
Oocyte-Specific Knockout of Histone Lysine Demethylase KDM2a Compromises Fertility by Blocking the Development of Follicles and Oocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Confirmation of Kdm2a Deletion
2.2. Expression and Subcellular Localization of Kdm2a
2.3. Decrease the Fecundity of Kdm2a cKO Female Mice
2.4. Effects of Kdm2a on Follicular Development
2.5. Kdm2a cKO Females Exhibited Decreased Levels of Estradiol
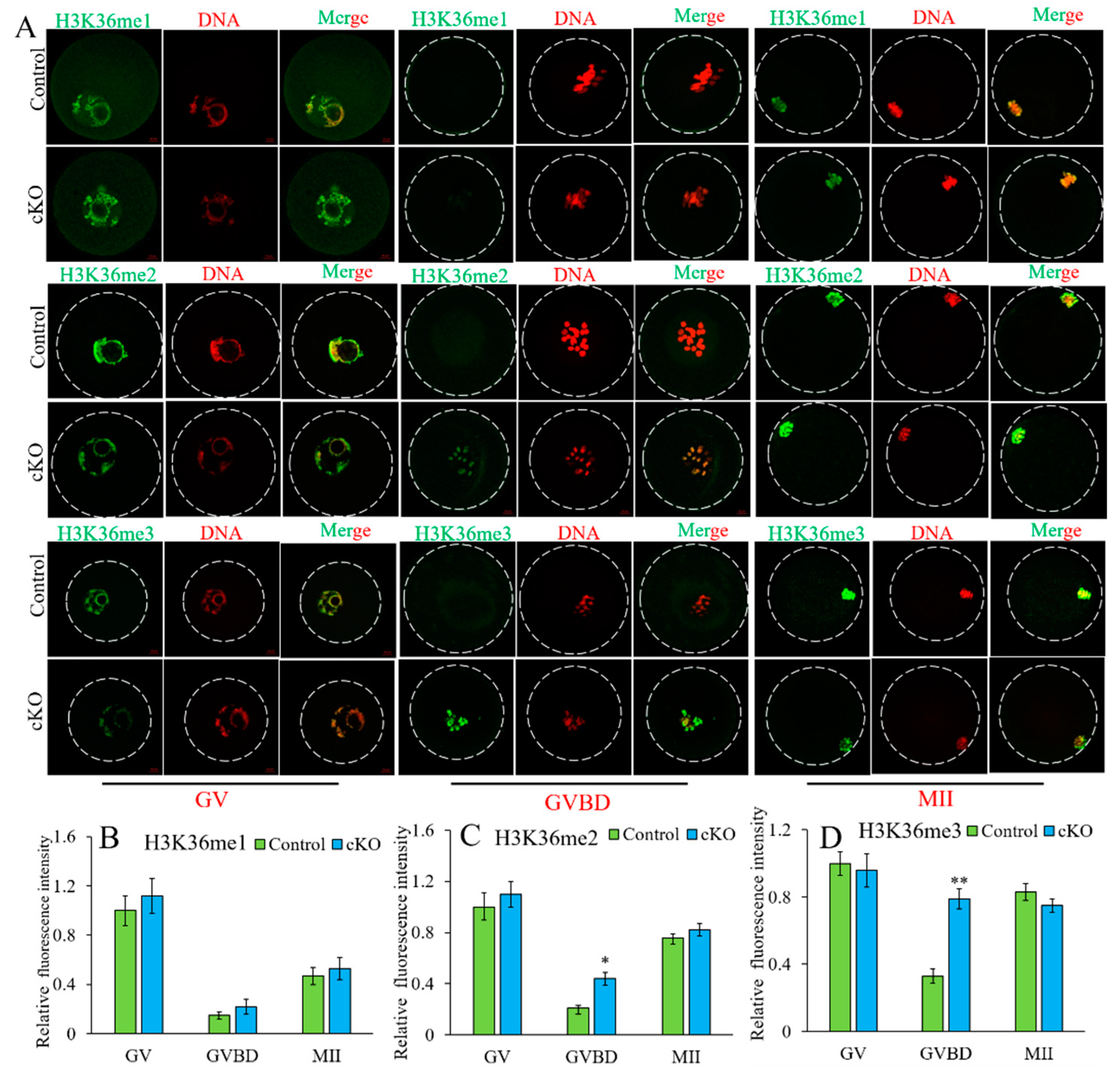
2.6. Kdm2a cKO Increased the Level of H3K36me2/3 during Oocyte Growth
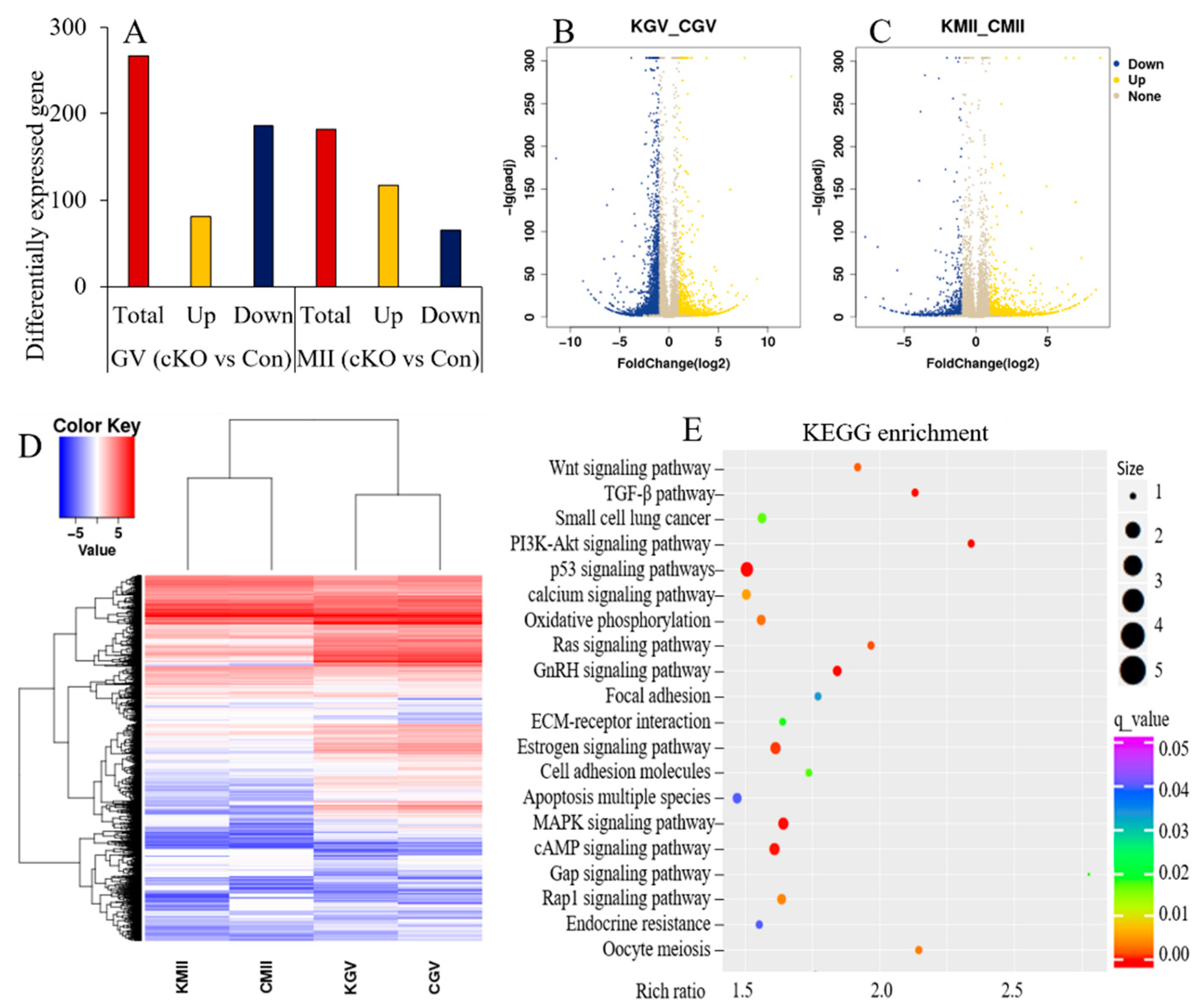
2.7. Loss of Kdm2a Changed the mRNA Expression Pattern in Oocytes
3. Discussion
4. Materials and Methods
4.1. Generation of Kdm2a-Deficient Mice
4.2. Assessment of Female Reproduction
4.3. Ovarian HE Staining and Immunohistochemistry (IHC)
4.4. Oocyte Collection and In Vitro Culture
4.5. Detection of Hormone Levels and Evaluation of Hormone Sensitivity
4.6. Immunofluorescence Assay
4.7. RNA Library Construction and Sequencing
4.8. RNA Extraction and RT-qPCR
4.9. Western Blot
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, V.P.; Yueh, W.T.; Gerton, J.L.; Duncan, F.E. Oocyte-specific deletion of Hdac8 in mice reveals stage-specific effects on fertility. Reproduction 2019, 157, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Szabó, P.E. Maternal DOT1L is dispensable for mouse development. Sci. Rep. 2020, 10, 20636. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Yano, H.; Ogasawara, S.; Yoshioka, S.; Imamura, H.; Okamoto, K.; Tsuneoka, M. Mild glucose starvation induces KDM2A mediated H3K36me2 demethylation through AMPK to reduce rRNA transcription and cell proliferation. Mol. Cell Biol. 2015, 35, 4170–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Li, Q.; Wong, S.H.; Huang, M.; Klein, B.J.; Shen, J.; Ikenouye, L.; Onishi, M.; Schneidawind, D.; Buechele, C.; et al. ASH1L links histone H3 lysine 36 dimethylation to MLL leukemia. Cancer Discov. 2016, 6, 770–783. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.J.; Xu, W.Q.; Lan, F. Histone lysine demethylases in mammalian embryonic development. Exp. Mol. Med. 2017, 49, e325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambacher, S.; Hahn, M.; Schotta, G. Epigenetic regulation of development by histone lysine methylation. Heredity 2010, 105, 24–37. [Google Scholar] [CrossRef]
- Greer, E.L.; Shi, Y. Histone methylation: A dynamic mark in health, disease and inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Wang, E.; Huang, Y.; Guo, X.; Yu, Y.; Du, Q.; Ding, X.; Sun, Y. Inhibition of lysine-specific demethylase-1 (LSD1/KDM1A) promotes the adipogenic differentiation of hESCs through H3K4 methylation. Stem Cell Rev. 2016, 12, 298–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, G.; Ma, B.; Yuan, W.; Zhang, Z.; Chen, P.; Ding, X.; Feng, L.; Shen, X.; Chen, S.; Li, G.; et al. Histone H2A ubiquitination inhibits the enzymatic activity of H3 lysine 36 methyltransferases. J. Bio. Chem. 2013, 288, 30832–30842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Hsu, F.N.; Xu, W.; Xie, X.J.; Ren, X.; Gao, X.; Ni, J.Q.; Ji, J.Y. A developmental genetic analysis of the lysine demethylase KDM2 mutations in Drosophila melanogaster. Mech. Dev. 2014, 133, 36–53. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.L.; Wei, F.; Du, Y.; Song, B.; Wang, D.; Shen, C.; Lu, X.; Cao, Z.; Yang, Q.; Gao, Y.; et al. ATM-mediated KDM2A phosphorylation is required for the DNA damage repair. Oncogene 2016, 35, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, E.; Lupo, G.; Licursi, V.; Negri, R. The role of histone lysine methylation in the response of mammalian cells to ionizing radiation. Front. Genet. 2021, 12, 639602. [Google Scholar] [CrossRef]
- Ho, C.H.; Takizawa, Y.; Kobayashi, W.; Arimura, Y.; Kimura, H.; Kurumizaka, H. Structural basis of nucleosomal histone H4 lysine 20 methylation by SET8 methyltransferase. Life Sci. Alliance 2021, 4, e202000919. [Google Scholar] [CrossRef]
- He, J.; Shen, L.; Wan, M.; Taranova, O.; Wu, H.; Zhang, Y. Kdm2b maintains murine embryonic stem cell status by recruiting PRC1 complex to CpG islands of developmental genes. Nat. Cell Biol. 2013, 15, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulard, M.; Edwards, J.R.; Bestor, T.H. Abnormal X chromosome inactivation and sex-specific gene dysregulation after ablation of FBXL10. Epigenetics Chromatin 2016, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Boulard, M.; Edwards, J.R.; Bestor, T.H. FBXL10 protects polycomb-bound genes from hypermethylation. Nat. Genet. 2015, 47, 479–485. [Google Scholar] [CrossRef]
- Kawakami, E.; Tokunaga, A.; Ozawa, M.; Sakamoto, R.; Yoshida, N. The histone demethylase Fbxl11/Kdm2a plays an essential role in embryonic development by repressing cell-cycle regulators. Mech. Dev. 2015, 135, 31–42. [Google Scholar] [CrossRef]
- Eguizabal, C.; Herrera, L.; De Oñate, L.; Montserrat, N.; Hajkova, P.; Izpisua Belmonte, J.C. Characterization of the epigenetic changes during human gonadal primordial germ cells reprogramming. Stem Cells 2016, 34, 2418–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.H.; Liang, D.Y.; Wang, Q.; Shen, J.; Liu, Q.H.; Peng, Y.B. Knockdown of KDM2A inhibits proliferation associated with TGF-β expression in HEK293T cell. Mol. Cell Biochem. 2019, 456, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Fukuda, T.; Sakamoto, R.; Honda, H.; Yoshida, N. The histone demethylase FBXL10 regulates the proliferation of spermatogonia and ensures long-term sustainable spermatogenesis in mice. Biol. Reprod. 2016, 94, 92. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Yao, R.; Du, J.; Wang, S.; Fan, Z. Depletion of histone demethylase KDM2A enhanced the adipogenic and chondrogenic differentiation potentials of stem cells from apical papilla. Exp. Cell Res. 2013, 319, 2874–2882. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ma, Y.; Ma, P.; Wang, S.; Fan, Z. Demethylation of epiregulin gene by histone demethylase FBXL11 and BCL6 corepressor inhibits osteo/dentinogenic differentiation. Stem Cells 2013, 31, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Dong, R.; Du, J.; Ma, P.; Wang, S.; Fan, Z. Depletion of histone demethylase KDM2A inhibited cell proliferation of stem cells from apical papilla by de-repression of p15INK4B and p27Kip1. Mol. Cell. Biochem. 2013, 379, 115–122. [Google Scholar] [CrossRef]
- Sha, Q.Q.; Jiang, Y.; Yu, C.; Xiang, Y.; Dai, X.X.; Jiang, J.C.; Ou, X.H.; Fan, H.Y. CFP1-dependent histone H3K4 trimethylation in murine oocytes facilitates ovarian follicle recruitment and ovulation in a cell-nonautonomous manner. Cell Mol. Life Sci. 2020, 77, 2997–3012. [Google Scholar] [CrossRef]
- Bilmez, Y.; Talibova, G.; Ozturk, S. Expression of the histone lysine methyltransferases SETD1B, SETDB1, SETD2, and CFP1 exhibits significant changes in the oocytes and granulosa cells of aged mouse ovaries. Histochem. Cell Biol. 2022, 158, 79–95. [Google Scholar] [CrossRef]
- Ijaz, T.; Wakamiya, M.; Sun, H.; Recinos, A.; Tilton, R.G.; Brasier, A.R. Generation and characterization of a novel transgenic mouse harboring conditional nuclear factor-kappa B/RelA knockout alleles. BMC Dev. Biol. 2016, 16, 32. [Google Scholar] [CrossRef] [Green Version]
- Sandlesh, P.; Juang, T.; Safina, A.; Higgins, M.J.; Gurova, K.V. Uncovering the fine print of the CreERT2-LoxP system while generating a conditional knockout mouse model of Ssrp1 gene. PLoS ONE 2018, 13, e0199785. [Google Scholar] [CrossRef] [Green Version]
- Bouabe, H.; Okkenhaug, K. Gene targeting in mice: A review. Methods Mol. Biol. 2013, 1064, 315–336. [Google Scholar]
- Hou, X.; Zhu, S.; Zhang, H.; Li, C.; Qiu, D.; Ge, J.; Guo, X.; Wang, Q. Mitofusin1 in oocyte is essential for female fertility. Redox Biol. 2019, 21, 101110. [Google Scholar] [CrossRef]
- Cao, Y.; Li, M.; Liu, F.; Ni, X.; Wang, S.; Zhang, H.; Sui, X.; Huo, R. Deletion of maternal UHRF1 severely reduces mouse oocyte quality and causes developmental defects in preimplantation embryos. FASEB J. 2019, 33, 8294–8305. [Google Scholar] [CrossRef]
- Mihalas, B.P.; Western, P.S.; Loveland, K.L.; McLaughlin, E.A.; Holt, J.E. Changing expression and subcellular distribution of karyopherins during murine oogenesis. Reproduction 2015, 150, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Hułas-Stasiak, M.; Jakubowicz-Gil, J.; Dobrowolski, P.; Tomaszewska, E.; Muszyński, S. Maternal β-hydroxy-β-methylbutyrate (HMB) supplementation during pregnancy affects early folliculogenesis in the ovary of newborn piglets. Theriogenology 2019, 128, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Petri, T.; Dankert, D.; Demond, H.; Wennemuth, G.; Horsthemke, B.; Grümmer, R. In vitro postovulatory oocyte aging affects H3K9 trimethylation in two-cell embryos after IVF. Ann. Anat. 2020, 227, 151424. [Google Scholar] [CrossRef] [PubMed]
- Ciccone, D.N.; Su, H.; Hevi, S.; Gay, F.; Lei, H.; Bajko, J.; Xu, G.; Li, E.; Chen, T. KDM1B is a histone H3K4 demethylase required to establish maternal genomic imprints. Nature 2009, 461, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, X.; Zhou, S.; Liao, L.; Jiang, R.; Xu, J. The Histone H3K9 demethylase Kdm3b is required for somatic growth and female reproductive function. Int. J. Biol. Sci. 2015, 11, 494–507. [Google Scholar] [CrossRef]
- Andricovich, J.; Perkail, S.; Kai, Y.; Casasanta, N.; Peng, W.; Tzatsos, A. Loss of KDM6A activates super-enhancers to induce gender-specific squamous-like pancreatic cancer and confers sensitivity to BET inhibitors. Cancer Cell. 2018, 33, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.F.; Shah, N.M. Representing sex in the brain, one module at a time. Neuron 2014, 82, 261–278. [Google Scholar] [CrossRef] [Green Version]
- Eckery, D.C.; Moeller, C.L.; Nett, T.M.; Sawyer, H.R. Localization and quantification of binding sites for follicle-stimulating hormone, luteinizing hormone, growth hormone, and insulin-like growth factor I in sheep ovarian follicles. Biol. Reprod. 1997, 57, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Prochazka, R.; Nemcova, L. Mechanisms of FSH- and amphiregulin-induced MAP Kinase 3/1 activation in pig cumulus-oocyte complexes during maturation in vitro. Int. J. Mol. Sci. 2019, 20, 1179. [Google Scholar] [CrossRef] [Green Version]
- Tsukada, Y.; Fang, J.; Erdjument-Bromage, H.; Warren, M.E.; Borchers, C.H.; Tempst, P.; Zhang, Y. Histone demethylation by a family of JmjC domain-containing proteins. Nature 2006, 439, 811–816. [Google Scholar] [CrossRef]
- Cheng, Z.; Cheung, P.; Kuo, A.J.; Yukl, E.T.; Wilmot, C.M.; Gozani, O.; Patel, D.J. A molecular threading mechanism underlies Jumonji lysine demethylase KDM2A regulation of methylated H3K36. Genes Dev. 2014, 28, 1758–1771. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Walport, L.J.; Hopkinson, R.J.; Madden, S.K.; Chowdhury, R.; Schofield, C.J.; Kawamura, A. Studies on the catalytic domains of multiple JmjC oxygenases using peptide substrates. Epigenetics 2014, 9, 1596–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frescas, D.; Guardavaccaro, D.; Bassermann, F.; Koyama-Nasu, R.; Pagano, M. JHDM1B/FBXL10 is a nucleolar protein that represses transcription of ribosomal RNA genes. Nature 2007, 450, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; He, J.; Zhang, Y. Kdm2b promotes induced pluripotent stem cell generation by facilitating gene activation early in reprogramming. Nat. Cell Biol. 2012, 14, 457–466. [Google Scholar] [CrossRef]
- Maidarti, M.; Anderson, R.A.; Telfer, E.E. Crosstalk between PTEN/PI3K/Akt signalling and DNA damage in the oocyte: Implications for primordial follicle activation, oocyte quality and ageing. Cells 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Lee, B.C. Canine oviductal exosomes improve oocyte development via EGFR/MAPK signaling pathway. Reproduction 2020, 160, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, R.; Liu, C.; Ma, R.; Wang, L.; Chen, B.; Li, L.; Niu, J.; Zhao, D.; Mo, F.; et al. Evaluation of decalcification techniques for rat femurs using HE and immunohistochemical staining. Biomed. Res. Int. 2017, 2017, 9050754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, T.; Peters, H. Proposal for a classification of oocytes and follicles in the mouse ovary. J. Reprod. Fertil. 1968, 17, 555–557. [Google Scholar] [CrossRef] [Green Version]
- Mihalas, B.P.; De Iuliis, G.N.; Redgrove, K.A.; McLaughlin, E.A.; Nixon, B. The lipid peroxidation product 4-hydroxynonenal contributes to oxidative stress-mediated deterioration of the ageing oocyte. Sci. Rep. 2017, 7, 6247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, W.; Li, G.; Xu, C.; Nie, X.; Qin, D.; Wang, Q.; Lu, X.; Liu, J.; Li, L. BCAS2 is involved in alternative splicing and mouse oocyte development. FASEB J. 2022, 36, e22128. [Google Scholar] [CrossRef]
- Hu, S.; Liang, X.; Ren, X.; Shi, Y.; Su, H.; Li, Y.; Du, K.; Wang, J.; Jia, X.; Chen, S.; et al. Integrated analysis of mRNA and miRNA expression profiles in the ovary of Oryctolagus cuniculus in response to gonadotrophic stimulation. Front. Endocrinol. 2019, 10, 744. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, H.; Corachán, A.; Aguilar, A.; Quiñonero, A.; Carbajo-García, M.C.; Alamá, P.; Tejera, A.; Taboas, E.; Muñoz, E.; Pellicer, A.; et al. Single-cell RNA sequencing of oocytes from ovarian endometriosis patients reveals a differential transcriptomic profile associated with lower quality. Hum. Reprod. 2019, 34, 1302–1312. [Google Scholar] [CrossRef] [Green Version]
- Spinozzi, G.; Tini, V.; Adorni, A.; Falini, B.; Martelli, M.P. ARPIR: Automatic RNA-Seq pipelines with interactive report. BMC Bioinformatics 2020, 21 (Suppl. S19), 574. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, L.; Deng, T.; Zou, P.; Wang, Y.; Quan, F.; Zhang, Y. Effects of oocyte vitrification on epigenetic status in early bovine embryos. Theriogenology 2016, 86, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Ma, H.; Min, X.; Su, F.; Xiong, Y.; Li, J. Effects of demethylase KDM4B on the biological characteristics and function of yak cumulus cells in vitro. Theriogenology 2021, 174, 85–93. [Google Scholar] [CrossRef]
- Xu, D.; Jiang, X.; He, H.; Liu, D.; Yang, L.; Chen, H.; Wu, L.; Geng, G.; Li, Q. SIRT2 functions in aging, autophagy, and apoptosis in post-maturation bovine oocytes. Life Sci. 2019, 232, 116639. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, X.; Zhang, X.; Yang, M.; Zhu, Y.; Yu, H.; Fei, X.; Mastuda, F.; Lan, D.; Xiong, Y.; Fu, W.; et al. Oocyte-Specific Knockout of Histone Lysine Demethylase KDM2a Compromises Fertility by Blocking the Development of Follicles and Oocytes. Int. J. Mol. Sci. 2022, 23, 12008. https://doi.org/10.3390/ijms231912008
Xiong X, Zhang X, Yang M, Zhu Y, Yu H, Fei X, Mastuda F, Lan D, Xiong Y, Fu W, et al. Oocyte-Specific Knockout of Histone Lysine Demethylase KDM2a Compromises Fertility by Blocking the Development of Follicles and Oocytes. International Journal of Molecular Sciences. 2022; 23(19):12008. https://doi.org/10.3390/ijms231912008
Chicago/Turabian StyleXiong, Xianrong, Xiaojian Zhang, Manzhen Yang, Yanjin Zhu, Hailing Yu, Xixi Fei, Fuko Mastuda, Daoliang Lan, Yan Xiong, Wei Fu, and et al. 2022. "Oocyte-Specific Knockout of Histone Lysine Demethylase KDM2a Compromises Fertility by Blocking the Development of Follicles and Oocytes" International Journal of Molecular Sciences 23, no. 19: 12008. https://doi.org/10.3390/ijms231912008