mesangial IgA1 in IgA nephropathy exhibits aberrant O-glycosylation: Observations in three patients. Kidney Int. 2001, 60, 969–973. [Google Scholar] [CrossRef] [Green Version]
- Hiki, Y.; Odani, H.; Takahashi, M.; Yasuda, Y.; Nishimoto, A.; Iwase, H.; Shinzato, T.; Kobayashi, Y.; Maeda, K. Mass spectrometry proves under-O-glycosylation of glomerular IgA1 in IgA nephropathy. Kidney Int. 2001, 59, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floege, J.; Feehally, J. IgA Nephropathy: Recent Developments. J. Am. Soc. Nephrol. 2000, 11, 2395–2403. [Google Scholar] [CrossRef]
- Novak, K.F.; Nonnemacher, M.R.; Pourhamidi, J. Molecular Characterization and Functional Analysis of asecAGene Homolog inActinobacillus actinomycetemcomitans. Microbiol. Immunol. 2000, 44, 143–148. [Google Scholar] [CrossRef]
- Van Der Boog, P.J.; van Kooten, C.; De Fijter, J.W.; Daha, M.R. Role of macromolecular IgA in IgA nephropathy. Kidney Int. 2005, 67, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Kiryluk, K.; Novak, J.; Moldoveanu, Z.; Herr, A.B.; Renfrow, M.B.; Wyatt, R.J.; Scolari, F.; Mestecky, J.; Gharavi, A.G.; et al. The Pathophysiology of IgA Nephropathy. J. Am. Soc. Nephrol. 2011, 22, 1795–1803. [Google Scholar] [CrossRef] [Green Version]
- Crago, S.S.; Kutteh, W.H.; Moro, I.; Allansmith, M.R.; Radl, J.; Haaijman, J.J.; Mestecky, J. Distribution of IgA1-, IgA2-, and J chain-containing cells in human tissues. J. Immunol. 1984, 132, 16–18. [Google Scholar]
- Kett, K.; Brandtzaeg, P.; Radl, J.; Haaijman, J.J. Different subclass distribution of IgA-producing cells in human lymphoid organs and various secretory tissues. J. Immunol. 1986, 136, 3631–3635. [Google Scholar]
- Brandtzaeg, P.; Baekkevold, E.S.; Farstad, I.N.; Jahnsen, F.L.; Johansen, F.-E.; Nilsen, E.M.; Yamanaka, T. Regional specialization in the mucosal immune system: What happens in the microcompartments? Immunol. Today 1999, 20, 141–151. [Google Scholar] [CrossRef]
- Fukuyama, S.; Hiroi, T.; Yokota, Y.; Rennert, P.D.; Yanagita, M.; Kinoshita, N.; Terawaki, S.; Shikina, T.; Yamamoto, M.; Kurono, Y.; et al. Initiation of NALT Organogenesis Is Independent of the IL-7R, LTβR, and NIK Signaling Pathways but Requires the Id2 Gene and CD3−CD4+CD45+ Cells. Immunity 2002, 17, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Kiyono, H.; Fukuyama, S. NALT- versus PEYER’S-patch-mediated mucosal immunity. Nat. Rev. Immunol. 2004, 4, 699–710. [Google Scholar] [CrossRef]
- Hricik, D.E.; Chung-Park, M.; Sedor, J.R. Glomerulonephritis. N. Engl. J. Med. 1998, 339, 888–899. [Google Scholar] [CrossRef]
- Pawar, R.D.; Patole, P.S.; Wornle, M.; Anders, H.-J. Microbial nucleic acids pay a Toll in kidney disease. Am. J. Physiol. Physiol. 2006, 291, F509–F516. [Google Scholar] [CrossRef] [Green Version]
- Piccolo, M.; De Angelis, M.; Lauriero, G.; Montemurno, E.; Di Cagno, R.; Gesualdo, L.; Gobbetti, M. Salivary Microbiota Associated with Immunoglobulin A Nephropathy. Microb. Ecol. 2015, 70, 557–565. [Google Scholar] [CrossRef]
- Watanabe, H.; Goto, S.; Mori, H.; Higashi, K.; Hosomichi, K.; Aizawa, N.; Takahashi, N.; Tsuchida, M.; Suzuki, Y.; Yamada, T.; et al. Comprehensive microbiome analysis of tonsillar crypts in IgA nephropathy. Nephrol. Dial. Transplant. 2017, 32, 2072–2079. [Google Scholar] [CrossRef]
- Cao, Y.; Qiao, M.; Tian, Z.; Yu, Y.; Xu, B.; Lao, W.; Ma, X.; Li, W. Comparative Analyses of Subgingival Microbiome in Chronic Periodontitis Patients with and Without IgA Nephropathy by High Throughput 16S rRNA Sequencing. Cell. Physiol. Biochem. 2018, 47, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Iliescu, E.L.; Mercan-Stanciu, A.; Toma, L. Safety and efficacy of direct-acting antivirals for chronic hepatitis C in patients with chronic kidney disease. BMC Nephrol. 2020, 21, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandtzaeg, P. Potential of Nasopharynx-associated lymphoid tissue for vaccine responses in the airways. Am. J. Respir. Crit. Care Med. 2011, 183, 1595–1604. [Google Scholar] [CrossRef] [PubMed]
- Korsrud, F.R.; Brandtzaeg, P. Immunohistochemical Evaluation of J-Chain Expression by Intra- and Extra-follicular Immunoglobulin-producing Human Tonsillar Cells. Scand. J. Immunol. 1981, 13, 271–280. [Google Scholar] [CrossRef]
- Iwatani, H.; Inoue, T.; Wada, Y.; Nagasawa, Y.; Yamamoto, R.; Iijima, H.; Takehara, T.; Imai, E.; Rakugi, H.; Isaka, Y. Quantitative change of IgA hinge O-glycan composition is a novel marker of therapeutic responses of IgA nephropathy. Biochem. Biophys. Res. Commun. 2012, 428, 339–342. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, A.J.; McCoy, K.D.; Johansen, F.-E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Cheng, O.Z.; Palaniyar, N. NET balancing: A problem in inflammatory lung diseases. Front. Immunol. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, D.; Suzuki, Y.; Kano, T.; Suzuki, H.; Matsuoka, J.; Yokoi, H.; Horikoshi, S.; Ikeda, K.; Tomino, Y. Tonsillar TLR9 expression and efficacy of tonsillectomy with steroid pulse therapy in IgA nephropathy patients. Nephrol. Dial. Transplant. 2012, 27, 1090–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, J.; Suzuki, Y.; Suzuki, H.; Sato, D.; Kano, T.; Yanagawa, H.; Matsuzaki, K.; Horikoshi, S.; Novak, J.; Tomino, Y. Changes in Nephritogenic Serum Galactose-Deficient IgA1 in IgA Nephropathy following Tonsillectomy and Steroid Therapy. PLoS ONE 2014, 9, e89707. [Google Scholar] [CrossRef]
- Horie, A.; Hiki, Y.; Odani, H.; Yasuda, Y.; Takahashi, M.; Kato, M.; Iwase, H.; Kobayashi, Y.; Nakashima, I.; Maeda, K. IgA1 molecules produced by tonsillar lymphocytes are under-O-glycosylated in IgA nephropathy. Am. J. Kidney Dis. 2003, 42, 486–496. [Google Scholar] [CrossRef]
- Inoue, T.; Sugiyama, H.; Hiki, Y.; Takiue, K.; Morinaga, H.; Kitagawa, M.; Maeshima, Y.; Fukushima, K.; Nishizaki, K.; Akagi, H.; et al. Differential expression of glycogenes in tonsillar B lymphocytes in association with proteinuria and renal dysfunction in IgA nephropathy. Clin. Immunol. 2010, 136, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Peng, Y.; Liu, H.; Yin, W.; Chen, X.; Peng, X.; Shao, J.; Liu, Y.; Liu, F. Activation of the Interleukin-4/Signal Transducer and Activator of Transcription 6 Signaling Pathway and Homeodomain-Interacting Protein Kinase 2 Production by Tonsillar Mononuclear Cells in IgA Nephropathy. Am. J. Nephrol. 2013, 38, 321–332. [Google Scholar] [CrossRef]
- Chen, X.; Liu, H.; Peng, Y.; He, L.; Zhang, Y.; Xie, Y.; Peng, X.; Liu, C.; Liu, F. Expression and correlation analysis of IL-4, IFN-γ and FcαRI in tonsillar mononuclear cells in patients with IgA nephropathy. Cell. Immunol. 2014, 289, 70–75. [Google Scholar] [CrossRef]
- Van den Wall Bake, A.; Daha, M.R.; Evers-Schouten, J.; van Es, L.A. Serum IgA and the Production of IgA by peripheral blood and bone marrow lymphocytes in patients with primary iga nephropathy: Evidence for the bone marrow as the source of mesangial IgA. Am. J. Kidney Dis. 1988, 12, 410–414. [Google Scholar] [CrossRef]
- van den Wall Bake, A.W.; Daha, M.R.; Radl, J.; Haaijman, J.J.; Van der Ark, A.; Valentijn, R.M.; Van Es, L.A. The bone marrow as production site of the IgA deposited in the kidneys of patients with IgA nephropathy. Clin. Exp. Immunol. 1988, 72, 321–325. [Google Scholar]
- Harper, S.J.; Allen, A.C.; Pringle, J.H.; Feehally, J. Increased dimeric IgA producing B cells in the bone marrow in IgA nephropathy determined by in situ hybridisation for J chain mRNA. J. Clin. Pathol. 1996, 49, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Rickert, R.C.; Jellusova, J.; Miletic, A.V. Signaling by the tumor necrosis factor receptor superfamily in B-cell biology and disease. Immunol. Rev. 2011, 244, 115–133. [Google Scholar] [CrossRef]
- McCarthy, D.D.; Kujawa, J.; Wilson, C.; Papandile, A.; Poreci, U.; Porfilio, E.A.; Ward, L.; Lawson, M.A.; MacPherson, A.J.; McCoy, K.D.; et al. Mice overexpressing BAFF develop a commensal flora–dependent, IgA-associated nephropathy. J. Clin. Investig. 2011, 121, 3991–4002. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Suzuki, Y.; Yamanaka, T.; Hirose, S.; Nishimura, H.; Toei, J.; Horikoshi, S.; Tomino, Y. Genome-Wide Scan in a Novel IgA Nephropathy Model Identifies a Susceptibility Locus on Murine Chromosome 10, in a Region Syntenic to HumanIGAN1on Chromosome 6q22–23. J. Am. Soc. Nephrol. 2005, 16, 1289–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muto, M.; Manfroi, B.; Suzuki, H.; Joh, K.; Nagai, M.; Wakai, S.; Righini, C.; Maiguma, M.; Izui, S.; Tomino, Y.; et al. Toll-Like Receptor 9 Stimulation Induces Aberrant Expression of a Proliferation-Inducing Ligand by Tonsillar Germinal Center B Cells in IgA Nephropathy. J. Am. Soc. Nephrol. 2017, 28, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feehally, J.; Coppo, R.; Troyanov, S.; Bellur, S.S.; Cattran, D.; Cook, T.; Roberts, I.S.; Verhave, J.C.; Camilla, R.; Vergano, L.; et al. Tonsillectomy in a European Cohort of 1,147 Patients with IgA Nephropathy. Nephron 2016, 132, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, A.; Codognotto, M.; Tabbi, M.-G.; Favaro, E.; Rossi, B. Influence of tonsillectomy on the progression of mesangioproliferative glomerulonephritis. Nephrol. Dial. Transplant. 2010, 25, 2583–2589. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Anders, R.A.; Wu, Q.; Peng, D.; Cho, J.H.; Sun, Y.; Karaliukas, R.; Kang, H.-S.; Turner, J.R.; Fu, Y.-X. Dysregulated LIGHT expression on T cells mediates intestinal inflammation and contributes to IgA nephropathy. J. Clin. Investig. 2004, 113, 826–835. [Google Scholar] [CrossRef]
- Kano, T.; Suzuki, H.; Makita, Y.; Fukao, Y.; Suzuki, Y. Nasal-associated lymphoid tissue is the major induction site for nephritogenic IgA in murine IgA nephropathy. Kidney Int. 2021, 100, 364–376. [Google Scholar] [CrossRef]
- Inoue, T.; Iijima, H.; Tajiri, M.; Shinzaki, S.; Shiraishi, E.; Hiyama, S.; Mukai, A.; Nakajima, S.; Iwatani, H.; Nishida, T.; et al. Deficiency of N-acetylgalactosamine in O-linked oligosaccharides of IgA is a novel biologic marker for Crohnʼs disease. Inflamm. Bowel Dis. 2012, 18, 1723–1734. [Google Scholar] [CrossRef]
- Helin, H.; Mustonen, J.; Reunala, T.; Pasternack, A. IgA nephropathy associated with celiac disease and dermatitis herpeti-formis. Arch. Pathol. Lab. Med. 1983, 107, 324–327. [Google Scholar]
- Katz, A.; Dyck, R.F.; Bear, R.A. Celiac disease associated with immune complex glomerulonephritis. Clin. Nephrol. 1979, 11, 39–44. [Google Scholar]
- Coppo, R.; Basolo, B.; Rollino, C.; Roccatello, D.; Martina, G.; Amore, A.; Piccoli, G. Dietary Gluten and Primary IgA Nephropathy. N. Engl. J. Med. 1986, 315, 1167–1168. [Google Scholar] [CrossRef]
- Coppo, R.; Basolo, B.; Rollino, C.; Roccatello, D.; Martina, G.; Amore, A.; Bongiorno, G.; Piccoli, G. Mediterranean diet and primary IgA nephropathy. Clin. Nephrol. 1986, 26, 72–82. [Google Scholar] [PubMed]
- Coppo, R.; Amore, A.; Roccatello, D.; Gianoglio, B.; Molino, A.; Piccoli, G.; Clarkson, A.R.; Woodroffe, A.J.; Sakai, H.; Tomino, Y. IgA antibodies to dietary antigens and lectin-binding iga in sera from italian, australian, and japanese IgA Nephropathy Patients. Am. J. Kidney Dis. 1991, 17, 480–487. [Google Scholar] [CrossRef]
- Coppo, R.; Mazzucco, G.; Martina, G.; Roccatello, D.; Amore, A.; Novara, R.; Bargoni, A.; Piccoli, G.; Sena, L.M. Gluten-induced experimental IgA glomerulopathy. Lab. Investig. J. Tech. Methods Pathol. 1989, 60, 499–506. [Google Scholar]
- Coppo, R.; Amore, A.; Roccatello, D. Dietary antigens and primary immunoglobulin A nephropathy. J. Am. Soc. Nephrol. 1992, 2, S173. [Google Scholar] [CrossRef]
- Papista, C.; Lechner, S.; Ben Mkaddem, S.; LeStang, M.-B.; Abbad, L.; Bex-Coudrat, J.; Pillebout, E.; Chemouny, J.M.; Jablonski, M.; Flamant, M.; et al. Gluten exacerbates IgA nephropathy in humanized mice through gliadin–CD89 interaction. Kidney Int. 2015, 88, 276–285. [Google Scholar] [CrossRef]
- Coppo, R.; Roccatello, D.; Amore, A.; Quattrocchio, G.; Molino, A.; Gianoglio, B.; Amoroso, A.; Bajardi, P.; Piccoli, G. Effects of a gluten-free diet in primary IgA nephropathy. Clin. Nephrol. 1990, 33, 72–86. [Google Scholar]
- Moeller, S.; Canetta, P.A.; Taylor, A.K.; Arguelles-Grande, C.; Snyder, H.; Green, P.H.; Kiryluk, K.; Alaedini, A. Lack of Serologic Evidence to Link IgA Nephropathy with Celiac Disease or Immune Reactivity to Gluten. PLoS ONE 2014, 9, e94677. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Zhong, X.; Fan, J.M.; Zhang, Y.J.; Liu, X.R.; Ma, X.Y. External suppression causes the low expression of the Cosmc gene in IgA nephropathy. Nephrol. Dial. Transplant. 2008, 23, 1608–1614. [Google Scholar] [CrossRef] [Green Version]
- Blaas, S.H.; Stieber-Gunckel, M.; Falk, W.; Obermeier, F.; Rogler, G. CpG-oligodeoxynucleotides stimulate immunoglobulin A secretion in intestinal mucosal B cells. Clin. Exp. Immunol. 2009, 155, 534–540. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Montemurno, E.; Piccolo, M.; Vannini, L.; Lauriero, G.; Maranzano, V.; Gozzi, G.; Serrazanetti, D.I.; Dalfino, G.; Gobbetti, M.; et al. Microbiota and Metabolome Associated with Immunoglobulin A Nephropathy (IgAN). PLoS ONE 2014, 9, e99006. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Du, J.; Xie, Y.; Huang, Q.; Xiao, Y.; Chen, J.; Yan, S.; Gong, Z.; Ouyang, S. Fecal microbiota characteristics of Chinese patients with primary IgA nephropathy: A cross-sectional study. BMC Nephrol. 2020, 21, 97. [Google Scholar] [CrossRef] [Green Version]
- Kiryluk, K.; Li, Y.; Scolari, F.; Sanna-Cherchi, S.; Choi, M.; Verbitsky, M.; Fasel, D.; Lata, S.; Prakash, S.; Shapiro, S.; et al. Discovery of new risk loci for IgA nephropathy implicates genes involved in immunity against intestinal pathogens. Nat. Genet. 2014, 46, 1187–1196. [Google Scholar] [CrossRef]
- Smerud, H.K.; Bárány, P.; Lindström, K.; Fernström, A.; Sandell, A.; Påhlsson, P.; Fellström, B. New treatment for IgA nephropathy: Enteric budesonide targeted to the ileocecal region ameliorates proteinuria. Nephrol. Dial. Transplant. 2011, 26, 3237–3242. [Google Scholar] [CrossRef] [Green Version]
- Fellström, B.C.; Barratt, J.; Cook, H.; Coppo, R.; Feehally, J.; de Fijter, J.W.; Floege, J.; Hetzel, G.; Jardine, A.G.; Locatelli, F.; et al. Targeted-release budesonide versus placebo in patients with IgA nephropathy (NEFIGAN): A double-blind, randomised, placebo-controlled phase 2b trial. Lancet 2017, 389, 2117–2127. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R. The Gut-Renal Connection in IgA Nephropathy. Semin. Nephrol. 2018, 38, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.C.; Berthelot, L. Role of gut–kidney axis in renal diseases and IgA nephropathy. Curr. Opin. Gastroenterol. 2021, 37, 565–571. [Google Scholar] [CrossRef]
- Mertowska, P.; Mertowski, S.; Wojnicka, J.; Korona-Głowniak, I.; Grywalska, E.; Błażewicz, A.; Załuska, W. A Link between Chronic Kidney Disease and Gut Microbiota in Immunological and Nutritional Aspects. Nutrients 2021, 13, 3637. [Google Scholar] [CrossRef]
- Rehnberg, J.; Symreng, A.; Ludvigsson, J.F.; Emilsson, L. Inflammatory Bowel Disease Is More Common in Patients with IgA Nephropathy and Predicts Progression of ESKD: A Swedish Population-Based Cohort Study. J. Am. Soc. Nephrol. 2020, 32, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- Misaki, T.; Naka, S.; Kuroda, K.; Nomura, R.; Shiooka, T.; Naito, Y.; Suzuki, Y.; Yasuda, H.; Isozaki, T.; Nakano, K. Distribution of Streptococcus mutans strains with collagen-binding proteins in the oral cavity of IgA nephropathy patients. Clin. Exp. Nephrol. 2015, 19, 844–850. [Google Scholar] [CrossRef]
- Misaki, T.; Naka, S.; Hatakeyama, R.; Fukunaga, A.; Nomura, R.; Isozaki, T.; Nakano, K. Presence of Streptococcus mutans strains harbouring the cnm gene correlates with dental caries status and IgA nephropathy conditions. Sci. Rep. 2016, 6, 36455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Okamoto, K.; Kagami, A.; Yamamoto, Y.; Igarashi, T.; Kizaki, H. Streptococcus mutans Strains Harboring Collagen-binding Adhesin. J. Dent. Res. 2004, 83, 534–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, R.; Naka, S.; Nemoto, H.; Inagaki, S.; Taniguchi, K.; Ooshima, T.; Nakano, K. Potential involvement of collagen-binding proteins ofStreptococcus mutansin infective endocarditis. Oral Dis. 2013, 19, 387–393. [Google Scholar] [CrossRef]
- Nomura, R.; Naka, S.; Nemoto, H.; Otsugu, M.; Nakamura, S.; Ooshima, T.; Nakano, K. Potential high virulence for infective endocarditis in Streptococcus mutans strains with collagen-binding proteins but lacking PA expression. Arch. Oral Biol. 2013, 58, 1627–1634. [Google Scholar] [CrossRef]
- Misaki, T.; Naka, S.; Wato, K.; Hatakeyama, R.; Nagasawa, Y.; Ito, S.; Inaba, H.; Nomura, R.; Matsumoto-Nakano, M.; Nakano, K. Campylobacter rectus in the Oral Cavity Correlates with Proteinuria in Immunoglobulin A Nephropathy Patients. Nephron 2018, 139, 143–149. [Google Scholar] [CrossRef]
- Ito, S.; Misaki, T.; Naka, S.; Wato, K.; Nagasawa, Y.; Nomura, R.; Otsugu, M.; Matsumoto-Nakano, M.; Nakano, K.; Kumagai, H.; et al. Specific strains of Streptococcus mutans, a pathogen of dental caries, in the tonsils, are associated with IgA nephropathy. Sci. Rep. 2019, 9, 20130. [Google Scholar] [CrossRef]
- Naka, S.; Wato, K.; Misaki, T.; Ito, S.; Nagasawa, Y.; Nomura, R.; Matsumoto-Nakano, M.; Nakano, K. Intravenous administration of Streptococcus mutans induces IgA nephropathy-like lesions. Clin. Exp. Nephrol. 2020, 24, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Naka, S.; Wato, K.; Misaki, T.; Ito, S.; Matsuoka, D.; Nagasawa, Y.; Nomura, R.; Matsumoto-Nakano, M.; Nakano, K. Streptococcus mutans induces IgA nephropathy-like glomerulonephritis in rats with severe dental caries. Sci. Rep. 2021, 11, 5784. [Google Scholar] [CrossRef] [PubMed]
- Stinson, M.W.; Nisengard, R.J.; Neiders, M.E.; Albini, B. Serology and tissue lesions in rabbits immunized with Streptococcus mutans. J. Immunol. 1983, 131, 3021–3027. [Google Scholar] [PubMed]
- Albini, B.; Nisengard, R.J.; Glurich, I.; Neiders, M.E.; Stinson, M.W. Streptococcus mutans-induced nephritis in rabbits. Am. J. Pathol. 1985, 118, 408–418. [Google Scholar]
- Miyata, M.; Glurich, I.; Kasukawa, R.; Milgrom, F.; Stinson, M.; Albini, B. Streptococcus-mutans-Induced Nephritis in Rabbits: Rheumatoid Factors and Nephritogenicity. Int. Arch. Allergy Immunol. 1995, 108, 360–367. [Google Scholar] [CrossRef]
- Muyzer, G.; Brinkhoff, T.; Nübel, U.; Santegoeds, C.; Schäfer, H.; Wawer, C. Denaturant Gradient Gel Electrophoresis in Microbial Ecology. In Molecular Microbial Ecology Manual; Akkermans, A., Van Elsas, J.D., De Bruijin, F., Eds.; Springer: New York, NY, USA, 1998; Volume 3.4.4. [Google Scholar]
- Nagasawa, Y.; Iio, K.; Fukuda, S.; Date, Y.; Iwatani, H.; Yamamoto, R.; Horii, A.; Inohara, H.; Imai, E.; Nakanishi, T.; et al. Periodontal Disease Bacteria Specific to Tonsil in IgA Nephropathy Patients Predicts the Remission by the Treatment. PLoS ONE 2014, 9, e81636. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Suzuki, H.; Nishida, T.; Mishima, M.; Tachibana, M.; Fujishima, M.; Shimizu, T.; Watarai, M. Identification of novel Legionella genes required for endosymbiosis in Paramecium based on comparative genome analysis with Holospora spp. FEMS Microbiol. Ecol. 2018, 94, fiy162. [Google Scholar] [CrossRef]
- Gao, L.; Xu, T.; Huang, G.; Jiang, S.; Gu, Y.; Chen, F. Oral microbiomes: More and more importance in oral cavity and whole body. Protein Cell 2018, 9, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Khasnobish, A.; Takayasu, L.; Watanabe, K.-I.; Nguyen, T.T.T.; Arakawa, K.; Hotta, O.; Joh, K.; Nakano, A.; Hosomi, S.; Hattori, M.; et al. Dysbiosis in the salivary microbiome associated with IgA Nephropathy—A Japanese cohort study. Microbes Environ. 2021, 36, ME21006. [Google Scholar] [CrossRef]
- Nagasawa, Y.; Nomura, R.; Misaki, T.; Ito, S.; Naka, S.; Wato, K.; Okunaka, M.; Watabe, M.; Fushimi, K.; Tsuzuki, K.; et al. Relationship between IgA nephropathy and Porphyromonas gingivalis; red complex of perio-dontopathic bacterial species. Int. J. Mol. Sci. 2021, 22, 13022. [Google Scholar] [CrossRef]
- Nomura, R.; Nagasawa, Y.; Misaki, T.; Ito, S.; Naka, S.; Wato, K.; Okunaka, M.; Watabe, M.; Tsuzuki, K.; Matsumoto-Nakano, M.; et al. Relationship of distribution of periodontopathic bacterial species between saliva and tonsils. BMC Oral Health, 2022; in press. [Google Scholar]
- Nagasawa, Y.; Yamamoto, R.; Rakugi, H.; Isaka, Y. Cigarette smoking and chronic kidney diseases. Hypertens. Res. 2012, 35, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasawa, Y.; Kida, A.; Nakanisihi, T. Effect of cigarette smoking cessation on CKD: Is it a cancer-suppression-like effect or a CVD-suppression-like effect? Hypertens. Res. 2016, 39, 690–691. [Google Scholar] [CrossRef]
- Sharma, S.; Mishra, S.K.; Mittal, N. Influence of tobacco dependence on caries development in young male adults: A cross-sectional study. J. Conserv. Dent. 2018, 21, 597–601. [Google Scholar] [CrossRef]
- Jiang, X.; Jiang, X.; Wang, Y.; Huang, R. Correlation between tobacco smoking and dental caries: A systematic review and meta-analysis. Tob. Induc. Dis. 2019, 17, 34. [Google Scholar] [CrossRef]
- Leite, F.R.M.; Nascimento, G.G.; Scheutz, F.; López, R. Effect of smoking on periodontitis: A systematic review and meta-regression. Am. J. Prev. Med. 2018, 54, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Anis, K.H.; Weinrauch, L.A.; D’Elia, J.A. Effects of smoking on solid organ transplantation outcomes. Am. J. Med. 2019, 132, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Koshino, K.; Ushigome, H.; Sakai, K.; Suzuki, T.; Nobori, S.; Okajima, H.; Masuzawa, N.; Yoshimura, N. Outcome of tonsillectomy for recurrent IgA nephropathy after kidney transplantation. Clin. Transplant. 2013, 27, 22–28. [Google Scholar] [CrossRef]
- Sato, Y.; Ishida, H.; Shimizu, T.; Tanabe, K. Evaluation of tonsillectomy before kidney transplantation in patients with IgA nephropathy. Transpl. Immunol. 2014, 30, 12–17. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, Z.; Zhang, H.; Liu, X. Aberrant IgA1 Glycosylation in IgA Nephropathy: A Systematic Review. PLoS ONE 2016, 11, e0166700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H. Biomarkers for IgA nephropathy on the basis of multi-hit pathogenesis. Clin. Exp. Nephrol. 2019, 23, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Malycha, F.; Eggermann, T.; Hristov, M.; Schena, F.P.; Mertens, P.R.; Zerres, K.; Floege, J.; Eitner, F. No evidence for a role of cosmc-chaperone mutations in European IgA nephropathy patients. Nephrol. Dial. Transplant. 2009, 24, 321–324. [Google Scholar] [CrossRef] [Green Version]
- Serino, G.; Sallustio, F.; Curci, C.; Cox, S.N.; Pesce, F.; De Palma, G.; Schena, F.P.; Keyzer, C.A.; De Jong, M.A.; Van Breda, G.F.; et al. Role of let-7b in the regulation ofN-acetylgalactosaminyltransferase 2 in IgA nephropathy. Nephrol. Dial. Transplant. 2015, 30, 1132–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serino, G.; Sallustio, F.; Cox, S.N.; Pesce, F.; Schena, F.P. Abnormal miR-148b Expression Promotes Aberrant Glycosylation of IgA1 in IgA Nephropathy. J. Am. Soc. Nephrol. 2012, 23, 814–824. [Google Scholar] [CrossRef] [Green Version]
- Kouri, N.M.; Stangou, M.; Lioulios, G.; Mitsoglou, Z.; Serino, G.; Chiurlia, S.; Cox, S.N.; Stropou, P.; Schena, F.P.; Papagianni, A. Serum Levels of miR-148b and Let-7b at diagnosis may have important impact in the response to treatment and long-term outcome in IgA Nephropathy. J. Clin. Med. 2021, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Lan, J.; Zhang, H.; Zhou, N.; Liang, Y.; Liu, X. MicroRNA-196b targets COSMC in pediatric IgA nephropathy. Mol. Med. Rep. 2020, 21, 2260–2266. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, J.; Zhao, Y. MiR-320 promotes B cell proliferation and the production of aberrant glycosylated IgA1 in IgA nephropathy. J. Cell. Biochem. 2018, 119, 4607–4614. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Bao, H.; Xu, X.; Zhou, X.; Qin, W.; Zeng, C.; Liu, Z. Increased miR-374b promotes cell proliferation and the production of aberrant glycosylated IgA1 in B cells of IgA nephropathy. FEBS Lett. 2015, 589, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, X.; Peng, W.; Wei, M.; Qin, W. MicroRNA-155-induced T lymphocyte subgroup drifting in IgA nephropathy. Int. Urol. Nephrol. 2017, 49, 353–361. [Google Scholar] [CrossRef]
- Liu, C.; Ye, M.-Y.; Yan, W.-Z.; Peng, X.-F.; He, L.-Y.; Peng, Y.-M. microRNA-630 regulates underglycosylated IgA1 production in the Tonsils by targeting TLR4 in IgA Nephropathy. Front. Immunol. 2020, 11, 563699. [Google Scholar] [CrossRef]
- Mogitate, M.; Sasaki, Y.; Komiyama, A. Outcome of an outpatient specialty clinic for chronic epipharyngitis. Auris Nasus Larynx 2021, 48, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Fan, R.; Zhang, Z.; Brown, R.; Hall, S.; Julian, B.A.; Chatham, W.W.; Suzuki, Y.; Wyatt, R.J.; Moldoveanu, Z.; et al. Aberrantly glycosylated IgA1 in IgA nephropathy patients is recognized by IgG antibodies with restricted heterogeneity. J. Clin. Investig. 2009, 119, 1668–1677. [Google Scholar] [CrossRef] [Green Version]
- Gharavi, A.G.; Yan, Y.; Scolari, F.; Schena, F.P.; Frasca, G.M.; Ghiggeri, G.M.; Cooper, K.; Amoroso, A.; Viola, B.F.; Battini, G.; et al. IgA nephropathy, the most common cause of glomerulonephritis, is linked to 6q22–23. Nat. Genet. 2000, 26, 354–357. [Google Scholar] [CrossRef]
- Yamamoto, R.; Nagasawa, Y.; Shoji, T.; Inoue, K.; Uehata, T.; Kaneko, T.; Okada, T.; Yamauchi, A.; Tsubakihara, Y.; Imai, E.; et al. A candidate gene approach to genetic prognostic factors of IgA nephropathy—A result of Polymorphism REsearch to DIstinguish genetic factors Contributing To progression of IgA Nephropathy (PREDICT-IgAN). Nephrol. Dial. Transplant. 2009, 24, 3686–3694. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, R.; Nagasawa, Y.; Shoji, T.; Katakami, N.; Ohtoshi, K.; Hayaishi-Okano, R.; Yamasaki, Y.; Yamauchi, A.; Tsubakihara, Y.; Imai, E.; et al. A candidate gene approach to genetic contributors to the development of IgA nephropathy. Nephrol. Dial. Transplant. 2012, 27, 1020–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Rodriguez, E.; Southard, C.T.; Kiryluk, K. GWAS-Based Discoveries in IgA nephropathy, membranous nephropathy, and steroid-sensitive nephrotic syndrome. Clin. J. Am. Soc. Nephrol. 2021, 16, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Ishihara, K.; Sugihara, N.; Kusumoto, M.; Yakushiji, M.; Okuda, K. Colonization pattern of periodontal bacteria in Japanese children and their mothers. J. Periodontal Res. 2008, 43, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagasawa, Y.; Misaki, T.; Ito, S.; Naka, S.; Wato, K.; Nomura, R.; Matsumoto-Nakano, M.; Nakano, K. Title IgA Nephropathy and Oral Bacterial Species Related to Dental Caries and Periodontitis. Int. J. Mol. Sci. 2022, 23, 725. https://doi.org/10.3390/ijms23020725
Nagasawa Y, Misaki T, Ito S, Naka S, Wato K, Nomura R, Matsumoto-Nakano M, Nakano K. Title IgA Nephropathy and Oral Bacterial Species Related to Dental Caries and Periodontitis. International Journal of Molecular Sciences. 2022; 23(2):725. https://doi.org/10.3390/ijms23020725
Chicago/Turabian StyleNagasawa, Yasuyuki, Taro Misaki, Seigo Ito, Shuhei Naka, Kaoruko Wato, Ryota Nomura, Michiyo Matsumoto-Nakano, and Kazuhiko Nakano. 2022. "Title IgA Nephropathy and Oral Bacterial Species Related to Dental Caries and Periodontitis" International Journal of Molecular Sciences 23, no. 2: 725. https://doi.org/10.3390/ijms23020725