1. Introduction
Worldwide prevalence of obesity is considered to be one of the main promoters for several common metabolic diseases such as diabetes, cardiovascular problems, and cancer [
1]. Reported links between obesity progression and the prevalence of various other diseases have become a big concern for public health. Obesity is characterized basically with inflammation caused by the hormonal secretions of excessive adipose tissue formation and consequent lipid accumulation in the body. Formation of the adipose tissue and accumulating lipid are results of overstimulated adipogenesis, which is an intricate differentiation process of pre-adipocytes involving several intertwined signaling pathways resulting in differentiation into mature adipocytes [
2]. Maturation of adipocytes is characterized with intracellular lipid accumulation and adipocyte-specific gene expressions. Initiation of the adipogenesis is strictly regulated by adipogenic transcription factors of which peroxisome proliferator activated receptor gamma (PPARγ) and CCAAT/enhancer binding proteins (C/EBP) are the key factors [
3]. Expression and transcriptional activities of these adipogenic factors, as well as their downstream effector proteins, are mandatory for the progression of adipogenesis.
In multipotent mesenchymal stromal cells (MSCs), increased adipocyte formation as a result of adipogenic differentiation promotes a set of bone disorders, osteoporosis being the most prevalent one [
4,
5]. Adipogenic tendencies of the bone marrow MSCs against osteogenic differentiation are the main causes of sponge-like bone formation seen in osteoporotic conditions. Several reports have suggested that wingless-type MMTV integration site (Wnt)/β-catenin is the positive regulator for the osteogenic differentiation of bone marrow MSCs, simultaneously suppressing the adipocyte differentiation [
6]. In healthy individuals, negative regulation of adipogenesis via canonical Wnt/β-catenin pathway keeps the balance between adipocyte and osteocytes in bone structure. As the downstream effector of this canonical pathway, nuclear translocation of β-catenin negatively regulates the expression of PPARγ and C/EBPα, consequently inhibiting the adipogenesis [
7]. Therefore, regulation of PPARγ and Wnt/β-catenin signaling might be considered as a target to prevent or control obesity via hindering adiposity.
Natural bioactive substances have been gaining increasing attention due to their ability to regulate adipogenic differentiation. To date, several types of phytochemicals have been reported to possess adipogenesis inhibitory properties [
8]. Phenolic substances allocate a big portion of aforementioned bioactive phytochemicals.
Artemisia princeps, also known as Korean or Japanese mugwort, is a plant native to Asia and is reported to possess adipogenesis inhibitory properties in mouse 3T3-L1 pre-adipocytes among its other in vitro bioactivities such as antioxidant, antimicrobial, anti-inflammatory and antitumor [
9,
10,
11]. Several phenolic substances have been isolated from
A. princeps as an active component. Eupatilin and jaceosidin flavonoids inhibited inflammation in mice [
12] while 6-acetyl-2,2-dimethylchroman-4-one [
13] and loliolide [
14] inhibited the adipogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Dracunculin, which is the simple name given to 6-methoxy-7,8-methylenedioxycoumarin, is a known coumarin that can be found mainly in mugworts. It has been isolated mostly from
Artemisia species, tarragon (
Artemisia dracunculus) being the first, which has given this compound its common name, dracunculin. Although several other coumarins were isolated from
Artemisia species and their bioactivities have been reported, the literature lacks detailed information in terms of the health beneficial properties of dracunculin. Therefore, in this study, the potential anti-adipogenic effect of dracunculin (DCC) isolated from
A. princeps has been evaluated. To the best of our knowledge, this is the first report that exhibits the effect of DCC on adipogenic differentiation.
3. Discussion
DCC is a coumarin derivative, also named as 7,8-Methylenedioxy-6-methoxycoumarin [
18]. It has been known as a phytochemical for decades and is mainly isolated from
Artemisia spp. Although
Artemisia spp. are known to possess anti-adipogenic properties and DCC is a common phytochemical, the literature lacks studies on the bioactive potential of DCC. Like other coumarins similar to DCC in terms of structure, DCC is expected to possess some bioactive properties. To the best of our knowledge, the only bioactive property reported for DCC is its antitrypanosomal activity [
19]. The current study proposes, for the first time, that DCC possesses bioactive potential to suppress adipogenic differentiation of bone marrow cells in vitro by activation of Wnt/β-catenin and AMPK pathways.
Increased adipogenic differentiation of bone marrow cells is suggested to be one of the causes for obesity-related bone complications such as osteoporosis [
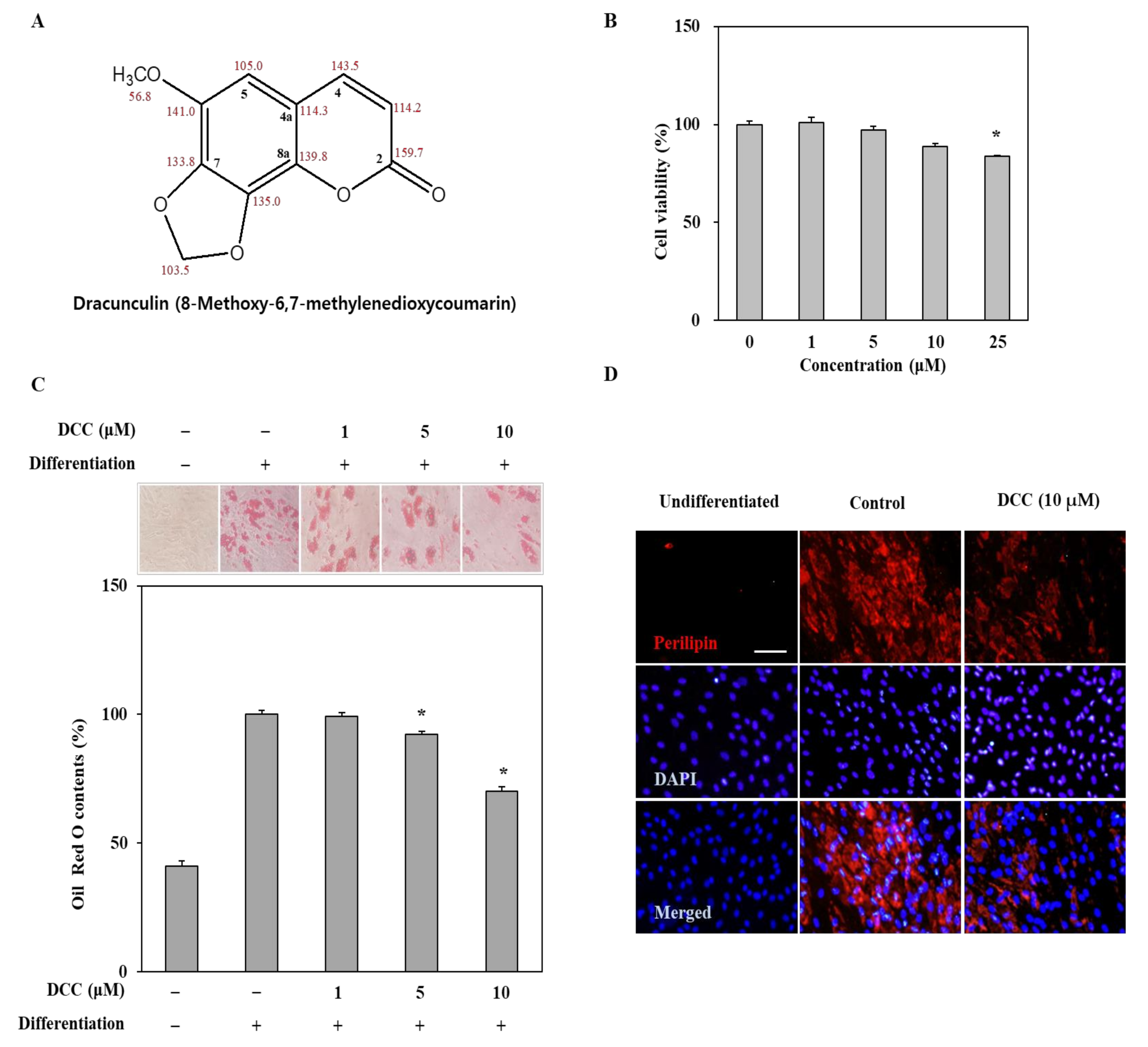
1]. Formation of adipocytes is characterized by accumulation of intracellular lipid droplets as a marker for mature adipocytes. In this study, DCC was shown to decrease the lipid accumulation of hBM-MSCs when they were induced to undergo adipogenesis compared with the untreated group that differentiated into mature adipocytes with substantial lipid accumulation. This result indicated that DCC treatment hindered adipogenesis at some point that the adipo-induced hBM-MSCs were not able to mature and accumulate lipid droplets.
The adipogenesis of mesenchymal stromal cells is regulated by a web of different signaling cascades and transcription factors [
3]. The main adipogenic transcription factors that initiate and progress the adipocyte formation are PPARγ and C/EBP-α. PPARγ activation can stimulate the beginning of adipogenesis by itself via a positive feedback loop and crosstalk between C/EBP-α and its upstream activators, C/EBP-β and C/EBP-δ [
20]. The adipogenic differentiation tendencies of hBM-MSCs, instead of osteoblastogenesis, are also regulated by this cascade very similarly to adipose tissue. Activation of this signaling cascade occurs at the early stages of adipogenic differentiation. Hence, in the current study, DCC was introduced to the cells during the initial induction of adipogenesis. The results showed that DCC inhibited the mRNA and protein levels of PPARγ and C/EBP-α, as well as adipocyte-specific SREBP1c, showing that DCC was able to suppress adipogenesis by interfering with the transcription factor signaling cascade.
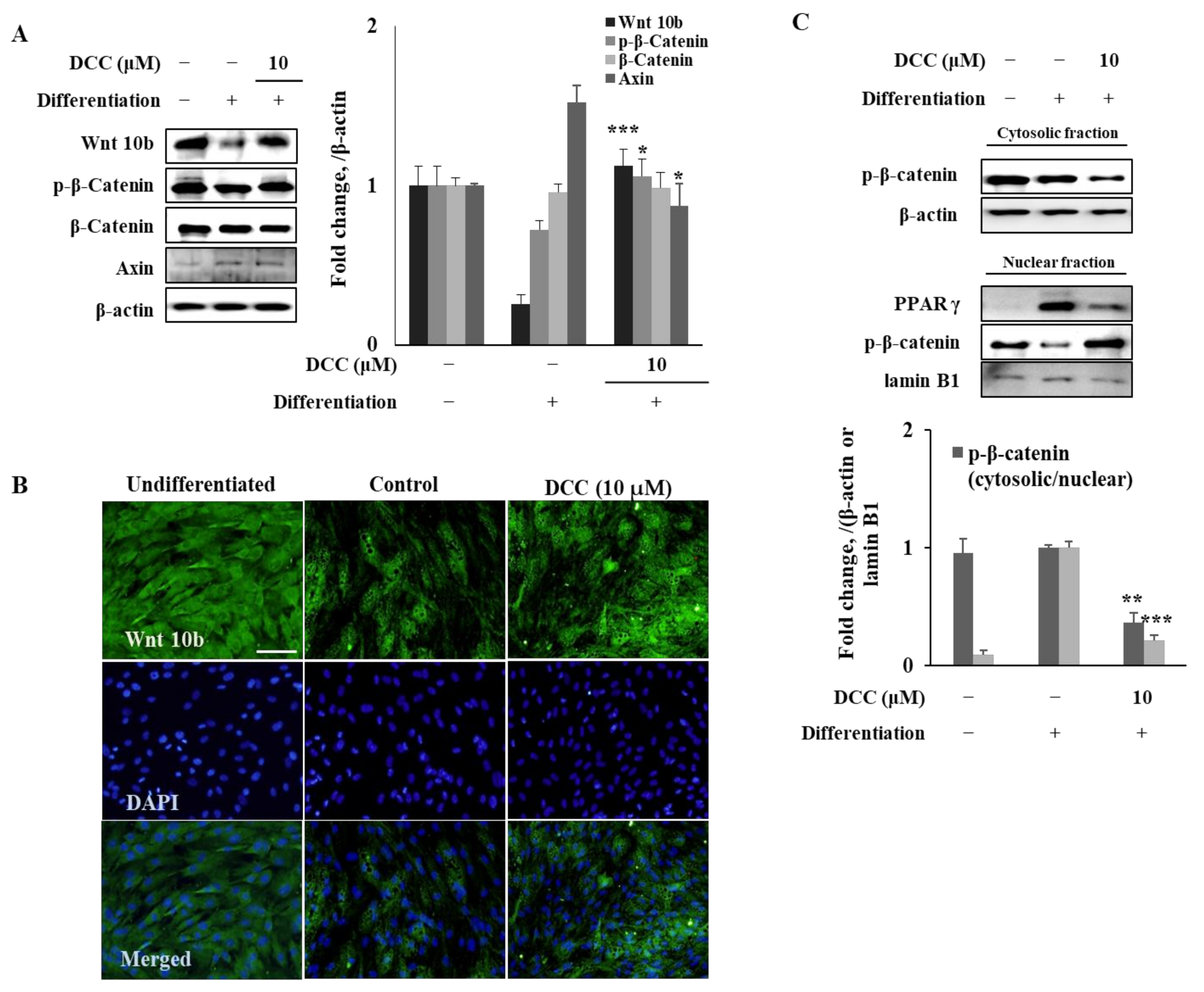
Another known regulator of adipogenesis is Wnt/β-catenin signaling, which downregulates PPARγ and C/EBPα cascade when activated [
6]. It has been shown that the activation of Wnt receptors negatively regulates the adipogenesis via suppression of the activity of PPARγ by nuclear translocation of β-catenin. Results showed that DCC treatment significantly increased the Wnt10b protein expression, which was suppressed in untreated adipocytes as an indicator of activated Wnt pathway. However, DCC was not able elevate the β-catenin protein levels as expected. This showed that DCC was able to activate the Wnt pathway in a non-canonical way. Therefore, Ser
552 phosphorylated β-catenin levels were examined. When phosphorylated at Ser
552 via AMPK, β-catenin is stabilized and accumulated in the nucleus, which consequently suppresses the PPARγ signaling [
15]. DCC treatment significantly increased Ser
552 phosphorylated β-catenin levels in adipo-induced hBM-MSCs but did not affect the total β-catenin levels, indicating that DCC was able to activate β-catenin transcriptional activity through AMPK. This was further supported by the nuclear levels of phosphorylated β-catenin and PPARγ. Adipocytes expressed a high amount of PPARγ compared with a minimal presence of phosphorylated β-catenin in the nucleus. However, DCC treatment reverted the nuclear protein profile of adipo-induced hBM-MSCs. DCC-treated cells showed increased levels of nuclear phosphorylated β-catenin and diminished PPARγ. This supported the suggestion that DCC hinders the adipogenesis via activated β-catenin signaling.
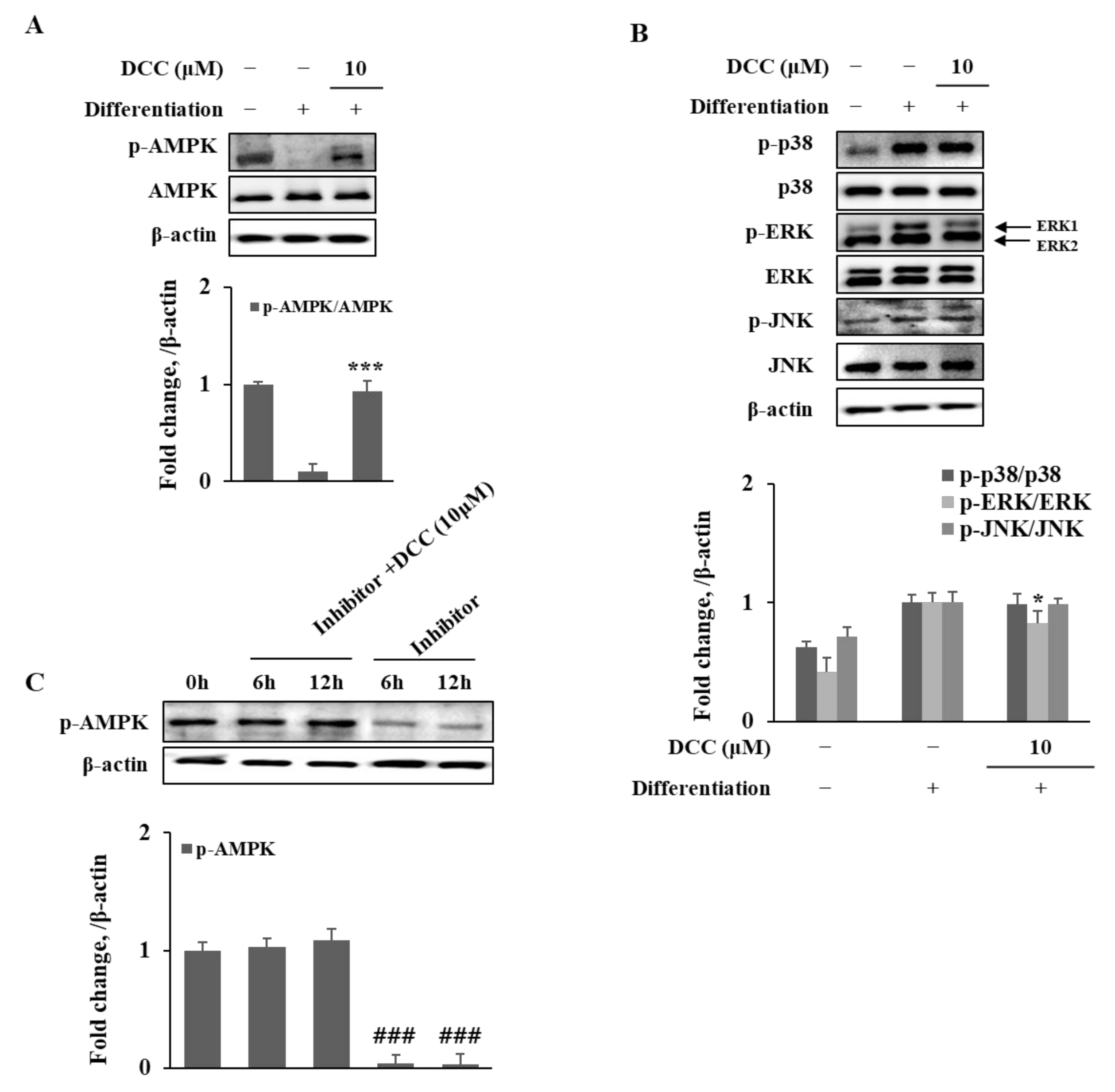
AMPK is also known as a negative regulator for adipogenesis via different ways, which include suppression of MAPK activation on ERK1/2 and activation of β-catenin via phosphorylation. The stimulation of AMPK activation by DCC treatment was shown in this study. The presence of DCC significantly increased the AMPK phosphorylation, which was suppressed during adipogenesis. In addition, DCC was able to revert the effects of AMPK inhibitor compound C, further suggesting strong AMPK activation activity for DCC. Abiola et al. [
21] suggested a reciprocal crosstalk between AMPK and Wnt10b activation. They reported that activation of Wnt/beta-catenin signaling by means of elevated Wnt10b protein in skeletal muscle cells resulted in activated AMPK pathway and suppressed MAPK activation. These results are consistent with the findings of the current study, which explain the Wnt10b stimulation by DCC treatment along with AMPK-mediated non-canonical β-catenin activation. However, future studies are needed to determine the detailed mechanism of action for DCC to show where and how DCC interferes with the crosstalk between AMPK and Wnt/β-catenin signaling, which resulted in suppressed adipogenesis. Another coumarin derivative, 6-acetyl-2,2-dimethylchroman-4-one, previously isolated from
A. princeps shares similarities with DCC in structure and was shown to inhibit adipogenic differentiation via similar pathways to further support the AMPK activating ability of DCC [
13].
In conclusion, DCC inhibited the adipogenic differentiation and lipid accumulation of hBM-MSCs through activation of AMPK-mediated Wnt/β-catenin signaling in vitro. Future studies are expected to further examine its potential to be utilized as a natural bioactive agent against bone marrow adiposity.
4. Materials and Methods
4.1. Materials
The isolation of DCC from
A. princeps and the chemical characterization of the DCC were carried out as reported [
22]. The NMR data of DCC used for identification is given in
Figure S1. The human bone marrow-derived mesenchymal stromal cell (hBM-MSC) line, and cell culture (Mesenchymal Stem Cell Growth Medium, C-28009) and differentiation medium (Mesenchymal Stem Cell Adipogenic Differentiation Medium 2, C-28016) were purchased from PromoCell (Heildelberg, Germany). Other reagents used in cell culture, maintenance and differentiation including phosphate buffer saline (PBS) were purchased from Gibco BRL (New York City, NY, USA) unless otherwise noted. 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), and isopropanol were purchased from Sigma-Aldrich (St. Louis, MO, USA). The AccuPrep Universal RNA extraction kit was purchased from Bioneer (Daejeon, Korea). The Cell Script All-in-One cDNA synthesis premix was obtained from CellSafe (Yongin, Korea). The NE-PER nuclear protein extraction kit, BCA protein assay kit for total protein quantification, and primary antibody against JNK were from Thermo Fisher Scientific (Waltham, MA, USA). Polyvinylidene fluoride membrane and enhanced chemiluminescence kit for the protein band detection were from Amersham Biosciences (Amersham, England, UK). Primary antibodies against PPARγ (#2443), CCAAT/enhancer-binding protein (C/EBP) α (#2295), p38 (#8690), phospho(p)-p38 (#4511), ERK (#4695), p-ERK (#4370), AMPK (#2603), and p-AMPK (#2531) were obtained from Cell Signal Technology (Danvers, MA, USA) while sterol regulatory element-binding protein 1c (SREBP1c) antibody (ab3259) was from Abcam (Cambridge, England, UK). Other primary secondary antibodies for Western blotting were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). For the immunocytochemical fluorescence staining, Alexa Fluor 488-conjugated secondary antibody (A-11008) was from Invitrogen (Waltham, MA, USA) while perilipin-1 (ab3526) and PPARγ (ab9256) primary antibodies were from Abcam. Solutions, buffers, and reagents (#12727) for the fluorescence staining were purchased from Cell Signal Technology.
4.2. Cell Culture and Differentiation
The hBM-MSCs were differentiated into adipocytes as previously reported [
14]. Briefly, hBM-MSCs were grown to confluence, and the culture medium was swapped with differentiation medium two days later (day 0). The differentiation medium was changed with a fresh one every third day until most of the cells exhibited mature adipocyte characteristics by accumulating lipid droplets on the twelfth day of differentiation (day 12). DCC was introduced to differentiating cells on day 0 until the next medium change (day 3). One group of differentiated cells were not treated with DCC as a control group while another group of cells were not induced to differentiate (kept with growth medium) nor treated with DCC as a blank group.
4.3. Assessment of Cytotoxicity
The cytotoxic concentrations of DCC were assessed by traditional MTT assay. The hBM-MSCs were grown in 96-well plates to approximately 80% confluency. Next, cells were treated with increasing concentrations of DCC in growth medium and incubated for 72 h. After incubation, growth medium was replaced with a fresh one containing 20 μL MTT (1 mg/mL) instead of DCC, and the cells were kept at 37 °C in a 5% CO2 incubator for 4 h. The wells were then aspirated and supplied with 100% DMSO. The absorbance values of the wells at 540 nm were measured on a MultiSkan GO microplate reader (Thermo Fisher Scientific). The cytotoxicity of DCC was measured as a relative percentage of cell viability compared to the untreated group.
4.4. Lipid Droplet Staining
The Oil Red O staining was employed to demonstrate the cellular lipid accumulation as a marker of mature adipocyte. On day 12, the differentiated hBM-MSCs were washed with PBS and fixed on 6-well plates by addition on 10% formaldehyde for 1 h. After 1 h, the wells were aspirated and supplied with 0.5% Oil Red O staining solution (wt/v, dissolved in 3:2 isopropanol and distilled water). Staining was carried out by keeping the cells at room temperature for 1 h, after which the wells were aspirated again, washed with PBS and photographed using a microscope. Quantification of the Oil Red O stain bound by cellular lipid droplets was carried out via elution of the stain from cells through addition of 100% isopropanol. The wells were then measured for their absorbance at 500 nm on a MultiSkan GO microplate reader. The lipid accumulation was given as the stain absorbance percentage relative to the differentiated untreated control group.
4.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
The reverse transcription-polymerase chain reaction was performed using the total RNA isolated from the hBM-MSCs on day 12. Total RNA was extracted from cells with an AccuPrep Universal RNA extraction kit. The cDNA was synthesized from equal amounts of total RNA (2 μg) from each well. Reverse transcription was performed through the application of Cell Script All-in-One cDNA synthesis kit using a T100 thermocycler (Bio-Rad, Hercules, USA) with 42 °C for 60 min and 72 °C for 5 min settings. This was followed by standard PCR analysis consisting of 30 cycles of denaturing at 95 °C for 45 s, annealing at 60 °C for 1 min and extending at 72 °C for 45 s. The PCR reaction was carried out via Lune Universal PCR mix and following its instructions. The primers for the PCR analysis used were given in a previous report [
14]. Finally, the PCR bands of the specific proteins were obtained after electrophoresing on 2% agarose gel and imaging with CAS-400SM imaging system (Davinch-K, Seoul, Korea).
4.6. Western Blot Analysis
The Western blot was carried out to detect the protein levels of specific proteins in hBM-MSCs on day 12. The homogenized cellular total protein lysates were obtained using RIPA buffer and vigorous pipetting. The lysates were then centrifuged at 4 °C for 15 min (13,000× g), and the supernatants were used for Western blot. Nuclear proteins were extracted from total cell lysates using a NE-PER nuclear extraction kit. The protein amounts of each well were assessed by BCA assay kit, and equal amounts of protein (20 μg) were loaded onto sodium dodecyl sulfate–polyacrylamide gel (4% stacking and 10% separating gels) for electrophoretic separation of the proteins. Next, the proteins were transferred to polyvinylidene fluoride membranes, which were then blocked with 5% skim milk in TBS-T buffer (v/v) for 1 h at room temperature. Blocked membranes were kept overnight at 4 °C with primary antibodies. Membranes were washed and hybridized with horseradish peroxidase-conjugated secondary antibodies. The hybridized protein bands were then visualized using enhanced chemiluminescence kit with a CAS-400SM imaging system.
4.7. Fluorescence STAINING
The cellular perilipin-1, PPARγ, and Wnt 10b proteins were visualized by immunofluorescence staining in hBM-MSCs on day 12. The hBM-MSCs were treated with the fixation, washing, and permeabilization reagents of the Immunofluorescence Application Solutions Kit according to its protocol. Next, cells were introduced to perilipin-1, PPARγ and Wnt10b primary antibodies. The cells were stained with Alexa Fluor 488-conjugated anti-rabbit secondary antibodies for visualization. For normalization, viable cells were also stained with DAPI (#8961; Cell Signaling Technology) containing ProLong Gold Antifade reagent. Stained cell images were taken using an Olympus fluorescence microscope equipped with a digital camera, and the images were then processed with the Multi Gauge software.
4.8. Statistical Analysis
The numerical data are given as mean of three separated results ± SD (n = 3). The statistically significant difference between two groups was decided according to the results of one-way analysis of variance followed by Duncan’s multiple range post hoc test. SAS (v9.1) software was used for statistical analysis and calculation of p-values. The meaningful significance was determined at p < 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}



