
Super-Resolution Imaging Reveals Dynamic Reticular Cytoophidia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
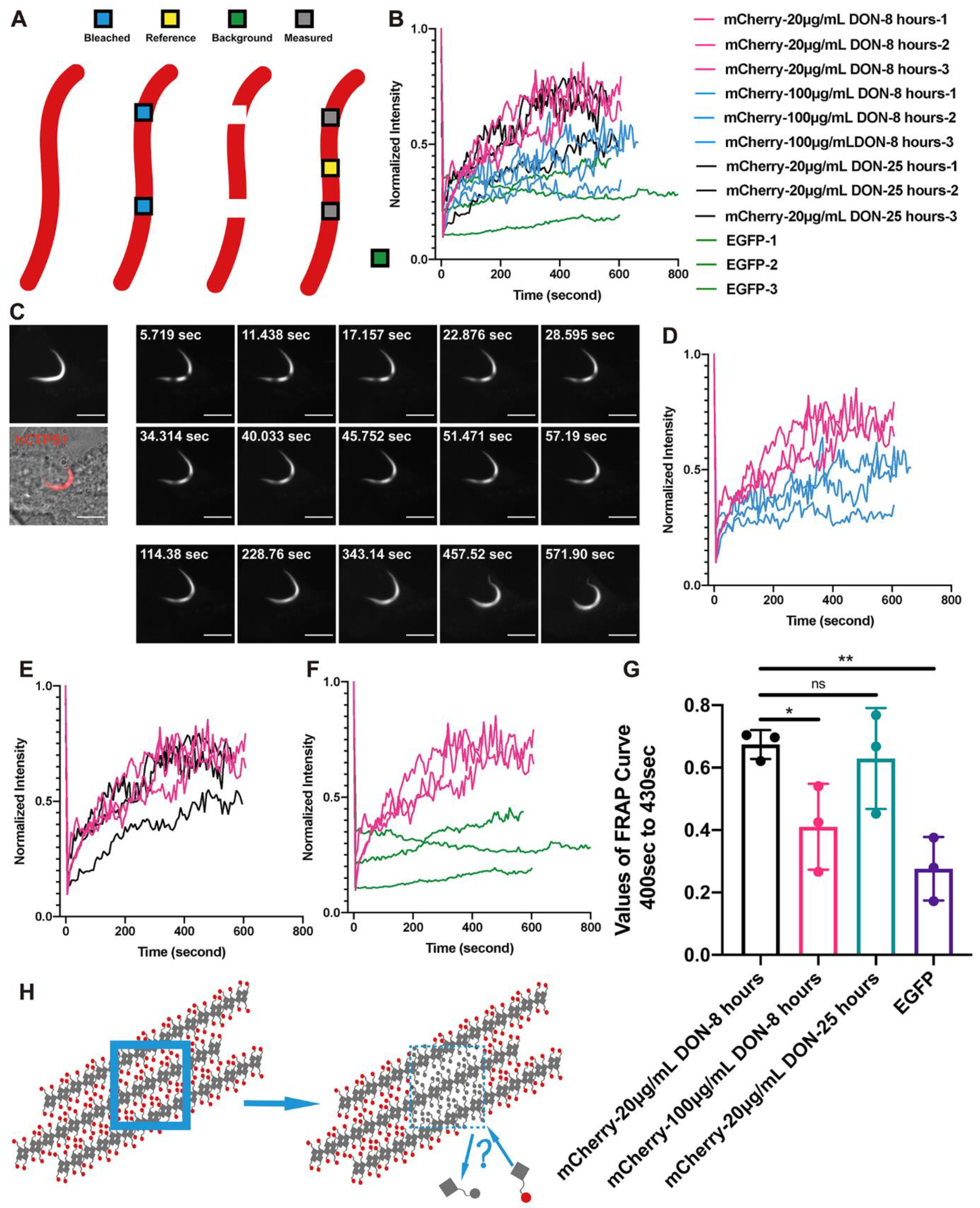
2. Results
2.1. Assembly of CTPS Filaments into Cytoophidia
2.2. Dynamic Equilibria of Cytoophidia
2.3. The Reticular Structure of the hCTPS1 Cytoophidium and Its Localization with hIMPDH2
2.4. CTPS Granules with Tentacles
3. Discussion
3.1. Cytoophidia Are Not Condensates
3.2. Cytoophidia Are Dynamic
3.3. Cytoophidia Are Reticular
3.4. CTPS Can Form Granules with Tentacles
4. Materials and Methods
4.1. Cell Culture
4.2. Constructs and Transfection
4.3. Immunoblotting
4.4. Immunofluorescence
4.5. Microscopy
4.6. Live Imaging
4.7. Image Analysis
4.8. Fluorescence-Activated Cell Sorting (FACS) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.-L. Intracellular compartmentation of CTP synthase in Drosophila. J. Genet. Genom. 2010, 37, 281–296. [Google Scholar] [CrossRef]
- Ingerson-Mahar, M.; Briegel, A.; Werner, J.N.; Jensen, G.J.; Gitai, Z. The metabolic enzyme CTP synthase forms cytoskeletal filaments. Nat. Cell Biol. 2010, 12, 739–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noree, C.; Sato, B.K.; Broyer, R.M.; Wilhelm, J.E. Identification of novel filament-forming proteins in Sac-charomyces cerevisiae and Drosophila melanogaster. J. Cell Biol. 2010, 190, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-L. The enigmatic cytoophidium: Compartmentation of CTP synthase via filament formation. BioEssays 2011, 33, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Tastan, O.Y.; Deussen, Z.A.; Siswick, M.Y.; Liu, J.L. Glutamine analogs promote cy-toophidium assembly in human and Drosophila cells. J. Genet. Genom. 2011, 38, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Carcamo, W.C.; Satoh, M.; Kasahara, H.; Terada, N.; Hamazaki, T.; Chan, J.Y.; Yao, B.; Tamayo, S.; Covini, G.; von Muhlen, C.A.; et al. Induction of cytoplasmic rods and rings structures by inhibition of the CTP and GTP synthetic pathway in mammalian cells. PLoS One 2011, 6, e29690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hulme, L.; Liu, J.-L. Asymmetric inheritance of cytoophidia in Schizosaccharomyces pombe. Biol. Open 2014, 3, 1092–1097. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.J.; Kassim, H.; Huang, Y.; Li, H.; Zhang, J.; Li, G.; Wang, P.Y.; Yan, J.; Ye, F.; Liu, J.L. Filamentation of Metabolic Enzymes in Saccharomyces cerevisiae. J. Genet. Genom. 2016, 43, 393–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.-L. The Cytoophidium and Its Kind: Filamentation and Compartmentation of Metabolic Enzymes. Annu. Rev. Cell Dev. Biol. 2016, 32, 349–372. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-C.; Jeng, Y.-M.; Peng, M.; Keppeke, G.D.; Sung, L.-Y.; Liu, J.-L. CTP synthase forms the cytoophidium in human hepatocellular carcinoma. Exp. Cell Res. 2017, 361, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ye, F.; Ren, J.Y.; Wang, P.Y.; Du, L.L.; Liu, J.L. Active transport of cytoophidia in Schizosaccharomyces pombe. FASEB J. 2018, 32, 5891–5898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daumann, M.; Hickl, D.; Zimmer, D.; DeTar, R.A.; Kunz, H.H.; Möhlmann, T. Characterization of filament-forming CTP synthases from Arabidopsis thaliana. Plant J. 2018, 96, 316–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Ding, K.; Shen, Q.J.; Zhao, S.; Liu, J.L. Filamentation of asparagine synthetase in Saccharomyces cerevisiae. PLoS Genet. 2018, 14, e1007737. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Xiang, H.; Liu, J.-L. CTP synthase forms cytoophidia in archaea. J. Genet. Genom. 2020, 47, 213–223. [Google Scholar] [CrossRef]
- Chang, C.C.; Keppeke, G.D.; Sung, L.Y.; Liu, J.L. CTPS forms the cytoophidium in zebrafish. Exp. Cell Res. 2021, 405, 112684. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Liu, J.-L. The atlas of cytoophidia in Drosophila larvae. J. Genet. Genom. 2020, 47, 321–331. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Zhao, P.A.; Tastan, O.Y.; Liu, J.L. Polarised maintenance of cytoophidia in Drosophila follicle epithelia. Exp. Cell Res. 2021, 402, 112564. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, J.L. Hypoosmolality impedes cytoophidium integrity during nitrogen starvation. Yeast 2021, 38, 276–289. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Zhou, Y.; Wang, Q.Q.; Ding, K.; Zhao, S.; Lu, P.; Liu, J.L. Cytoophidia coupling adipose architecture and metabolism. Cell Mol. Life Sci. 2022, 79, 534. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, C.-J.; Chang, C.-C.; Zhong, J.; Hu, H.-H.; Lu, G.-M.; Liu, J.-L. Structural basis for ligand binding modes of CTP synthase. Proc. Natl. Acad. Sci. USA 2021, 118, e2026621118. [Google Scholar] [CrossRef]
- Calise, S.J.; Carcamo, W.C.; Krueger, C.; Yin, J.D.; Purich, D.L.; Chan, E.K.L. Glutamine deprivation initiates reversible assembly of mammalian rods and rings. Cell. Mol. Life Sci. 2014, 71, 2963–2973. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Gu, J.; Makhov, A.M.; Griffith, J.D.; Mitchell, B.S. Regulation of the Interaction of Inosine Monophosphate Dehydrogenase with Mycophenolic Acid by GTP. J. Biol. Chem. 2006, 281, 206–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Keppeke, G.D.; Sung, L.Y.; Liu, J.L. Interfilament interaction between IMPDH and CTPS cy-toophidia. FEBS J. 2018, 285, 3753–3768. [Google Scholar] [CrossRef] [PubMed]
- Barry, R.M.; Bitbol, A.-F.; Lorestani, A.; Charles, E.J.; Habrian, C.H.; Hansen, J.M.; Li, H.-J.; Baldwin, E.P.; Wingreen, N.S.; Kollman, J.M.; et al. Large-scale filament formation inhibits the activity of CTP synthetase. eLife 2014, 3, e03638. [Google Scholar] [CrossRef] [PubMed]
- Lynch, E.; Hicks, D.R.; Shepherd, M.; Endrizzi, J.A.; Maker, A.; Hansen, J.M.; Barry, R.M.; Gitai, Z.; Baldwin, E.P.; Kollman, J.M. Human CTP synthase filament structure reveals the active enzyme conformation. Nat. Struct. Mol. Biol. 2017, 24, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Liu, J.-L. Forming cytoophidia prolongs the half-life of CTP synthase. Cell Discov. 2019, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Aughey, G.N.; Grice, S.J.; Shen, Q.J.; Xu, Y.; Chang, C.C.; Azzam, G.; Wang, P.Y.; Freeman-Mills, L.; Pai, L.M.; Sung, L.Y.; et al. Nucleotide synthesis is regulated by cytoophidium formation during neurodevelopment and adaptive metabolism. Biol. Open 2014, 3, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strochlic, T.I.; Stavrides, K.P.; Thomas, S.V.; Nicolas, E.; O’Reilly, A.M.; Peterson, J.R. Ack kinase regulates CTP synthase filaments during Drosophila oogenesis. EMBO Rep. 2014, 15, 1184–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Liu, J.-L. mTOR-S6K1 pathway mediates cytoophidium assembly. J. Genet. Genom. 2019, 46, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Guo, C.-J.; Hu, H.-H.; Zhong, J.; Sun, Q.; Liu, D.; Zhou, S.; Chang, C.C.; Liu, J.-L. Drosophila CTP synthase can form distinct substrate- and product-bound filaments. J. Genet. Genom. 2019, 46, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Jans, D.C.; Wurm, C.A.; Riedel, D.; Wenzel, D.; Stagge, F.; Deckers, M.; Rehling, P.; Jakobs, S. STED su-per-resolution microscopy reveals an array of MINOS clusters along human mitochondria. Proc. Natl. Acad. Sci. USA 2013, 110, 8936–8941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezgin, E.; Schneider, F.; Zilles, V.; Urbancic, I.; Garcia, E.; Waithe, D.; Klymchenko, A.S.; Eggeling, C. Polar-ity-Sensitive Probes for Superresolution Stimulated Emission Depletion Microscopy. Biophys. J. 2017, 113, 1321–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaner, N.; A Steinbach, P.; Tsien, R.Y. A guide to choosing fluorescent proteins. Nat. Methods 2005, 2, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Warrington, S.J.; Strutt, H.; Strutt, D. Use of Fluorescence Recovery After Photobleaching (FRAP)Fluorescence Recovery after Photobleaching (FRAP) to Measure In Vivo Dynamics of Cell Junction–Associated Polarity Pro-teins. In Cell Polarity Signaling: Methods and Protocols; Chang, C., Wang, J., Eds.; Springer: New York, NY, USA, 2022; pp. 1–30. [Google Scholar]
- Brangwynne, C.P.; Eckmann, C.R.; Courson, D.S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Julicher, F.; Hyman, A.A. Germline P Granules Are Liquid Droplets That Localize by Controlled Dissolution/Condensation. Science 2009, 324, 1729–1732. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular bio-chemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef]
- Chang, C.-C.; Lin, W.-C.; Pai, L.-M.; Lee, H.-S.; Wu, S.-C.; Ding, S.-T.; Liu, J.-L.; Sung, L.-Y. Cytoophidium assembly reflects upregulation of IMPDH activity. J. Cell Sci. 2015, 128, 3550–3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-C.; Peng, M.; Zhong, J.; Zhang, Z.; Keppeke, G.D.; Sung, L.-Y.; Liu, J.-L. Molecular crowding facilitates bundling of IMPDH polymers and cytoophidium formation. Cell. Mol. Life Sci. 2022, 79, 420. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.-F.; Li, Y.-L.; Li, X.-M.; Liu, J.-L. Super-Resolution Imaging Reveals Dynamic Reticular Cytoophidia. Int. J. Mol. Sci. 2022, 23, 11698. https://doi.org/10.3390/ijms231911698
Fang Y-F, Li Y-L, Li X-M, Liu J-L. Super-Resolution Imaging Reveals Dynamic Reticular Cytoophidia. International Journal of Molecular Sciences. 2022; 23(19):11698. https://doi.org/10.3390/ijms231911698
Chicago/Turabian StyleFang, Yi-Fan, Yi-Lan Li, Xiao-Ming Li, and Ji-Long Liu. 2022. "Super-Resolution Imaging Reveals Dynamic Reticular Cytoophidia" International Journal of Molecular Sciences 23, no. 19: 11698. https://doi.org/10.3390/ijms231911698