

Genome-Wide Identification, Evolution, and Expression Analysis of GASA Gene Family in Prunus mume

Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of GASA Genes across Six Plant Species
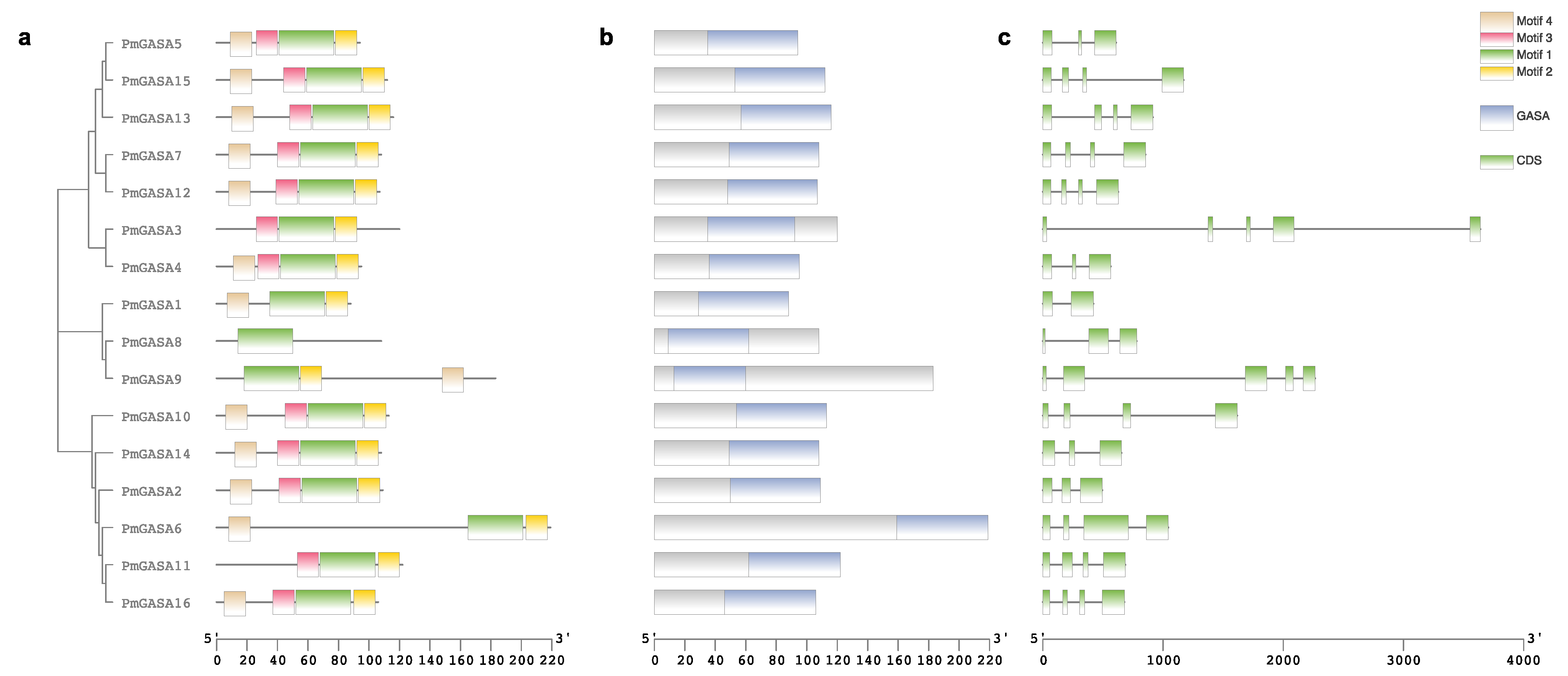
2.2. Gene Structure and Protein Motif Analysis of PmGASAs
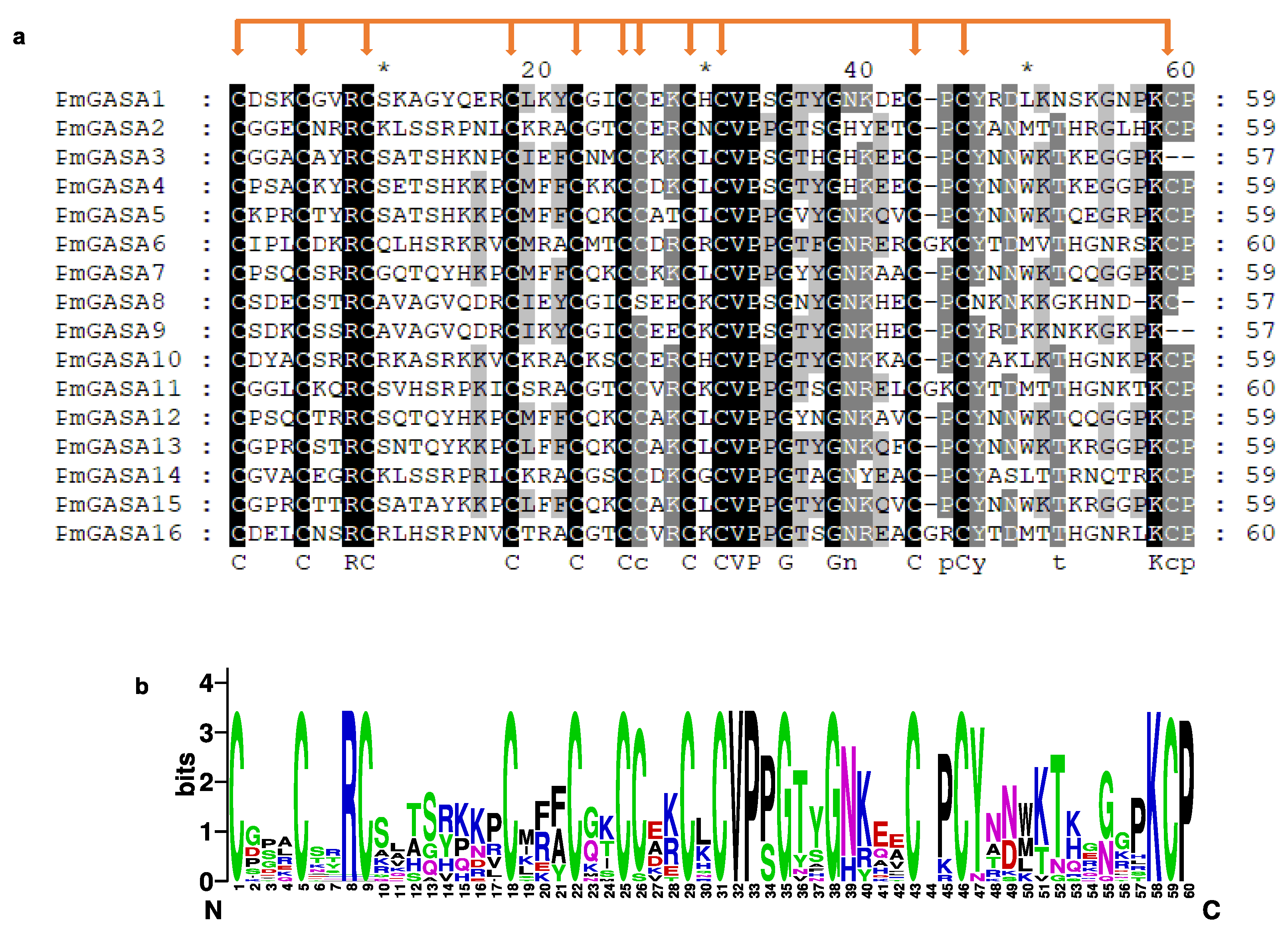
2.3. Physiochemical Analysis of PmGASA Proteins
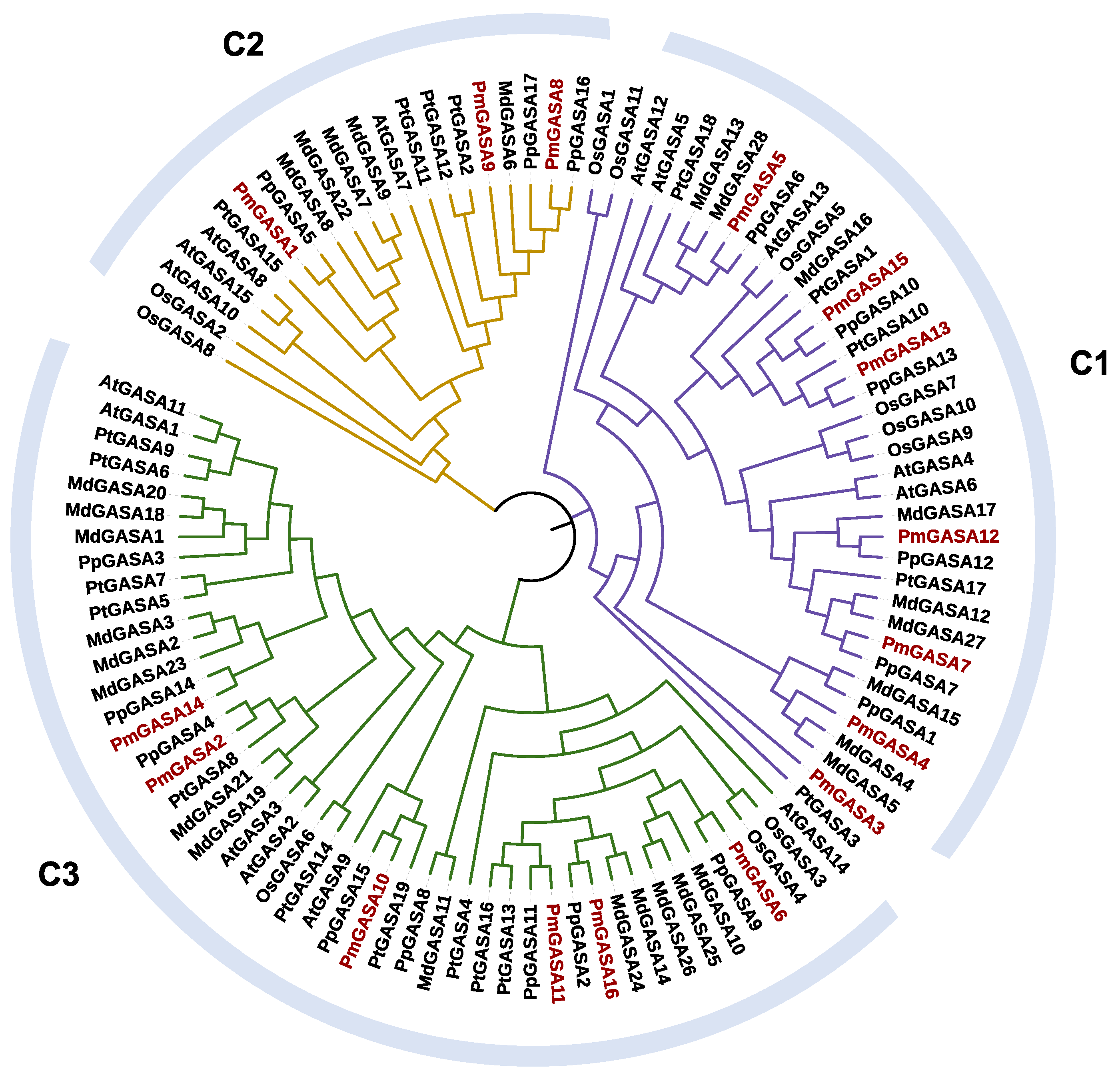
2.4. Phylogenetic Analysis of GASA Family Genes
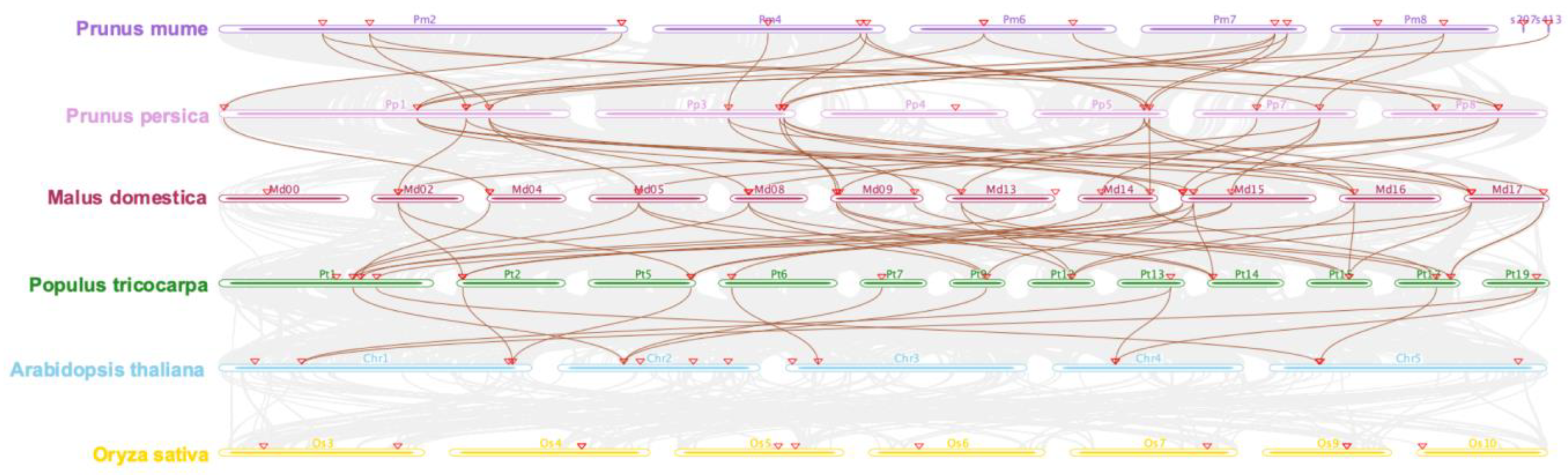
2.5. Evolutionary Analysis of GASA Family Genes in P. mume and Other Species
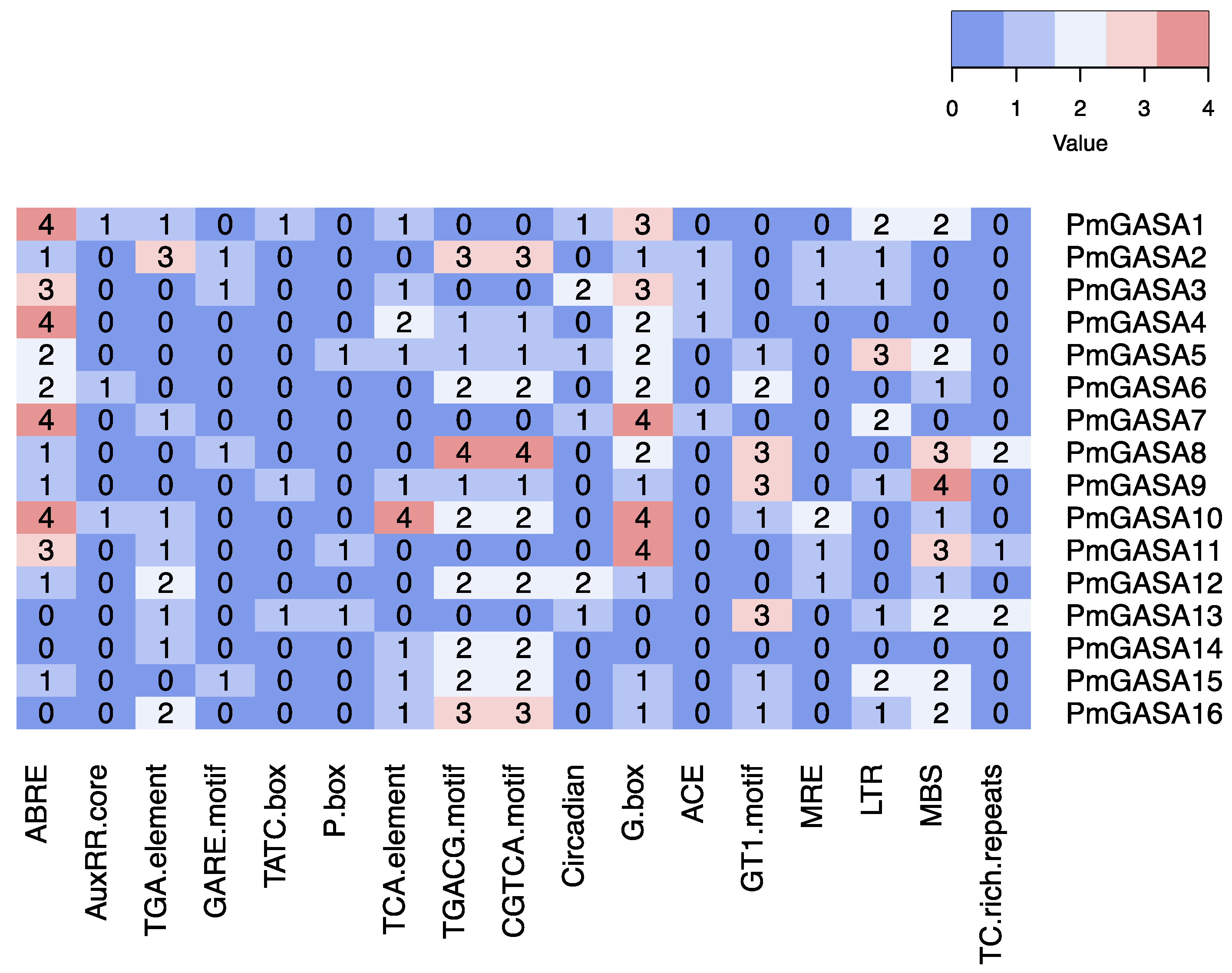
2.6. Promoter Analysis of PmGASA Genes and Their Possible Activators
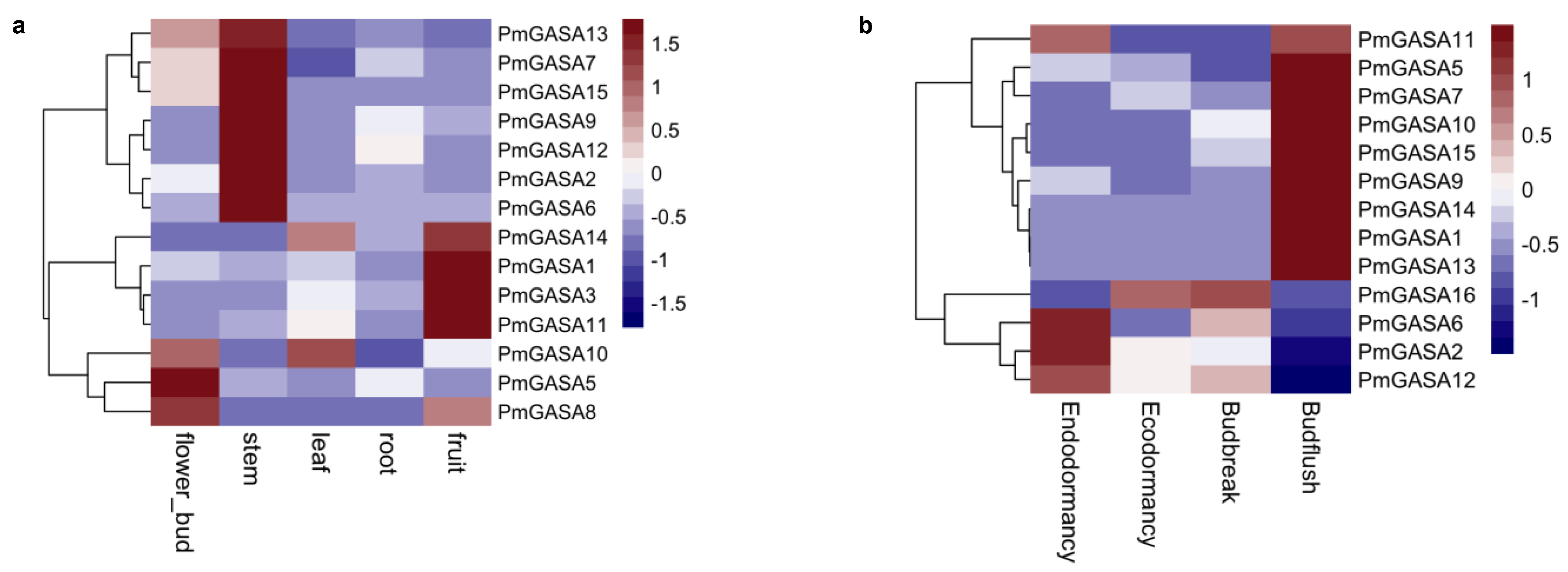
2.7. Tissue-Specific Expression Patterns of PmGASA Genes
2.8. Expression Patterns of PmGASA Genes during Floral Bud Development
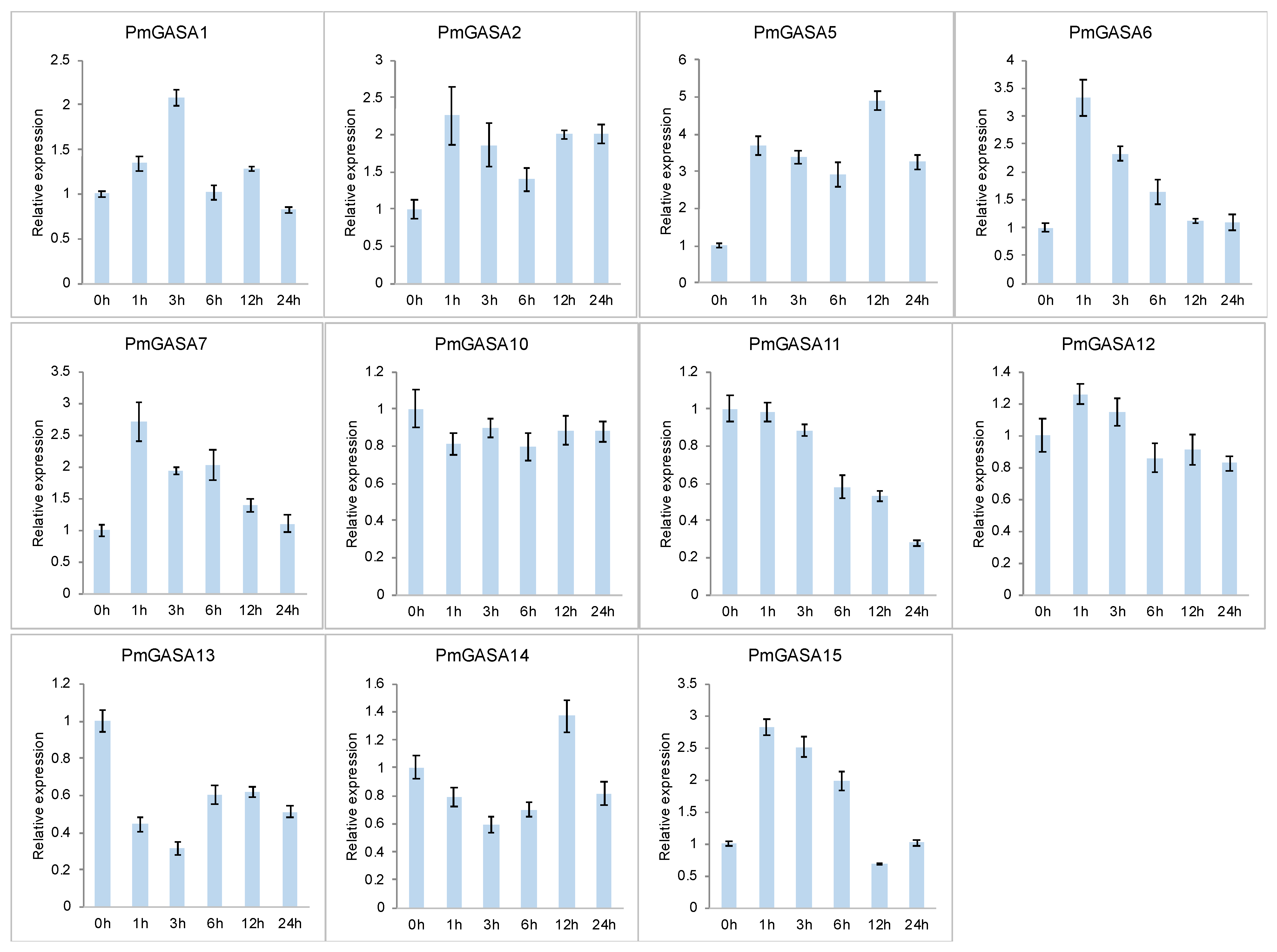
2.9. Expression Patterns of PmGASA Genes in Response to Gibberellin Treatments
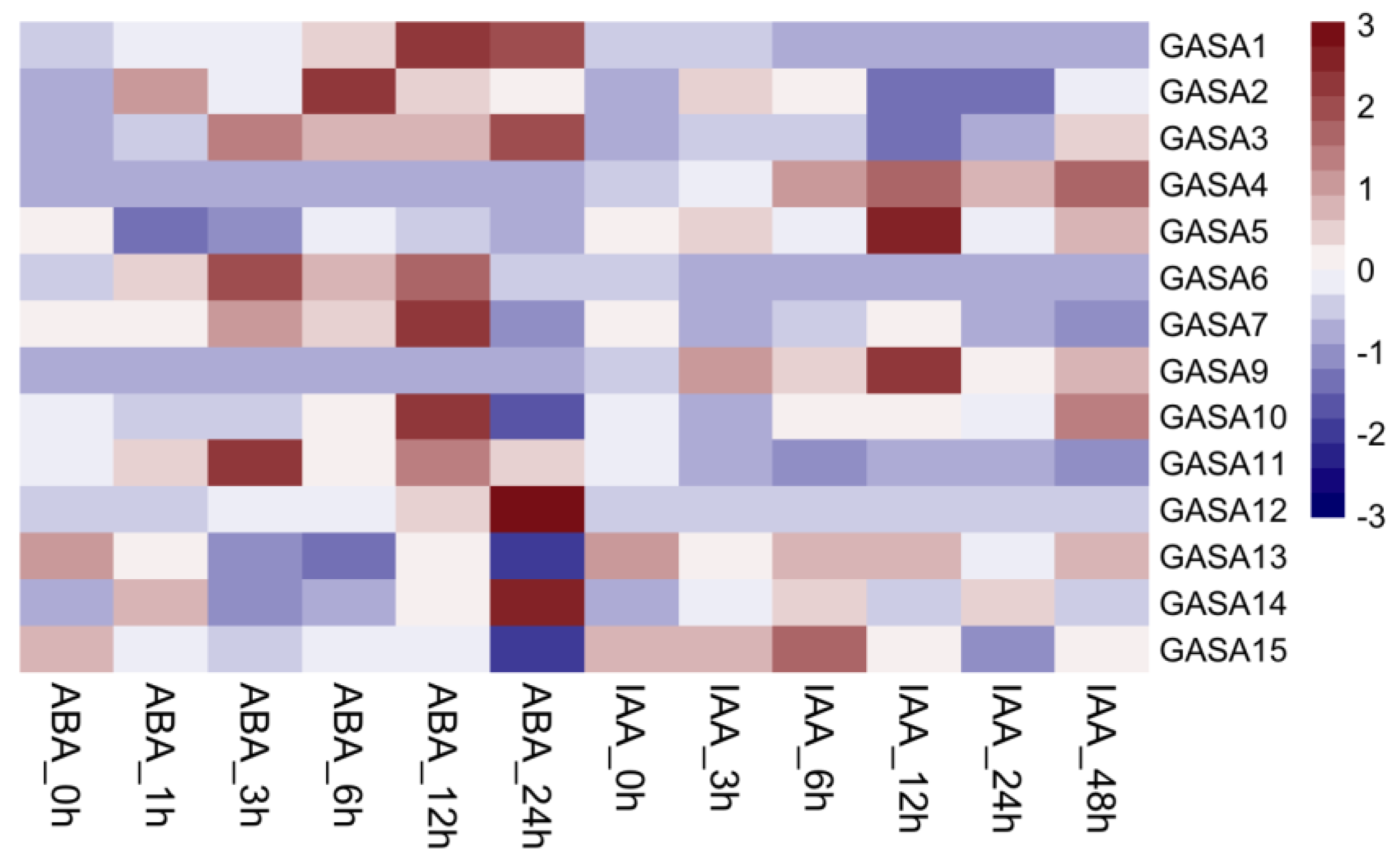
2.10. Expression Patterns of PmGASA Genes in Response to ABA and IAA Treatments
3. Discussion
4. Materials and Methods
4.1. Identification of GASA Genes in Six Plant Species
4.2. Physicochemical Property Analysis of PmGASA Proteins
4.3. Gene Structure and Protein Motif Analysis of PmGASA Genes
4.4. Phylogenetic Analysis of GASA Family Genes
4.5. Microsynteny and Selection Analysis of GASA Family Genes
4.6. Cis-Acting Element Analysis of PmGASA Genes
4.7. Tissue Specific Expression of PmGASA Genes
4.8. Expression Pattern of PmGASA Genes during Floral Bud Development
4.9. Plant Materials and qRT-PCR Analysis of PmGASA Genes in Gibberellin Treatment
4.10. Transcriptional Regulatory Network Prediction of PmGASAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, A.; Singh, A.; Kumar, P.; Sarkar, A.K. Giberellic Acid-Stimulated Transcript Proteins Evolved through Successive Conjugation of Novel Motifs and Their Subfunctionalization. Plant Physiol. 2019, 180, 998–1012. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Xu, Y.; Joo, S.H.; Kim, S.K.; Xue, Z.; Xu, Z.; Wang, Z.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhong, Y.; Chen, M.; Wu, B.; Wang, T.; Jiang, B.; Zhong, G. Analysis of CcGASA family members in Citrus clementina (Hort. ex Tan.) by a genome-wide approach. BMC Plant Biol. 2021, 21, 565. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Silverstein, K.A.T.; Moskal, W.A., Jr.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef]
- Shi, L.; Gast, R.T.; Gopalraj, M.; Olszewski, N.E. Characterization of a shoot-specific, GA3- and ABA-regulated gene from tomato. Plant J. 1992, 2, 153–159. [Google Scholar]
- Aubert, D.; Chevillard, M.; Dorne, A.M.; Arlaud, G.; Herzog, M. Expression patterns of GASA genes in Arabidopsis thaliana: The GASA4 gene is up-regulated by gibberellins in meristematic regions. Plant Mol.Biol. 1998, 36, 871–883. [Google Scholar] [CrossRef]
- Furukawa, T.; Sakaguchi, N.; Shimada, H. Two OsGASR genes, rice GAST homologue genes that are abundant in proliferating tissues, show different expression patterns in developing panicles. Genes Genet Syst. 2006, 81, 171–180. [Google Scholar] [CrossRef]
- Nahirñak, V.; Rivarola, M.; Gonzalez de Urreta, M.; Paniego, N.; Hopp, H.E.; Almasia, N.I.; Vazquez-Rovere, C. Genome-wide Analysis of the Snakin/GASA Gene Family in Solanum tuberosum cv. Kennebec. Am. J. Potato Res. 2016, 93, 172–188. [Google Scholar] [CrossRef]
- Han, S.; Jiao, Z.; Niu, M.X.; Yu, X.; Huang, M.; Liu, C.; Wang, H.L.; Zhou, Y.; Mao, W.; Wang, X.; et al. Genome-Wide Comprehensive Analysis of the GASA Gene Family in Populus. Int J Mol Sci 2021, 22, 12336. [Google Scholar] [CrossRef]
- García, A.N.; Ayub, N.D.; Fox, A.R.; Gómez, M.C.; Diéguez, M.J.; Pagano, E.M.; Berini, C.A.; Muschietti, J.P.; Soto, G. Alfalfa snakin-1 prevents fungal colonization and probably coevolved with rhizobia. BMC Plant Biol. 2014, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roxrud, I.; Lid, S.E.; Fletcher, J.C.; Schmidt, E.D.; Opsahl-Sorteberg, H.G. GASA4, one of the 14-member Arabidopsis GASA family of small polypeptides, regulates flowering and seed development. Plant Cell Physiol. 2007, 48, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, C.; Peng, J.; Sun, S.; Wang, X. GASA5, a regulator of flowering time and stem growth in Arabidopsis thaliana. Plant Mol. Biol. 2009, 69, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Trapalis, M.; Li, S.F.; Parish, R.W. The Arabidopsis GASA10 gene encodes a cell wall protein strongly expressed in developing anthers and seeds. Plant Sci. 2017, 260, 71–79. [Google Scholar] [CrossRef]
- Moyano-Cañete, E.; Bellido, M.L.; García-Caparrós, N.; Medina-Puche, L.; Amil-Ruiz, F.; González-Reyes, J.A.; Caballero, J.L.; Muñoz-Blanco, J.; Blanco-Portales, R. FaGAST2, a Strawberry Ripening-Related Gene, Acts Together with FaGAST1 to Determine Cell Size of the Fruit Receptacle. Plant Cell Physiol. 2012, 54, 218–236. [Google Scholar] [CrossRef]
- Kotilainen, M.; Helariutta, Y.; Mehto, M.; Pöllänen, E.; Albert, V.A.; Elomaa, P.; Teeri, T.H. GEG Participates in the Regulation of Cell and Organ Shape during Corolla and Carpel Development in Gerbera hybrida. Plant Cell 1999, 11, 1093–1104. [Google Scholar] [CrossRef]
- Han, M.; Jin, X.; Yao, W.; Kong, L.; Huang, G.; Tao, Y.; Li, L.; Wang, X.; Wang, Y. A Mini Zinc-Finger Protein (MIF) from Gerbera hybrida Activates the GASA Protein Family Gene, GEG, to Inhibit Ray Petal Elongation. Front. Plant Sci. 2017, 8, 1649. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, D.; Zhang, L.; Gao, C.; Xin, M.; Tahir, M.M.; Li, Y.; Ma, J.; Han, M. Comprehensive analysis of GASA family members in the Malus domestica genome: Identification, characterization, and their expressions in response to apple flower induction. BMC Genom. 2017, 18, 827. [Google Scholar] [CrossRef]
- Yang, Q.; Niu, Q.; Tang, Y.; Ma, Y.; Yan, X.; Li, J.; Tian, J.; Bai, S.; Teng, Y. PpyGAST1 is potentially involved in bud dormancy release by integrating the GA biosynthesis and ABA signaling in ‘Suli’ pear (Pyrus pyrifolia White Pear Group). Environ. Exp. Bot. 2019, 162, 302–312. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X. One new kind of phytohormonal signaling integrator: Up-and-coming GASA family genes. Plant Signal. Behav. 2017, 12, e1226453. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X. Expression pattern of GASA, downstream genes of DELLA, in Arabidopsis. Sci. Bull. 2008, 53, 3839–3846. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Liu, W.; Li, X.; Li, H. Overexpression of GmGASA32 promoted soybean height by interacting with GmCDC25. Plant Signal Behav. 2021, 16, 1855017. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Xu, H.; Ye, S.; Wang, S.; Li, L.; Zhang, S.; Wang, X. Gibberellic Acid-Stimulated Arabidopsis6 Serves as an Integrator of Gibberellin, Abscisic Acid, and Glucose Signaling during Seed Germination in Arabidopsis. Plant Physiol. 2015, 169, 2288–2303. [Google Scholar] [PubMed]
- Sun, S.; Wang, H.; Yu, H.; Zhong, C.; Zhang, X.; Peng, J.; Wang, X. GASA14 regulates leaf expansion and abiotic stress resistance by modulating reactive oxygen species accumulation. J. Exp. Bot. 2013, 64, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Wang, Q.; Zhang, X.; Zhang, B.; Luo, H.; He, C. Comprehensive transcriptional and functional analyses of HbGASA genes reveal their roles in fungal pathogen resistance in Hevea brasiliensis. Tree Genet Genomes 2018, 14, 41. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1–8. [Google Scholar] [CrossRef]
- Bailly, C. Anticancer properties of Prunus mume extracts (Chinese plum, Japanese apricot). J. Ethnopharmacol. 2020, 246, 112215. [Google Scholar] [CrossRef]
- Gao, Z.; Luo, W. Origin and Evolution of Prunus mume. In The Prunus mume Genome. Compendium of Plant Genomes; Springer: Cham, Switzerland, 2019; pp. 5–7. [Google Scholar]
- Zhang, M.; Yang, Q.; Yuan, X.; Yan, X.; Wang, J.; Cheng, T.; Zhang, Q. Integrating Genome-Wide Association Analysis With Transcriptome Sequencing to Identify Candidate Genes Related to Blooming Time in Prunus mume. Front. Plant Sci. 2021, 12, 1226. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Sana, A.; Jamil, A.; Nasir, J.A.; Ahmed, S.; Hameed, M.U.; Abdullah. A genome-wide approach to the comprehensive analysis of GASA gene family in Glycine max. Plant Mol. Biol. 2019, 100, 607–620. [Google Scholar] [CrossRef]
- Qiao, K.; Ma, C.; Lv, J.; Zhang, C.; Ma, Q.; Fan, S. Identification, characterization, and expression profiles of the GASA genes in cotton. J. Cotton Res. 2021, 4, 7. [Google Scholar] [CrossRef]
- Liu, Z.H.; Zhu, L.; Shi, H.Y.; Chen, Y.; Zhang, J.M.; Zheng, Y.; Li, X.B. Cotton GASL genes encoding putative gibberellin-regulated proteins are involved in response to GA signaling in fiber development. Mol. Biol. Rep. 2013, 40, 4561–4570. [Google Scholar] [CrossRef] [PubMed]
- Berrocal-Lobo, M.; Segura, A.; Moreno, M.; López, G.; García-Olmedo, F.; Molina, A. Snakin-2, an antimicrobial peptide from potato whose gene is locally induced by wounding and responds to pathogen infection. Plant Physiol. 2002, 128, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Yao, J.; Zhang, S.; Li, X.; Zhang, X.; Yadav, V.; Wang, X. Genome-Wide Characterization and Expression Profiling of GASA Genes during Different Stages of Seed Development in Grapevine (Vitis vinifera L.) Predict Their Involvement in Seed Development. Int. J. Mol. Sci. 2020, 21, 1088. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Sakai, H.; Hochholdinger, F. The Gibberellic Acid Stimulated-Like gene family in maize and its role in lateral root development. Plant Physiol. 2010, 152, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.-B.; Woo, Y.-M.; Lee, D.J.; Lee, M.-C.; Kim, C.S. Enhanced tolerance to heat stress in transgenic plants expressing the GASA4 gene. Plant Physiol. Biochem. 2007, 45, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2012, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Darby, N.; Creighton, T.E. Disulfide bonds in protein folding and stability. Methods Mol. Biol. 1995, 40, 219–252. [Google Scholar]
- Ben-Nissan, G.; Lee, J.Y.; Borohov, A.; Weiss, D. GIP, a Petunia hybrida GA-induced cysteine-rich protein: A possible role in shoot elongation and transition to flowering. Plant J. 2004, 37, 229–238. [Google Scholar] [CrossRef]
- Zhang, M.; Li, P.; Yan, X.; Wang, J.; Cheng, T.; Zhang, Q. Genome-wide characterization of PEBP family genes in nine Rosaceae tree species and their expression analysis in P. mume. BMC Ecol. Evol. 2021, 21, 1–23. [Google Scholar] [CrossRef]
- Meslin, C.; Mugnier, S.; Callebaut, I.; Laurin, M.; Pascal, G.; Poupon, A.; Goudet, G.; Monget, P. Evolution of genes involved in gamete interaction: Evidence for positive selection, duplications and losses in vertebrates. PLoS ONE 2012, 7, e44548. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.-y.; Cho, D.-I.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Bensmihen, S.; Giraudat, J.; Parcy, F. Characterization of three homologous basic leucine zipper transcription factors (bZIP) of the ABI5 family during Arabidopsis thaliana embryo maturation. J. Exp. Bot. 2005, 56, 597–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilfoyle, T.J.; Ulmasov, T.; Hagen, G. The ARF family of transcription factors and their role in plant hormone-responsive transcription. Cell Mol. Life Sci. 1998, 54, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud Dormancy in Perennial Fruit Tree Species: A Pivotal Role for Oxidative Cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- Zhang, M.; Suren, H.; Holliday, J.A.; Gaut, B. Phenotypic and Genomic Local Adaptation across Latitude and Altitude in Populus trichocarpa. Genome Biol. Evol. 2019, 11, 2256–2272. [Google Scholar] [CrossRef]
- Yang, Q.; Gao, Y.; Wu, X.; Moriguchi, T.; Bai, S.; Teng, Y. Bud endodormancy in deciduous fruit trees: Advances and prospects. Hortic. Res. 2021, 8, 139. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Hoffman, D.; Kaduk, M.; Mauriat, M.; Moritz, T. Transgenic hybrid aspen trees with increased gibberellin (GA) concentrations suggest that GA acts in parallel with FLOWERING LOCUS T2 to control shoot elongation. New Phytol. 2015, 205, 1288–1295. [Google Scholar] [CrossRef]
- Rinne, P.L.H.; Welling, A.; Vahala, J.; Ripel, L.; Ruonala, R.; Kangasjärvi, J.; van der Schoot, C. Chilling of Dormant Buds Hyperinduces FLOWERING LOCUS T and Recruits GA-Inducible 1,3-β-Glucanases to Reopen Signal Conduits and Release Dormancy in Populus. Plant Cell 2011, 23, 130–146. [Google Scholar] [CrossRef]
- Zhang, M.; Cheng, W.; Yuan, X.; Wang, J.; Cheng, T.; Zhang, Q. Integrated transcriptome and small RNA sequencing in revealing miRNA-mediated regulatory network of floral bud break in Prunus mume. Front. Plant Sci. 2022, 13, 931454. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids. Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Newbigin, E.; Chou, K.-C.; Shen, H.-B. Plant-mPLoc: A Top-Down Strategy to Augment the Power for Predicting Plant Protein Subcellular Localization. PLoS ONE 2010, 5, e11335. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids. Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Nicholas, K.B. GeneDoc: Analysis and visualization of genetic variation, EMBNEW. Embnew News 1997, 4, 14. [Google Scholar]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Stern, A.; Doron-Faigenboim, A.; Erez, E.; Martz, E.; Bacharach, E.; Pupko, T. Selecton 2007: Advanced models for detecting positive and purifying selection using a Bayesian inference approach. Nucleic Acids Res. 2007, 35, W506–W511. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome Profiles Reveal the Crucial Roles of Hormone and Sugar in the Bud Dormancy of Prunus mume. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2019, 48, D1104–D1113. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene_ID | Gene_Name | NCBI Accesion | Start | End | HMM Accession | HMM Name | E-Value (Pfam) |
|---|---|---|---|---|---|---|---|
| Pm005363 | PmGASA1 | XP_007223838.1 | 29 | 88 | PF02704 | GASA | 4.29 × 10−26 |
| Pm006215 | PmGASA2 | CAB4265575.1 | 50 | 109 | PF02704 | GASA | 3.41 × 10−27 |
| Pm009495 | PmGASA3 | XP_008224510.1 | 35 | 92 | PF02704 | GASA | 1.44 × 10−26 |
| Pm009496 | PmGASA4 | XP_008224511.1 | 36 | 95 | PF02704 | GASA | 8.24 × 10−25 |
| Pm014337 | PmGASA5 | XP_007215190.1 | 35 | 94 | PF02704 | GASA | 1.19 × 10−24 |
| Pm015762 | PmGASA6 | ONI19434.1 | 159 | 219 | PF02704 | GASA | 1.76 × 10−26 |
| Pm015883 | PmGASA7 | PQQ10068.1 | 49 | 108 | PF02704 | GASA | 2.24 × 10−18 |
| Pm021248 | PmGASA8 | KAH0977881.1 | 9 | 62 | PF02704 | GASA | 1.88 × 10−13 |
| Pm021249 | PmGASA9 | CAB4290623.1 | 13 | 60 | PF02704 | GASA | 2.52 × 10−26 |
| Pm022352 | PmGASA10 | XP_008237395.1 | 54 | 113 | PF02704 | GASA | 1.05 × 10−26 |
| Pm024681 | PmGASA11 | XP_008239716.1 | 62 | 122 | PF02704 | GASA | 3.03 × 10−27 |
| Pm024909 | PmGASA12 | XP_008239947.1 | 48 | 107 | PF02704 | GASA | 6.36 × 10−26 |
| Pm025746 | PmGASA13 | XP_008240742.1 | 57 | 116 | PF02704 | GASA | 5.82 × 10−31 |
| Pm026754 | PmGASA14 | XP_016651693.1 | 49 | 108 | PF02704 | GASA | 8.88 × 10−24 |
| Pm029238 | PmGASA15 | XP_008244953.1 | 53 | 112 | PF02704 | GASA | 2.15 × 10−30 |
| Pm030034 | PmGASA16 | XP_008245155.1 | 46 | 106 | PF02704 | GASA | 2.75 × 10−23 |
| Gene_Name | Protein Length (aa) | MW (Da) | pI | Instability Index | Aliphatic Index | GRAVY | Subcellular Prediction |
|---|---|---|---|---|---|---|---|
| PmGASA1 | 88 | 9613.29 | 8.58 | 35.77 | 59.77 | −0.064 | Golgi; Nucleus |
| PmGASA2 | 109 | 11,962.01 | 8.28 | 83.81 | 65.23 | −0.353 | Nucleus |
| PmGASA3 | 120 | 12,321.77 | 8.9 | 50.46 | 28.67 | −0.911 | Nucleus |
| PmGASA4 | 95 | 10,646.52 | 8.61 | 56.36 | 56.53 | −0.28 | Golgi |
| PmGASA5 | 94 | 10,595.56 | 9.09 | 43.35 | 61.17 | −0.181 | Golgi |
| PmGASA6 | 219 | 23,383.3 | 10 | 72.24 | 87.72 | −0.058 | Nucleus |
| PmGASA7 | 108 | 11,732.75 | 9.26 | 42.49 | 47.13 | −0.276 | Golgi |
| PmGASA8 | 108 | 12,081.8 | 8.12 | 76.86 | 44.26 | −0.867 | Golgi; Nucleus |
| PmGASA9 | 183 | 20,356.95 | 6.06 | 46.45 | 62.4 | −0.426 | Nucleus |
| PmGASA10 | 113 | 12,489.76 | 9.58 | 57.15 | 62.3 | −0.358 | Golgi |
| PmGASA11 | 122 | 13,454.97 | 9.16 | 43.61 | 79.02 | −0.018 | Nucleus |
| PmGASA12 | 107 | 11,856.05 | 9.11 | 39.51 | 57.48 | −0.176 | Golgi |
| PmGASA13 | 116 | 12,872.11 | 9.11 | 40.8 | 55.52 | −0.342 | Nucleus |
| PmGASA14 | 108 | 11,611.52 | 8.92 | 61.23 | 53.43 | −0.188 | Golgi |
| PmGASA15 | 112 | 12,392.59 | 9.02 | 39.73 | 58.39 | −0.178 | Golgi |
| PmGASA16 | 106 | 11,785.77 | 8.89 | 44.91 | 71.7 | −0.163 | Golgi; Nucleus |
| Gene 1 | Gene 2 | Ka | Ks | Ka/Ks | Selection Pressure | Gene Duplications |
|---|---|---|---|---|---|---|
| PmGASA3 | PmGASA4 | 0.36 | 1.17 | 0.31 | Purifying | Tandem |
| PmGASA1 | PmGASA8 | 0.39 | 8.17 | 0.05 | Purifying | WGD or Segmental |
| PmGASA2 | PmGASA14 | 0.41 | 1.67 | 0.24 | Purifying | WGD or Segmental |
| PmGASA6 | PmGASA11 | 0.43 | 3.39 | 0.13 | Purifying | WGD or Segmental |
| PmGASA7 | PmGASA12 | 0.16 | 0.92 | 0.18 | Purifying | WGD or Segmental |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Cheng, W.; Wang, J.; Cheng, T.; Zhang, Q. Genome-Wide Identification, Evolution, and Expression Analysis of GASA Gene Family in Prunus mume. Int. J. Mol. Sci. 2022, 23, 10923. https://doi.org/10.3390/ijms231810923
Zhang M, Cheng W, Wang J, Cheng T, Zhang Q. Genome-Wide Identification, Evolution, and Expression Analysis of GASA Gene Family in Prunus mume. International Journal of Molecular Sciences. 2022; 23(18):10923. https://doi.org/10.3390/ijms231810923
Chicago/Turabian StyleZhang, Man, Wenhui Cheng, Jia Wang, Tangren Cheng, and Qixiang Zhang. 2022. "Genome-Wide Identification, Evolution, and Expression Analysis of GASA Gene Family in Prunus mume" International Journal of Molecular Sciences 23, no. 18: 10923. https://doi.org/10.3390/ijms231810923