
Experimental Research Models to Assess the Cross-Reactivity between Can f 5 and Human PSA—Two Different Perspectives

Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Aim of the Study and Research Group
4.2. An Experimental Model of Inhibition Testing
4.2.1. Baseline Laboratory Procedure
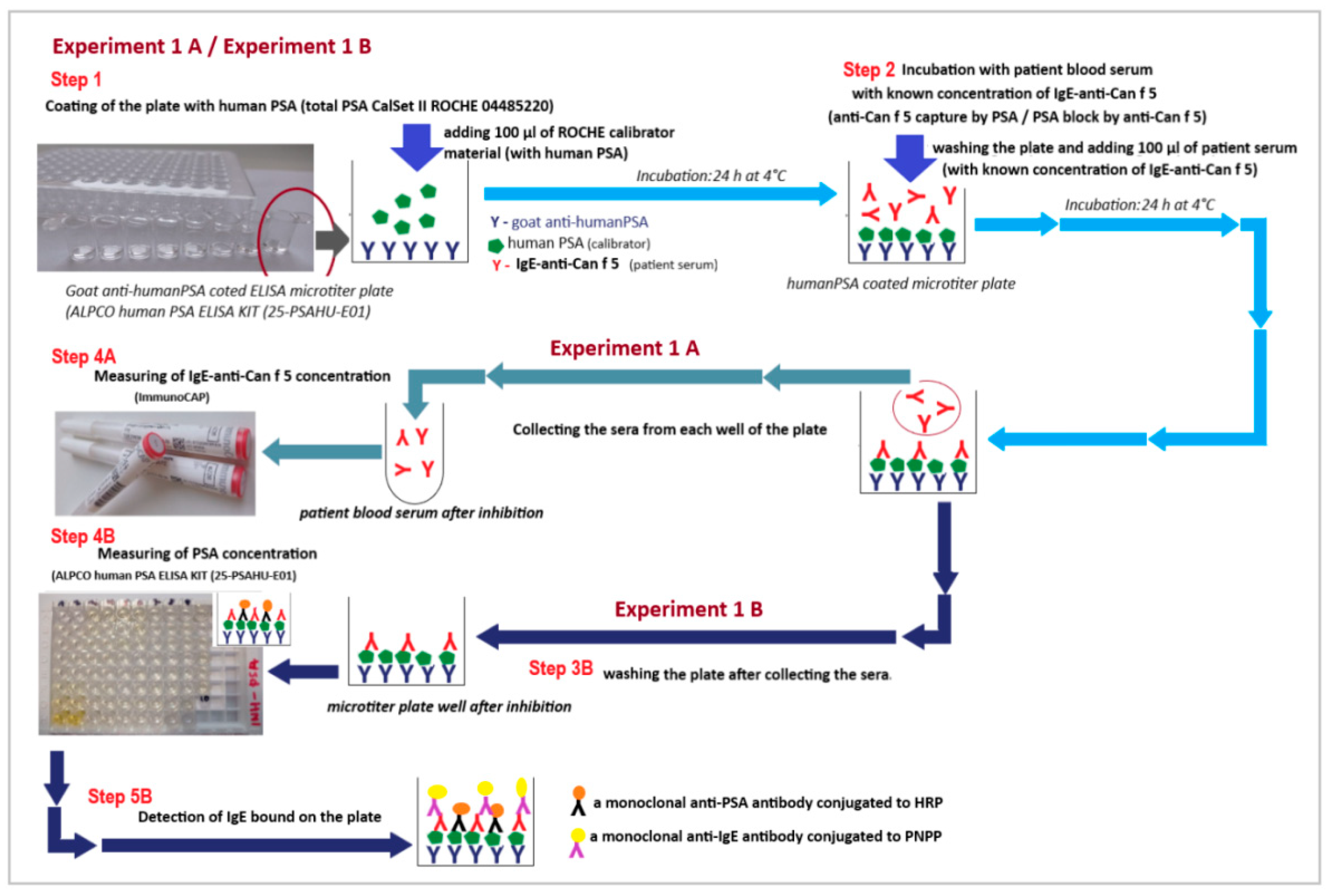
4.2.2. Experiment Model 1 (Experiment 1)—The Solid Phase Inhibition Model (SP-IT)
- Experiment 1 (branch A and branch B)
- Step 1:
- Step 2:
- Step 3:
- Experiment 1A
- Step 4A:
- Experiment 1B
- Step 4B:
- Step 5B:
4.2.3. Experiment Model 2 (Experiment 2)—The Liquid Phase Inhibition Model (LP-IT)
- Experiment 2 (branch A and branch B)
- Step 1—Experiment 2A and Experiment 2B:
- Experiment 2A
- Step 2A:
- Experiment 2B
- Step 2 (Experiment 2B)
- Step 3.1. (Experiment 2B)
- Step 3.2. (Experiment 2B)
- Step 4 (Experiment 2B)
4.3. Controls
4.4. Limitations and Future Perspectives
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI molecular allergology user’s guide. Pediatr. Allergy. Immunol. 2016, 27 (Suppl. S23), 1–250. [Google Scholar] [CrossRef]
- Popescu, F.D. Cross-reactivity between aeroallergens and food allergens. World. J. Methodol. 2015, 5, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Konradsen, J.R.; Borres, M.P.; Nilsson, C. Unusual and Unexpected Allergic Reactions Can Be Unraveled by Molecular Allergy Diagnostics. Int. Arch. Allergy Immunol. 2021, 182, 904–916. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C. Assessment of allergen cross-reactivity. Clin. Mol. Allergy 2007, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Kapingidza, A.B.; Dolamore, C.; Kowal, K. A robust method for the estimation and visualization of IgE cross-reactivity likelihood between allergens belonging to the same protein family. PLoS ONE 2018, 13, e0208276. [Google Scholar] [CrossRef]
- Liccardi, G.; Calzetta, L.; Milanese, M.; Passalacqua, G.; Rogliani, P. Can f 5 as a suitable marker of dog allergy: Assess male dog exposure before banning it. J. Allergy Clin. Immunol. 2019, 143, 1657–1658. [Google Scholar] [CrossRef]
- David, M.K.; Leslie, S.W. Prostate Specific Antigen. In Stat Pearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Laflamme, B.A.; Wolfner, M.F. Identification and function of proteolysis regulators in seminal fluid. Mol. Reprod. Dev. 2013, 80, 80–101. [Google Scholar] [CrossRef]
- Ghosh, D.; Bernstein, J.A. Systemic and localized seminal plasma hypersensitivity patients exhibit divergent immunologic characteristics. J. Allergy Clin. Immunol. 2014, 134, 969–972. [Google Scholar] [CrossRef]
- Basagaña, M.; Bartolomé, B.; Pastor, C.; Torres, F.; Alonso, R.; Vivanco, F.; Cisteró-Bahíma, A. Allergy to human seminal fluid: Cross-reactivity with dog dander. J. Allergy Clin. Immunol. 2008, 121, 233–239. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakagawa, Y.; Kotobuki, Y.; Katayama, I. A case of human seminal plasma allergy sensitized with dog prostatic kallikrein, Can f 5. Allergol. Int. 2019, 68, 259–260. [Google Scholar] [CrossRef]
- Sublett, J.W.; Bernstein, J.A. Seminal plasma hypersensitivity reactions: An updated review. Mt. Sinai J. Med. 2011, 78, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Liccardi, G.; Caminati, M.; Senna, G.; Calzetta, L.; Rogliani, P. Anaphylaxis and intimate behavior. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Mayerhofer, A.; Raemsch, R.; Ring, J.; Köhn, F.M. Prostate-specific antigen as allergen in human seminal plasma allergy. J. Allergy Clin. Immunol. 2006, 117, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Kofler, L.; Kofler, H.; Mattsson, L.; Lidholm, J. A case of dog-related human seminal plasma allergy. Eur. Ann. Allergy Clin. Immunol. 2012, 44, 89–92. [Google Scholar]
- Brazdova, A.; Senechal, H.; Peltre, G.; Poncet, P. Immune Aspects of Female Infertility. Int. J. Fertil. Steril. 2016, 10, 1–10. [Google Scholar] [CrossRef]
- Ukleja-Sokołowska, N.; Lis, K.; Żbikowska-Gotz, M.; Adamczak, R.; Bartuzi, Z. Analysis of allergen profile in patients sensitized to canine allergen and potential Can f 5 cross-reactivity with human PSA. Int. J. Immunopathol. Pharmacol. 2021, 35, 1–10. [Google Scholar] [CrossRef]
- Mattsson, L.; Lundgren, T.; Everberg, H.; Larsson, H.; Lidholm, J. Prostatic kallikrein: A new major dog allergen. J. Allergy Clin. Immunol. 2009, 123, 362–368. [Google Scholar] [CrossRef]
- Sankian, M.; Varasteh, A.; Pazouki, N.; Mahmoudi, M. Sequence homology: A poor predictive value for profilins cross-reactivity. Clin. Mol. Allergy 2005, 3, 13. [Google Scholar] [CrossRef]
- Schmidt-Hieltjes, Y.; Teodorowicz, M.; Jansen, A.; den Hartog, G.; Elfvering-Berendsen, L.; de Jong, N.W.; Savelkoul, H.F.; Ruinemans-Koerts, J. An alternative inhibition method for determining cross-reactive allergens. Clin. Chem. Lab. Med. 2017, 55, 248–253. [Google Scholar] [CrossRef]
- Jeongmin, L.; Se-Ah, J.; Sooyoung, L. Cross-reactivity of Can f 1 with Syrian hamster and Fel d 1 in children. Allergol. Immunopathol. 2021, 49, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ukleja-Sokołowska, N.; Gawrońska-Ukleja, E.; Żbikowska-Gotz, M.; Bartuzi, Z.; Sokołowski, Ł. Sunflower seed allergy. Int. J. Immunopathol. Pharmacol. 2016, 29, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukleja-Sokołowska, N.; Gawrońska-Ukleja, E.; Lis, K.; Żbikowska-Gotz, M.; Sokołowski, Ł.; Bartuzi, Z. Anaphylactic reaction in patient allergic to mango. Allergy Asthma Clin. Immunol. 2018, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Lipman, N.S.; Jackson, L.R.; Trudel, L.J.; Weis-Garcia, F. Monoclonal versus polyclonal antibodies: Distinguishing characteristics, applications, and information resources. ILAR J. 2005, 46, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, A.; Plückthun, A. Reproducibility: Standardize antibodies used in research. Nature 2015, 518, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.; Tokarz-Deptuła, B.; Deptuła, J.; Deptuła, W. Natural antibodies—Facts known and unknown. Cent. Eur. J. Immunol. 2018, 43, 466–475. [Google Scholar] [CrossRef]
- Fung, K.Y.; Glode, L.M.; Green, S.; Duncan, M.W. A comprehensive characterization of the peptide and protein constituents of human seminal fluid. Prostate 2004, 61, 171–181. [Google Scholar] [CrossRef]
- Anderton, S.M. Post-translational modifications of self antigens: Implications for autoimmunity. Curr. Opin. Immunol. 2004, 16, 753–758. [Google Scholar] [CrossRef]
- Bernardi, M.L.; Giangrieco, I.; Camardella, L.; Ferrara, R.; Palazzo, P.; Panico, M.R.; Crescenzo, R.; Carratore, V.; Zennaro, D.; Liso, M.; et al. Allergenic lipid transfer proteins from plant-derived foods do not immunologically and clinically behave homogeneously: The kiwifruit LTP as a model. PLoS ONE 2011, 6, e27856. [Google Scholar] [CrossRef]
- Yakhlef, M.; Giangrieco, I.; Ciardiello, M.A.; Fiume, I.; Mari, A.; Souiki, L.; Pocsfalvi, G. Potential allergenicity of Medicago sativa investigated by a combined IgE-binding inhibition, proteomics and in silico approach. J. Sci. Food Agric. 2021, 101, 1182–1192. [Google Scholar] [CrossRef]
- Tuppo, L.; Giangrieco, I.; Tamburrini, M.; Alessandri, C.; Mari, A.; Ciardiello, M.A. Detection of Allergenic Proteins in Foodstuffs: Advantages of the Innovative Multiplex Allergen Microarray-Based Immunoassay Compared to Conventional Methods. Foods 2022, 11, 878. [Google Scholar] [CrossRef]
- Inomata, N.; Miyakawa, M.; Aihara, M. Gibberellin-regulated protein in Japanese apricot is an allergen cross-reactive to Pru p 7. Immun. Inflamm. Dis. 2017, 5, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Savi, E.; Incorvaia, C.; Boni, E.; Mauro, M.; Peveri, S.; Pravettoni, V.; Quercia, O.; Reccardini, F.; Montagni, M.; Pessina, L.; et al. Which immunotherapy product is better for patients allergic to Polistes venom? A laboratory and clinical study. PLoS ONE 2017, 12, e0180270. [Google Scholar] [CrossRef] [PubMed]
- Caruso, B.; Bonadonna, P.; Severino, M.G.; Manfredi, M.; Dama, A.; Schiappoli, M.; Rizzotti, P.; Senna, G.; Passalacqua, G. Evaluation of the IgE cross-reactions among vespid venoms. A possible approach for the choice of immunotherapy. Allergy 2007, 62, 561–564. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indiwidual Case | anti-Can f 5 IgE [kUA/L] Baseline (before Inhibition) | anti-Can f 5 IgE [kUA/L] after Inhibition Experiment 1A | anti-Can f 5 IgE [kUA/L] after Inhibition Experiment 2A | [%] Decrease anti-Can f 5 IgE Experiment 1A | [%] Decrease anti-Can f 5 IgE Experiment 2A | PSA [ng/mL] Experiment 1B | PSA [ng/mL] Experiment 2A | PSA [ng/mL] Experiment 2B | [%] Decrease PSA Experiment 1B | [%] Decrease PSA Experiment 2A | [%] Decrease PSA Experiment 2B | IgE [O.D.] Experiment 1B | PSA [O.D.] Experiment 2B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4.92 | 4.38 | 3.11 | 11.0 | 36.8 | 75.80 | 72.80 | 51.95 | 10.6 | 14.1 | 28.6 | 0.036 | 0.122 |
| 2 | 3.83 | 3.43 | 2.34 | 10.4 | 38.9 | 77.50 | 74.50 | 55.88 | 8.6 | 12.1 | 25.0 | 0.055 | 0.119 |
| 3 | 2.38 | 2.05 | 1.42 | 13.9 | 40.3 | 77.98 | 73.29 | 58.69 | 8.0 | 13.5 | 19.9 | 0.051 | 0.115 |
| 4 | 0.54 | 0.40 | 0.43 | 25.9 | 20.4 | 75.64 | 70.01 | 54.28 | 10.8 | 17.4 | 22.5 | 0.050 | 0.085 |
| 5 | 0.50 | 0.40 | 0.38 | 20.0 | 24.0 | 76.60 | 71.12 | 59.06 | 9.7 | 16.1 | 17.0 | 0.059 | 0.085 |
| 6 | 1.48 | 1.26 | 0.97 | 14.9 | 34.5 | 77.66 | 75.66 | 60.97 | 8.4 | 10.7 | 19.4 | 0.166 | 0.111 |
| 7 | 23.40 | 16.83 | 7.70 | 28.1 | 67.1 | 77.18 | 74.98 | 65.44 | 8.9 | 11.5 | 12.7 | 0.055 | 0.151 |
| 8 | 6.76 | 5.93 | 4.12 | 12.3 | 39.1 | 76.92 | 74.22 | 53.23 | 9.2 | 12.4 | 25.6 | 0.112 | 0.131 |
| 9 | 5.22 | 3.68 | 3.07 | 29.5 | 41.2 | 71.45 | 68.58 | 43.66 | 15.7 | 19.1 | 36.3 | 0.114 | 0.129 |
| 10 | 0.69 | 0.60 | 0.55 | 13.0 | 20.3 | 76.65 | 74.21 | 62.26 | 9.6 | 12.4 | 16.1 | 0.069 | 0.099 |
| 11 | 20.40 | 14.50 | 12.10 | 28.9 | 40.7 | 78.72 | 75.36 | 50.04 | 7.1 | 11.1 | 33.6 | 0.121 | 0.149 |
| 12 | 5.13 | 3.44 | 3.34 | 32.9 | 34.9 | 72.51 | 68.78 | 45.79 | 14.4 | 18.8 | 27.6 | 0.110 | 0.128 |
| 13 | 1.09 | 0.81 | 0.77 | 26.5 | 29.4 | 68.10 | 65.12 | 53.86 | 19.6 | 23.2 | 17.3 | 0.039 | 0.099 |
| 14 | 8.85 | 5.79 | 5.31 | 36.6 | 40.0 | 71.82 | 70.59 | 53.86 | 15.3 | 16.7 | 23.7 | 0.120 | 0.132 |
| 15 | 0.48 | 0.39 | 0.39 | 18.8 | 18.8 | 73.09 | 71.22 | 58.49 | 18.8 | 16.0 | 17.9 | 0.056 | 0.082 |
| 16 | 3.00 | 2.43 | 1.96 | 19.0 | 34.7 | 69.11 | 66.31 | 48.02 | 18.5 | 21.8 | 27.3 | 0.059 | 0.117 |
| 17 | 34.40 | 21.42 | 19.10 | 37.7 | 44.5 | 59.97 | 57.19 | 48.34 | 29.2 | 32.5 | 15.5 | 0.123 | 0.172 |
| 18 | 0.92 | 0.82 | 0.70 | 10.9 | 23.9 | 81.17 | 79.01 | 65.34 | 4.2 | 6.8 | 17.3 | 0.071 | 0.100 |
| 19 | 75.70 | 56.88 | 35.50 | 24.9 | 53.1 | 78.94 | 78.36 | 77.34 | 6.9 | 7.5 | 1.3 | 0.223 | 0.289 |
| 20 | 8.69 | 5.65 | 5.23 | 35.0 | 39.8 | 72.77 | 69.11 | 50.46 | 14.1 | 18.5 | 27.0 | 0.110 | 0.145 |
| 21 | 7.30 | 6.64 | 4.39 | 36.4 | 39.9 | 72.46 | 68.54 | 46.75 | 14.5 | 19.1 | 31.8 | 0.099 | 0.142 |
| 22 | 7.06 | 5.18 | 4.99 | 26.7 | 29.3 | 69.16 | 59.36 | 43.24 | 18.4 | 30.0 | 27.2 | 0.089 | 0.144 |
| 23 | 1.78 | 1.59 | 1.17 | 12.0 | 34.3 | 69.27 | 58.21 | 39.95 | 18.3 | 31.3 | 31.4 | 0.069 | 0.112 |
| 24 | 1.09 | 0.93 | 0.82 | 14.7 | 24.8 | 81.38 | 80.36 | 57.90 | 4.0 | 5.2 | 28.0 | 0.059 | 0.111 |
| 25 | 0.59 | 0.45 | 0.48 | 23.7 | 18.6 | 78.94 | 75.27 | 72.34 | 6.9 | 11.2 | 3.9 | 0.039 | 0.089 |
| 26 | 1.07 | 0.75 | 0.71 | 29.9 | 33.6 | 81.22 | 78.90 | 54.39 | 4.2 | 6.9 | 30.7 | 0.045 | 0.110 |
| 27 | 0.99 | 0.87 | 0.78 | 12.1 | 21.2 | 81.33 | 78.63 | 61.41 | 4.0 | 7.2 | 21.9 | 0.036 | 0.100 |
| 28 | 1.15 | 0.97 | 0.75 | 15.7 | 34.8 | 80.42 | 71.33 | 40.80 | 5.1 | 15.8 | 42.8 | 0.039 | 0.115 |
| 29 | 2.82 | 2.36 | 1.84 | 16.4 | 34.8 | 80.05 | 71.96 | 37.82 | 5.6 | 15.1 | 47.5 | 0.045 | 0.136 |
| 30 | 0.93 | 0.79 | 0.73 | 15.1 | 21.5 | 80.16 | 78.94 | 38.25 | 5.4 | 6.9 | 51.6 | 0.055 | 0.116 |
| 31 | 19.70 | 16.38 | 8.87 | 16.9 | 55.0 | 71.61 | 68.47 | 32.93 | 15.2 | 19.2 | 51.9 | 0.112 | 0.143 |
| Median | Min. | Max. | Average | SD | |
|---|---|---|---|---|---|
| anti-Can f 5 IgE baseline (before inhibition) [kUA/L] | 2.82 | 0.48 | 75.70 | 8.16 | 14.87 |
| anti-Can f 5 IgE after inhibition Experiment 1A [kUA/L] | 2.36 | 0.39 | 56.88 | 6.06 | 10.91 |
| anti-Can f 5 IgE after inhibition Experiment 2A [kUA/L] | 1.84 | 0.38 | 35.50 | 4.32 | 7.07 |
| anti-Can f 5 IgE after inhibition Experiment 2B [kUA/L] | -------- | <0.1 | <0.1 | -------- | -------- |
| [%] anti-Can f 5 IgE Experiment 1A [kUA/L] | 19.00 | 10.44 | 37.73 | 21.60 | 8.79 |
| [%] anti-Can f 5 IgE Experiment 2A [kUA/L] | 34.75 | 18.64 | 67.09 | 34.51 | 11.25 |
| PSA baseline concentration [ng/mL] | 84.75 | 84.75 | 84.75 | 84.75 | 0.00 |
| PSA Experiment 1B [ng/mL] | 76.65 | 59.97 | 81.38 | 75.34 | 4.99 |
| PSA Experiment 2A [ng/mL] | 71.96 | 57.19 | 80.36 | 71.63 | 5.97 |
| PSA Experiment 2B [ng/mL] | 53.86 | 32.93 | 77.34 | 52.99 | 10.22 |
| [%] PSA Experiment 1B | 9.56 | 3.98 | 29.24 | 11.25 | 6.02 |
| [%] PSA Experiment 2A | 15.09 | 5.18 | 32.52 | 15.49 | 7.04 |
| [%] PSA Experiment 2B | 25.59 | 1.30 | 51.91 | 25.81 | 11.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lis, K.; Ukleja-Sokołowska, N.; Adamczak, R.; Bartuzi, Z. Experimental Research Models to Assess the Cross-Reactivity between Can f 5 and Human PSA—Two Different Perspectives. Int. J. Mol. Sci. 2022, 23, 11223. https://doi.org/10.3390/ijms231911223
Lis K, Ukleja-Sokołowska N, Adamczak R, Bartuzi Z. Experimental Research Models to Assess the Cross-Reactivity between Can f 5 and Human PSA—Two Different Perspectives. International Journal of Molecular Sciences. 2022; 23(19):11223. https://doi.org/10.3390/ijms231911223
Chicago/Turabian StyleLis, Kinga, Natalia Ukleja-Sokołowska, Rafał Adamczak, and Zbigniew Bartuzi. 2022. "Experimental Research Models to Assess the Cross-Reactivity between Can f 5 and Human PSA—Two Different Perspectives" International Journal of Molecular Sciences 23, no. 19: 11223. https://doi.org/10.3390/ijms231911223