



Bioactive Phytochemicals from Salix pseudolasiogyne Twigs: Anti-Adipogenic Effect of 2′-O-Acetylsalicortin in 3T3-L1 Cells
 ,
,  and
and
Abstract
:1. Introduction
2. Results
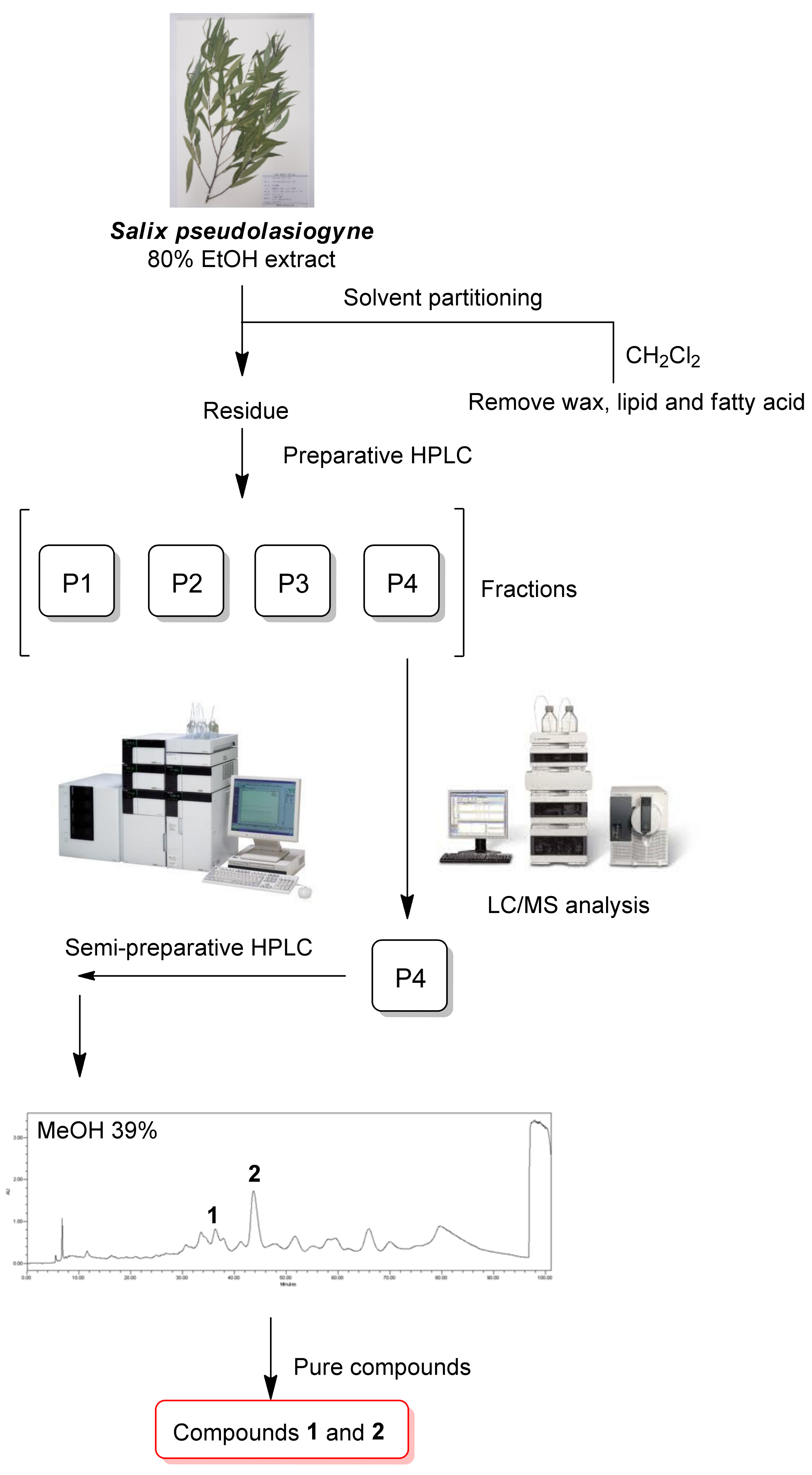
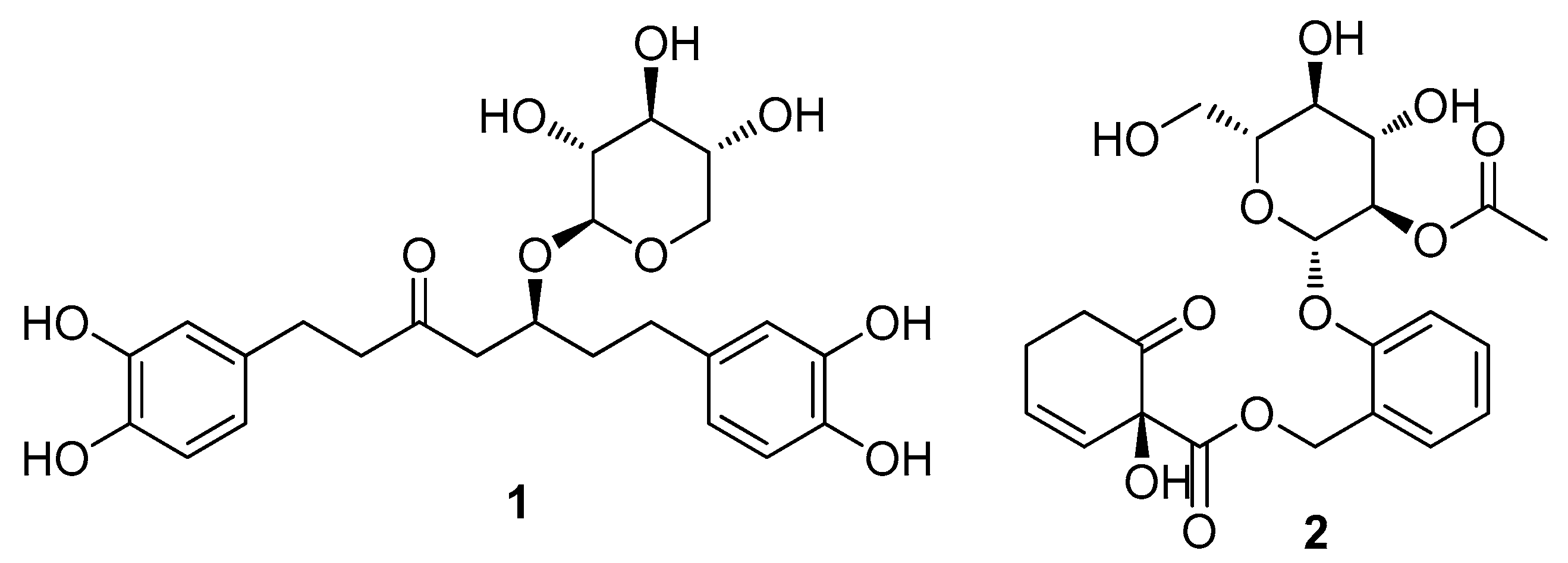
2.1. Isolation and Identification of the Compounds
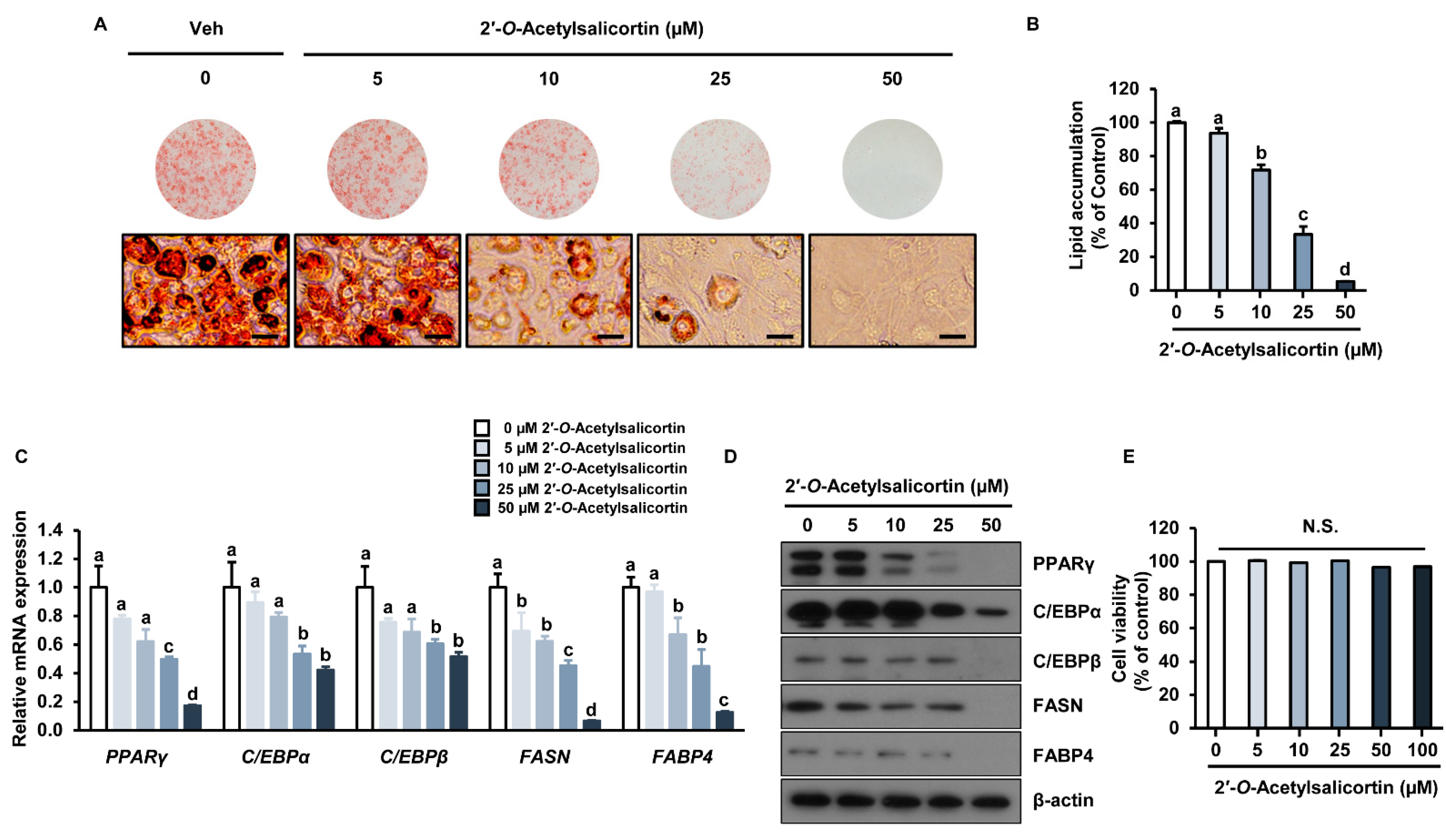
2.2. Effects of the Isolated Compounds on Intracellular Lipid Accumulation
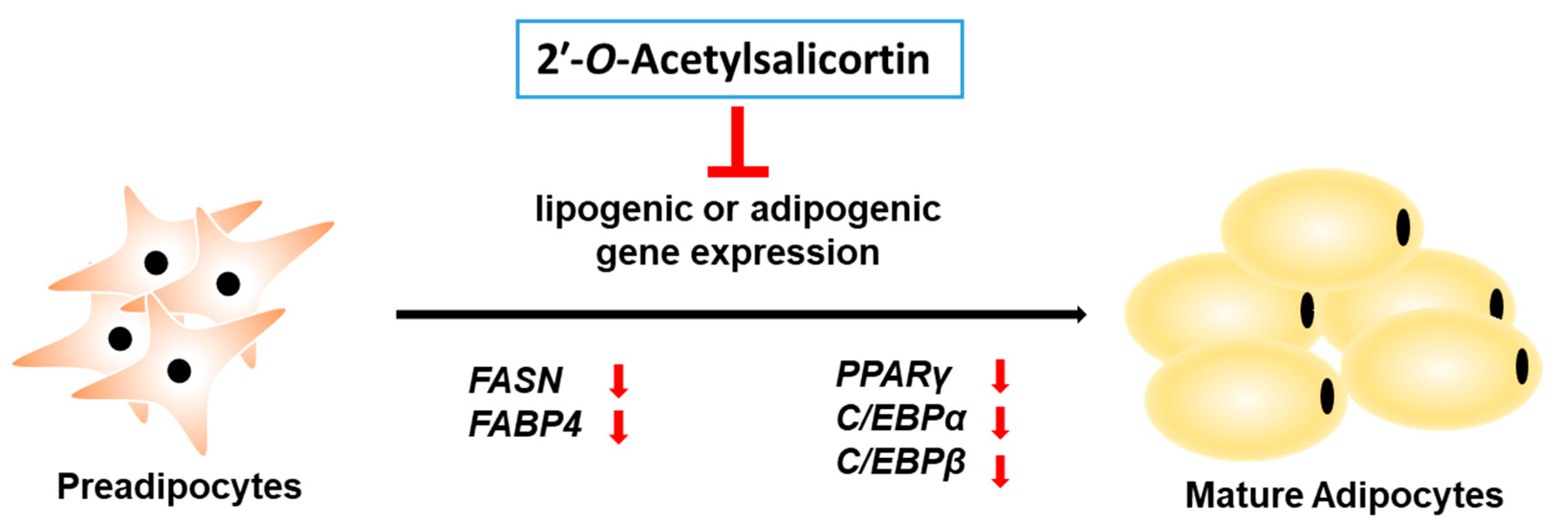
2.3. 2′-O-Acetylsalicortin Attenuates Adipogenesis by Inhibiting the Expression of Adipogenic Transcription Factors and Lipogenic Factors
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.4. 3T3-L1 Cell Culture and Adipocyte Differentiation
4.5. Oil Red O Staining
4.6. Extraction of Protein and Western Blotting
4.7. RNA Isolation and Quantitative Polymerase Chain Reaction (qPCR)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, J.G.; Mahdi, A.J.; Bowen, I.D. The historical analysis of aspirin discovery, its relation to the willow tree and antiproliferative and anticancer potential. Cell Prolif. 2006, 39, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Jerz, G.; Shen, L.; Xiu, L.; Winterhalter, P. Isolation and structure determination of a lignan from the bark of Salix alba. Nat. Prod. Res. 2007, 21, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Freischmidt, A.; Jürgenliemk, G.; Kraus, B.; Okpanyi, S.; Müller, J.; Kelber, O.; Weiser, D.; Heilmann, J. Contribution of flavonoids and catechol to the reduction of ICAM-1 expression in endothelial cells by a standardised Willow bark extract. Phytomed. Int. J. Phytother. Phytopharm. 2012, 19, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Saleem, M. Salix caprea inhibits skin carcinogenesis in murine skin: Inhibition of oxidative stress, ornithine decarboxylase activity and DNA synthesis. J. Ethnopharmacol. 2004, 91, 267–276. [Google Scholar] [CrossRef]
- Li, X.; Liu, Z.; Zhang, X.-F.; Wang, L.-J.; Zheng, Y.-N.; Yuan, C.-C.; Sun, G.-Z. Isolation and Characterization of Phenolic Compounds from the Leaves of Salix matsudana. Molecules 2008, 13, 1530–1537. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S.; Kaur, G.; Jabbar, Z.; Javed, K.; Athar, M. Evaluation of antioxidant activity of Salix caprea flowers. Phytother. Res. 2006, 20, 479–483. [Google Scholar] [CrossRef]
- Han, L.-K.; Sumiyoshi, M.; Zhang, J.; Liu, M.-X.; Zhang, X.-F.; Zheng, Y.-N.; Okuda, H.; Kimura, Y. Anti-obesity action of Salix matsudana leaves (Part 1). Anti-obesity action by polyphenols of Salix matsudana in high fat-diet treated rodent animals. Phytother. Res. 2003, 17, 1188–1194. [Google Scholar] [CrossRef]
- Yang, H.; Lee, S.H.; Sung, S.H.; Kim, J.; Kim, Y.C. Neuroprotective Compounds from Salix pseudo-lasiogyne Twigs and Their Anti-Amnesic Effects on Scopolamine-Induced Memory Deficit in Mice. Planta Med. 2013, 79, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Kyeongshik, K.; Euishik, J. Ferns, Fern-Allies and Seed-Bearing Plants of Korea; Iljinsa: Seoul, Korea, 2003. [Google Scholar]
- Kinlen, D.; Cody, D.; O’Shea, D. Complications of obesity. QJM Int. J. Med. 2018, 111, 437–443. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Green, H.; Meuth, M. An established pre-adipose cell line and its differentiation in culture. Cell 1974, 3, 127–133. [Google Scholar] [CrossRef]
- Birsoy, K.; Berry, R.; Wang, T.; Ceyhan, O.; Tavazoie, S.; Friedman, J.M.; Rodeheffer, M.S. Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis. Development 2011, 138, 4709–4719. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-adipogenic Role of AMP-Activated Protein Kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Kang, H.; Yoo, M.J.; Yu, J.S.; Lee, S.; Yi, S.A.; Beemelmanns, C.; Lee, J.; Kim, K.H. Anti-adipogenic Pregnane Steroid from a Hydractinia-associated Fungus, Cladosporium sphaerospermum SW67. Nat. Prod. Sci. 2020, 26, 230–235. [Google Scholar]
- Lee, S.; Ryoo, R.; Choi, J.H.; Kim, J.-H.; Kim, S.-H.; Kim, K.H. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity. Arch. Pharmacal Res. 2020, 43, 214–223. [Google Scholar] [CrossRef]
- Ha, J.W.; Kim, J.; Kim, H.; Jang, W.; Kim, K.H. Mushrooms: An Important Source of Natural Bioactive Compounds. Nat. Prod. Sci. 2020, 26, 118–131. [Google Scholar]
- Yu, J.S.; Park, M.; Pang, C.; Rashan, L.; Jung, W.H.; Kim, K.H. Antifungal Phenols from Woodfordia uniflora Collected in Oman. J. Nat. Prod. 2020, 83, 2261–2268. [Google Scholar] [CrossRef]
- Lee, K.H.; Kim, J.K.; Yu, J.S.; Jeong, S.Y.; Choi, J.H.; Kim, J.-C.; Ko, Y.-J.; Kim, S.-H.; Kim, K.H. Ginkwanghols A and B, osteogenic coumaric acid-aliphatic alcohol hybrids from the leaves of Ginkgo biloba. Arch. Pharmacal Res. 2021, 44, 514–524. [Google Scholar] [CrossRef]
- Lee, S.; Kim, C.S.; Yu, J.S.; Kang, H.; Yoo, M.J.; Youn, U.J.; Ryoo, R.; Bae, H.Y.; Kim, K.H. Ergopyrone, a Styrylpyrone-Fused Steroid with a Hexacyclic 6/5/6/6/6/5 Skeleton from a Mushroom Gymnopilus orientispectabilis. Org. Lett. 2021, 23, 3315–3319. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Yang, J.; Liu, B.; Huang, F.; Li, Y. Exosomes derived from mangiferin-stimulated perivascular adipose tissue ameliorate endothelial dysfunction. Mol. Med. Rep. 2019, 19, 4797–4805. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Keen, J.A.; Fordham, T.; Morgan, R.A. Adipose tissue dysfunction in obese horses with equine metabolic syndrome. Equine Vet. J. 2019, 51, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Lee, S.H.; Kang, J.; Yang, H.; Jeong, E.J.; Kim, H.P.; Kim, Y.C.; Sung, S.H. Salicortin-Derivatives from Salix pseudo-lasiogyne Twigs Inhibit Adipogenesis in 3T3-L1 Cells via Modulation of C/EBPα and SREBP1c Dependent Pathway. Molecules 2013, 18, 10484–10496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef]
- Shao, D.; Lazar, M.A. Peroxisome Proliferator Activated Receptor γ, CCAAT/ Enhancer-binding Protein α, and Cell Cycle Status Regulate the Commitment to Adipocyte Differentiation. J. Biol. Chem. 1997, 272, 21473–21478. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.Y. Stimulatory Effects of Extracts Prepared from Salix babylonica L. on Fat Catabolism in Mice Fed High-Fat Diet. Adv. Mater. Res. 2012, 518–523, 498–501. [Google Scholar] [CrossRef]
- Roh, C.; Jung, U. Screening of Crude Plant Extracts with Anti-Obesity Activity. Int. J. Mol. Sci. 2012, 13, 1710–1719. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lee, J.; Hernandez, M.A.S.; Mazitschek, R.; Ozcan, U. Treatment of Obesity with Celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Liu, J.; Feng, X.; Hernández, M.A.S.; Mucka, P.; Ibi, D.; Choi, J.W.; Ozcan, U. Withaferin A is a leptin sensitizer with strong antidiabetic properties in mice. Nat. Med. 2016, 22, 1023–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.; Park, S.H.; Park, J.H.; Hwang, J.-T. Anti-obesity effects of medicinal plants from Asian countries and related molecular mechanisms: A review. Rev. Cardiovasc. Med. 2021, 22, 1279–1293. [Google Scholar] [CrossRef]
- Shah, M.A.; Reanmongkol, W.; Radenahmad, N.; Khalil, R.; Ul-Haq, Z.; Panichayupakaranant, P. Anti-hyperglycemic and anti-hyperlipidemic effects of rhinacanthins-rich extract from Rhinacanthus nasutus leaves in nicotinamide-streptozotocin induced diabetic rats. Biomed. Pharmacother. 2019, 113, 108702. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.; Jakkawanpitak, C.; Sermwittayawong, D.; Panichayupakaranant, P. Rhinacanthins-rich Extract Enhances Glucose Uptake and Inhibits Adipogenesis in 3T3-L1 Adipocytes and L6 Myotubes. Pharmacogn. Mag. 2018, 13, S817–S821. [Google Scholar] [CrossRef] [PubMed]
- Panichayupakaranant, P.; Shah, M.A.; Muhammad, H.; Mehmood, Y.; Khalil, R.; Ul-Haq, Z. Superoxide scavenging and antiglycation activity of rhinacanthins-rich extract obtained from the leaves of Rhinacanthus nasutus. Pharmacogn. Mag. 2017, 13, 652–658. [Google Scholar] [CrossRef]
- Kim, C.S.; Subedi, L.; Park, K.J.; Kim, S.Y.; Choi, S.U.; Kim, K.H.; Lee, K.R. Salicin derivatives from Salix glandulosa and their biological activities. Fitoterapia 2015, 106, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Tawfeek, N.; Mahmoud, M.F.; I Hamdan, D.; Sobeh, M.; Farrag, N.; Wink, M.; El-Shazly, A.M. Phytochemistry, Pharmacology and Medicinal Uses of Plants of the Genus Salix: An Updated Review. Front. Pharmacol. 2021, 12, 593856. [Google Scholar] [CrossRef]
- Kim, H.J.; Im, D.U.; Chau, G.C.; Mishra, N.K.; Kim, I.S.; Um, S.H. Novel anti-adipogenic effect of CF3-allylated indole in 3T3-L1 cells. Chem.-Biol. Interact. 2022, 352, 109782. [Google Scholar] [CrossRef]
- Im, D.U.; Kim, S.C.; Chau, G.C.; Um, S.H. Carbamazepine Enhances Adipogenesis by Inhibiting Wnt/β-Catenin Expression. Cells 2019, 8, 1460. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| C/EBPα | ACAACATCGCGGTGCGCAAGA | TGCCATGGCCTTGACCAAGGAG |
| C/EBPβ | GTCCAAACCAACCGCACAT | CAGAGGGAGAAGCAGAGAGTT |
| PPARγ | GGGTGAAACTCTGGGAGATTCTCC | CAGCAACCATTGGGTCAGCTCT |
| FABP4 | TGG AAG CTT GTC TCC AGT GA | AAT CCC CAT TTA CGC TGA TG |
| FASN | CGGAAACTGCAGGAGCTGTC | CACGGAGTTGAGCCGCAT |
| GAPDH | GTCTTCCTGGGCAAGCAGTA | CTGGACAGAAACCCCACTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Jang, Y.S.; Ha, J.W.; Ra, M.-J.; Jung, S.-M.; Yu, J.-N.; Kim, K.; Kim, K.H.; Um, S.H. Bioactive Phytochemicals from Salix pseudolasiogyne Twigs: Anti-Adipogenic Effect of 2′-O-Acetylsalicortin in 3T3-L1 Cells. Int. J. Mol. Sci. 2022, 23, 12006. https://doi.org/10.3390/ijms231912006
Kim HJ, Jang YS, Ha JW, Ra M-J, Jung S-M, Yu J-N, Kim K, Kim KH, Um SH. Bioactive Phytochemicals from Salix pseudolasiogyne Twigs: Anti-Adipogenic Effect of 2′-O-Acetylsalicortin in 3T3-L1 Cells. International Journal of Molecular Sciences. 2022; 23(19):12006. https://doi.org/10.3390/ijms231912006
Chicago/Turabian StyleKim, Hee Jung, Yoon Seo Jang, Ji Won Ha, Moon-Jin Ra, Sang-Mi Jung, Jeong-Nam Yu, Kyunga Kim, Ki Hyun Kim, and Sung Hee Um. 2022. "Bioactive Phytochemicals from Salix pseudolasiogyne Twigs: Anti-Adipogenic Effect of 2′-O-Acetylsalicortin in 3T3-L1 Cells" International Journal of Molecular Sciences 23, no. 19: 12006. https://doi.org/10.3390/ijms231912006