


The Characterization of Columnar Apple Gene MdCoL Promoter and Its Response to Abscisic Acid, Brassinosteroid and Gibberellic Acid

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
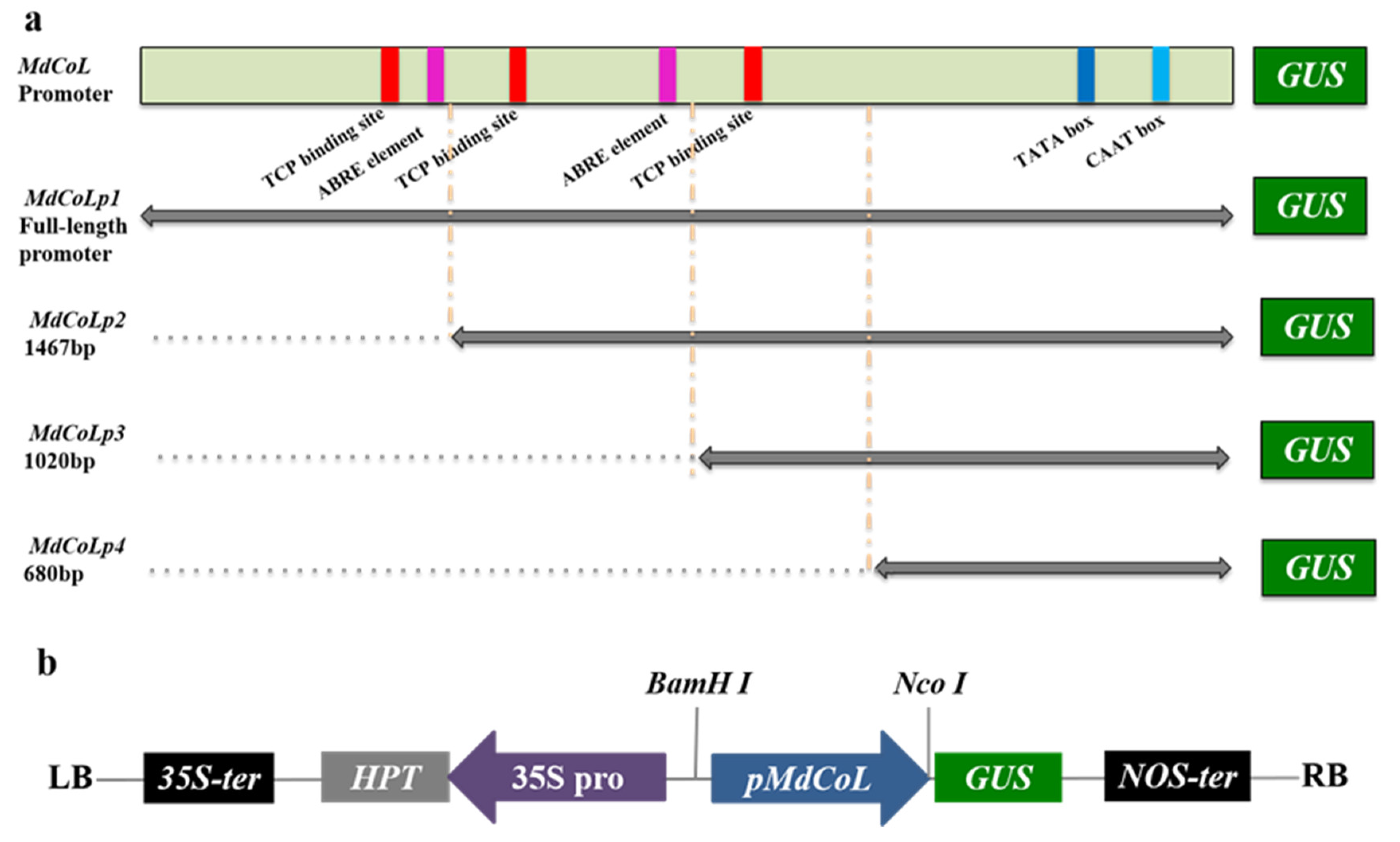
2.1. Isolation and Bioinformatics Analysis of MdCoL Promoter
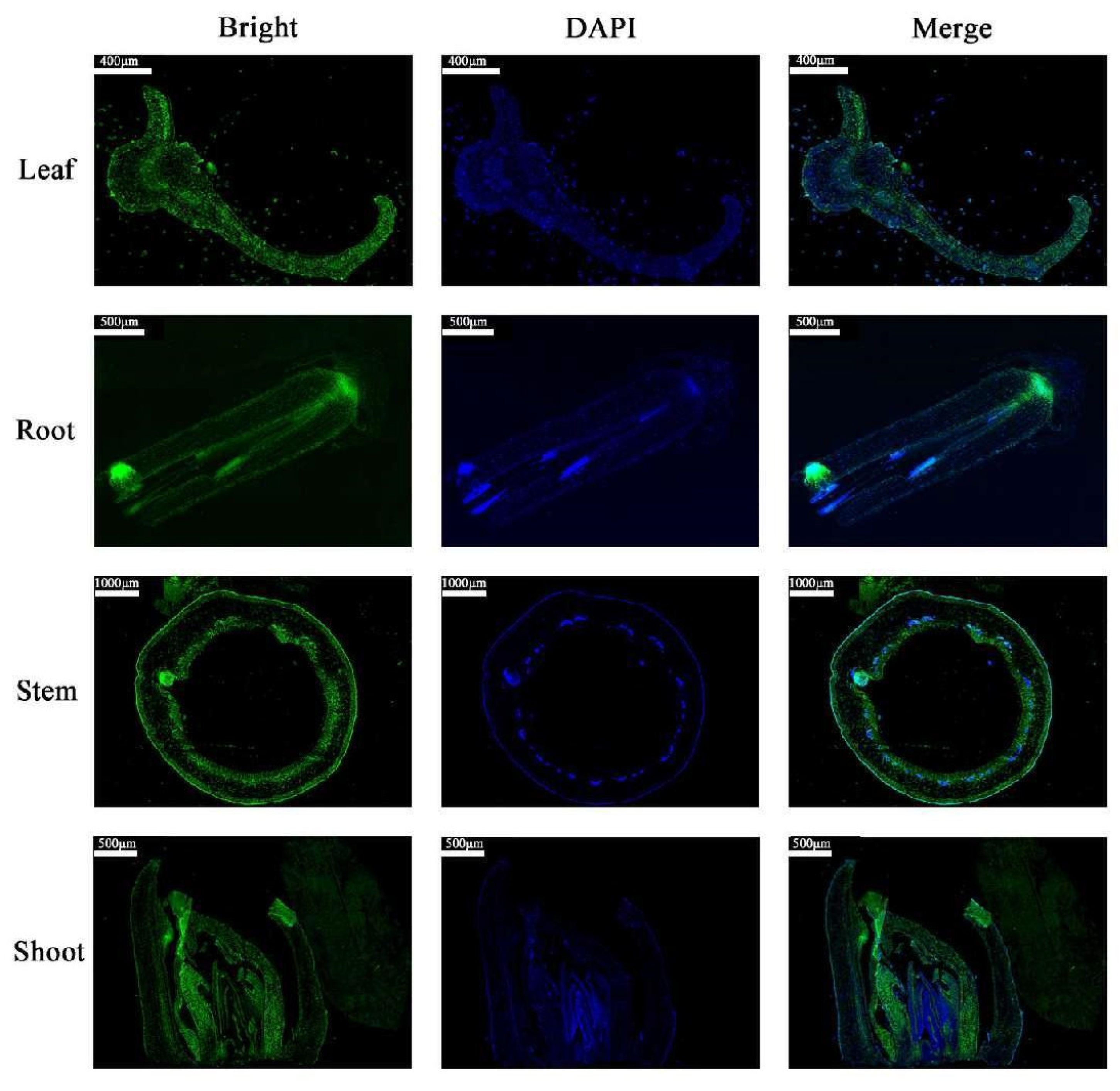
2.2. The Tissue Expression Characteristics of MdCoL in Columnar Apple
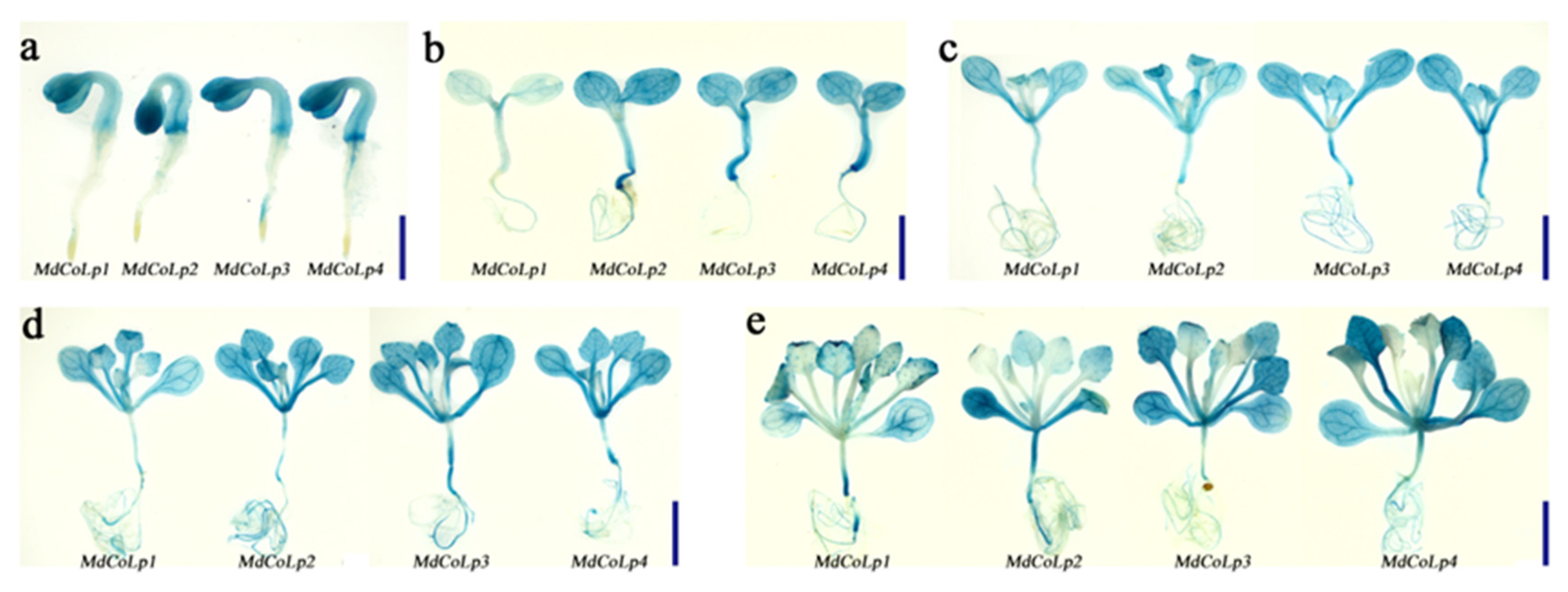
2.3. GUS Staining in Different Transgenic Arabidopsis Plants
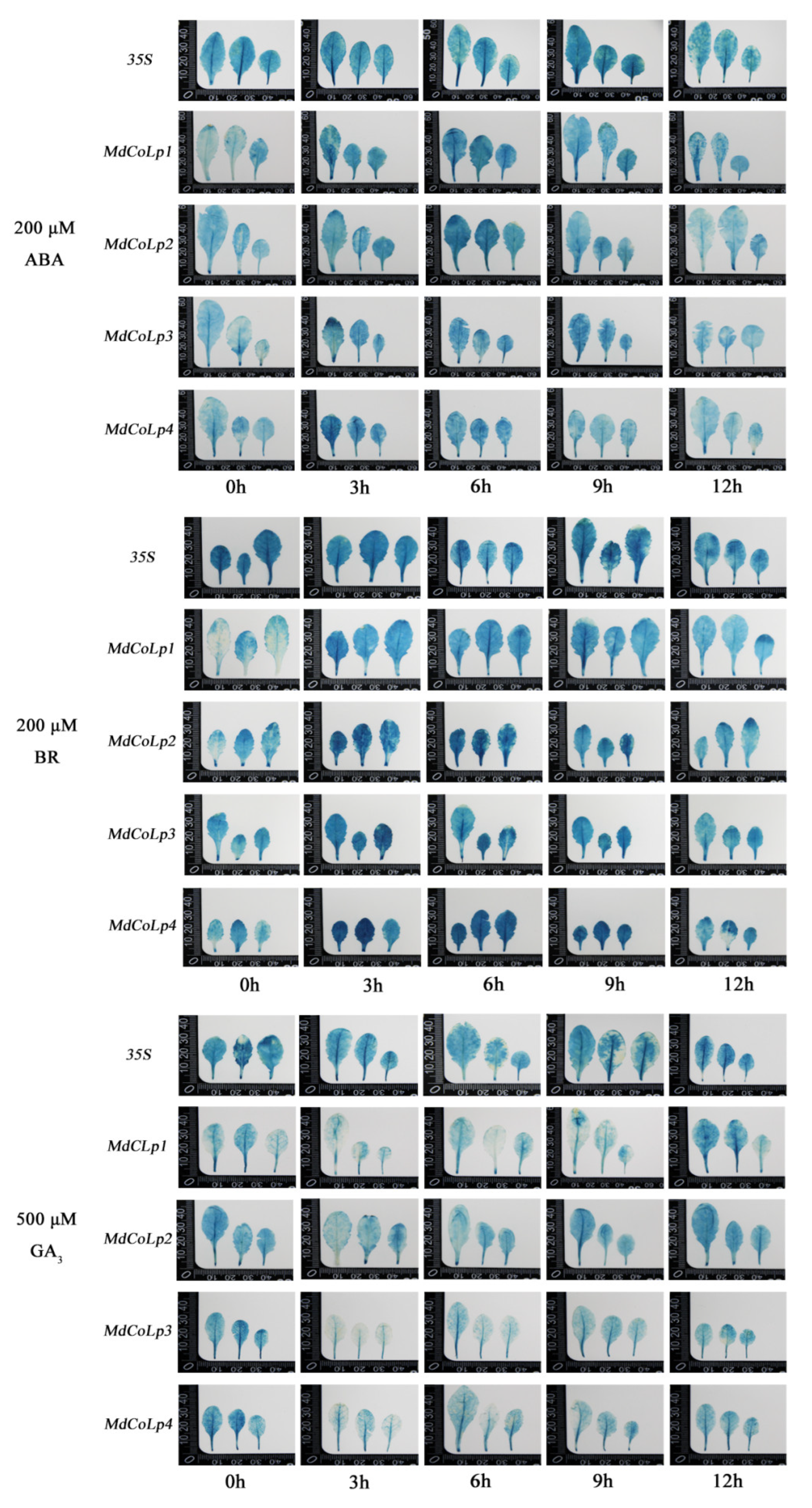
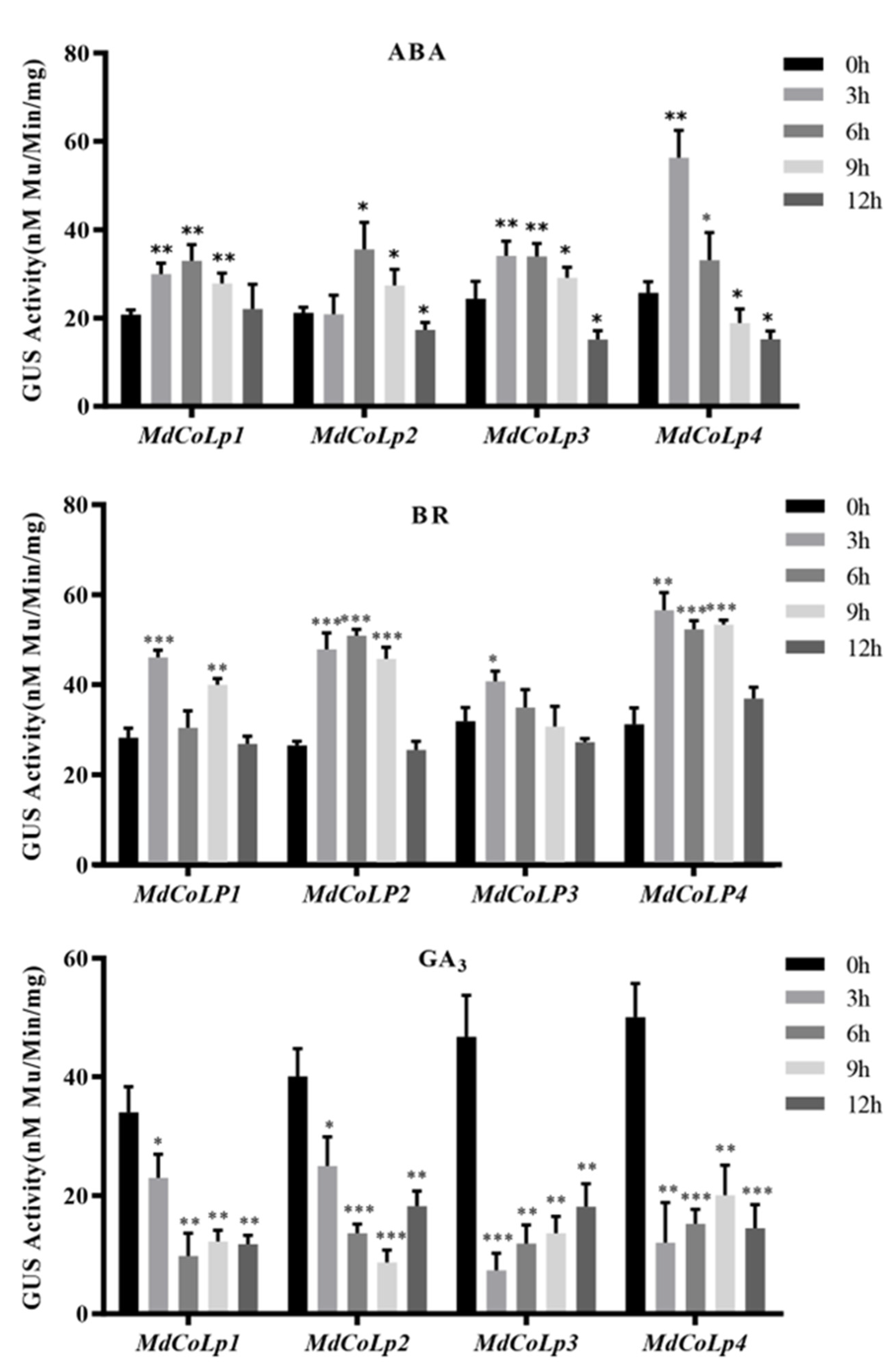
2.4. GUS Activity in Different Transgenic Arabidopsis Plants under Hormones Treatment
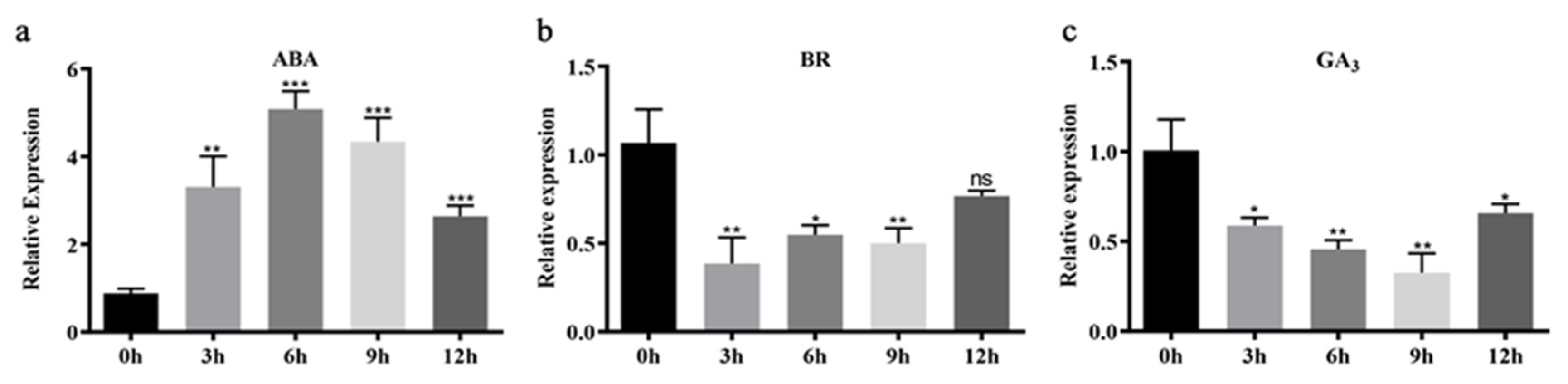
2.5. MdCoL Promoter Affected MdCoL Expression under Hormones Treatment
3. Discussions
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. In Situ Hybridization
4.3. Isolation and Bioinformatics Analysis of MdCoL Promoter
4.4. Construction of GUS Reporter Expression Vectors and Genetic Transformation of Arabidopsis
4.5. Histochemical GUS Staining
4.6. The Analysis of GUS Activity
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, B.; Smith, S.M.; Li, J.Y. Genetic regulation of shoot architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, D.F.; Brown, S.K. ‘McIntosh Wijcik’: A columnar mutation of ‘McIntosh’ apple proving useful in physiology and breeding research. Fruit Var. J. 1992, 46, 83–87. [Google Scholar]
- Lapins, K.O. Segregation of compact growth types in certain apple seedling progenies. Can. J. Plant Sci. 1969, 49, 765–768. [Google Scholar] [CrossRef]
- Tian, Y.K.; Wang, C.H.; Zhang, J.S.; James, C.; Dai, H.Y. Mapping Co, a gene controlling the columnar phenotype of apple, with molecular markers. Euphytica 2005, 145, 181–188. [Google Scholar] [CrossRef]
- Bai, T.; Zhu, Y.; Fernández-Fernández, F.; Keulemans, J.; Brown, S.; Xu, K. Fine genetic mapping of the Co locus controlling columnar growth habit in apple. Mol. Genet. Genom. 2012, 287, 437–450. [Google Scholar] [CrossRef]
- Petersen, R.; Krost, C. Tracing a key player in the regulation of plant architecture: The columnar growth habit of apple trees (Malus × domestica). Planta 2013, 238, 1–22. [Google Scholar] [CrossRef]
- Moriya, S.; Okada, K.; Haji, T.; Yamamoto, T.; Abe, K. Fine mapping of Co, a gene controlling columnar growth habit located on apple (Malus × domestica Borkh.) linkage group 10. Plant Breed. 2012, 131, 641–647. [Google Scholar] [CrossRef]
- Baldi, P.; Wolters, P.J.; Komjanc, M.; Viola, R.; Salvi, S. Genetic and physical characterisation of the locus controlling columnar habit in apple (Malus × domestica Borkh.). Mol. Breed. 2013, 1, 429–440. [Google Scholar] [CrossRef]
- Savel’ev, N.I.; Lyzhin, A.S.; Kudryavtzev, A.M.; Boris, K.V.; Shamshin, I.N. Use of molecular markers for identification of genotypes of columnar apple trees. Russ. Agric. Sci. 2015, 41, 323–325. [Google Scholar] [CrossRef]
- Wolters, P.J.; Schouten, H.J.; Velasco, R.; Si-Ammour, A.; Baldi, P. Evidence for regulation of columnar habit in apple by a putative 2OG-Fe(II) oxygenase. New Phytol. 2013, 200, 993–999. [Google Scholar] [CrossRef]
- Otto, D.; Petersen, R.; Brauksiepe, B.; Braun, P.; Schmidt, E.R. The columnar mutation (“Co gene”) of apple (Malus × domestica) is associated with an integration of a Gypsy-like retrotransposon. Mol. Breed. 2014, 33, 863–880. [Google Scholar] [CrossRef]
- Okada, K.; Wada, M.; Moriya, S.; Katayose, Y.; Fujisawa, H.; Wu, J.Z.; Kanamori, H.; Kurita, K.; Sasaki, H.; Fujii, H.; et al. Expression of a putative dioxygenase gene adjacent to an insertion mutation is involved in the short internodes of columnar apples (Malus × domestica). J. Plant Res. 2016, 129, 1109–1126. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wen, C.P.; Xu, J.H.; Wang, Y.H.; Zhu, J.; Zhang, Y.G. The apple columnar gene candidate MdCoL and the AP2/ERF factor MdDREB2 positively regulate ABA biosynthesis by activating the expression of MdNCED6/9. Tree Physiol. 2021, 41, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Finlayson, S.A. Abscisic acid is a general negative regulator of Arabidopsis axillary bud growth. Plant Physiol. 2015, 169, 611–626. [Google Scholar] [CrossRef] [PubMed]
- González-Grandío, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancón, C.; Immink, R.G.H.; Cubas, P. Abscisic acid signaling is controlled by a BRANCHED1/HD-ZIP I cascade in Arabidopsis axillary buds. Proc. Natl. Acad. Sci. USA 2017, 114, E245–E254. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.J.; Chu, Y.; Wang, Y.; Diao, Y.Y.; Zhao, Y.Y.; Liu, L.S.; Wei, X.; Meng, Y.; Li, F.G.; Ge, X.Y. The miR164-GhCUC2-GhBRC1 module regulates plant architecture through abscisic acid in cotton. Plant Biotechnol. J. 2021, 19, 1839–1851. [Google Scholar] [CrossRef]
- Bömke, C.; Tudzynski, B. Diversity, regulation, and evolution of the gibberellin biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry 2009, 70, 1876–1893. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.M.; Tan, B.; Jiang, Y.J.; Zheng, X.B.; Ye, X.; Guo, Z.J.; Xiong, T.T.; Wang, W.; Li, J.D.; et al. A single nucleotide mutation in GID1c disrupts its interaction with DELLA1 and causes a GA-insensitive dwarf phenotype in peach. Plant Biotechnol. J. 2019, 17, 1723–1735. [Google Scholar] [CrossRef]
- Singh, A.P.; Savaldi-Goldstein, S. Growth control: Brassinosteroid activity gets context. J. Exp. Bot. 2015, 66, 1123–1132. [Google Scholar] [CrossRef]
- Hu, J.; Sun, S.Y.; Wang, X.L. Regulation of shoot branching by strigolactones and brassinosteroids: Conserved and specific functions of Arabidopsis BES1 and rice BZR1. Mol. Plant 2020, 13, 808–810. [Google Scholar] [CrossRef]
- Mori, M.; Nomura, T.; Ooka, H.; Ishizaka, M.; Yokota, T.; Sugimoto, K.; Okabe, K.; Kajiwara, H.; Satoh, K.; Yamamoto, K.; et al. Isolation and characterization of a rice dwarf mutant with a defect in brassinosteroid biosynthesis. Plant Physiol. 2002, 130, 1152–1161. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.N.; Jin, Y.; Liu, W.B.; Li, F.; Fang, J.; Yin, Y.H.; Qian, Q.; Zhu, L.H.; Chu, C.C. DWARF AND LOW-TILLERING, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.J.; Dong, H.; Yin, Y.L.; Song, X.W.; Gu, X.H.; Sang, K.Q.; Zhou, J.; Shi, K.; Zhou, Y.H.; Foyer, C.H.; et al. Brassinosteroid signaling integrates multiple pathways to release apical dominance in tomato. Proc. Natl. Acad. Sci. USA 2021, 118, e2004384118. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Z.; Xu, Y.Y.; Joo, S.H.; Kim, S.K.; Xue, Z.; Xu, Z.H.; Wang, Z.Y.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Xu, F.; Chen, Z.; Teng, Z.F.; Sun, K.; Li, X.C.; Yu, J.Y.; Zhang, G.X.; Liang, Y.; Huang, X.H.; et al. Synergistic interplay of ABA and BR signal in regulating plant growth and adaptation. Nat. Plants 2021, 7, 1108–1118. [Google Scholar] [CrossRef]
- Liu, Y.J.; An, J.P.; Wang, X.F.; Gao, N.; Wang, X.; Zhang, S.; Gao, W.S.; Hao, Y.J.; You, C.X. MdBZR1 regulates ABA response by modulating the expression of MdABI5 in apple. Plant Cell Rep. 2021, 40, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, M.; Huibers, R.P.; Elberse, J.; Van den Ackerveken, G. Arabidopsis DMR6 encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility to downy mildew. Plant J. 2008, 54, 785–793. [Google Scholar] [CrossRef]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van den Ackerveken, G. Downy mildew resistant 6 and dmr6-like oxygenase 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2015, 81, 210–222. [Google Scholar] [CrossRef]
- Falcone, M.L.; Emiliani, J.; Rodriguez, E.J.; Campos-Bermudez, V.A.; Grotewold, E.; Casati, P. The Identification of Maize and Arabidopsis Type I Flavone Synthases Links Flavones with Hormones and Biotic Interactions. Plant Physiol. 2015, 169, 1090–1107. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.J.; Zhang, K. S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase That Fine-Tunes Salicylic Acid Homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef]
- Wada, M.; Iwanami, H.; Moriya, S.; Hanada, T.; Moriya-Tanaka, Y.; Honda, C.; Shimizu, T.; Abe, K.; Okada, K. A root-localized gene in normal apples is ectopically expressed in aerial parts of columnar apples. Plant Growth Regul. 2018, 85, 389–398. [Google Scholar] [CrossRef]
- Rutjens, B.; Bao, D.P.; Van Eck-Stouten, E.; Brand, M.; Smeekens, S.; Proveniers, M. Shoot apical meristem function in Arabidopsis requires the combined activities of three BEL1-like homeodomain proteins. Plant J. 2009, 58, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Braun, N.; de Saint Germain, A.; Pillot, J.P.; Boutet-Mercey, S.; Dalmais, M.; Antoniadi, I.; Li, X.; Maia-Grondard, A.; Le Signor, C.; Bouteiller, N.; et al. The pea TCP transcription factor PsBRC1 acts downstream of strigolactones to control shoot branching. Plant Physiol. 2012, 158, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.B.; Xiao, Y.G.; Govindarajulu, R.; Feil, R.; Siddoway, M.L.; Nielsen, T.; Lunn, J.E.; Hawkins, J.; Whipple, C.; Chuck, G. The regulatory landscape of a core maize domestication module controlling bud dormancy and growth repression. Nat. Commun. 2019, 10, 3810. [Google Scholar] [CrossRef]
- Dixon, L.E.; Pasquariello, M.; Boden, S.A. TEOSINTE BRANCHED1 regulates height and stem internode length in bread wheat. J. Exp. Bot. 2020, 71, 4742–4750. [Google Scholar] [CrossRef]
- Shen, J.J.; Zhang, Y.Q.; Ge, D.F.; Wang, Z.Y.; Song, W.Y.; Gu, R.; Che, G.; Cheng, Z.H.; Liu, R.Y.; Zhang, X.L. CsBRC1 inhibits axillary bud outgrowth by directly repressing the auxin efflux carrier CsPIN3 in cucumber. Proc. Natl. Acad. Sci. USA 2019, 116, 17105–17114. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Takahashi, I.; Jaroensanti-Tanaka, N.; Miyazaki, S.; Jiang, K.; Nakayasu, M.; Wada, M.; Asami, T.; Mizutani, M.; Okada, K.; et al. The apple gene responsible for columnar tree shape reduces the abundance of biologically active gibberellin. Plant J. 2021, 105, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.L.; Hollender, C.A. Branching out: New insights into the genetic regulation of shoot architecture in trees. Curr. Opin. Plant Biol. 2019, 47, 73–80. [Google Scholar] [CrossRef]
- Wang, H.J.; Tang, J.; Liu, J.; Hu, J.; Liu, J.J.; Chen, Y.X.; Chen, Z.Y.; Wang, X.L. Abscisic acid signaling inhibits brassinosteroid signaling through dampening the dephosphorylation of BIN2 by ABI1 and ABI2. Mol. Plant 2018, 11, 315–325. [Google Scholar] [CrossRef]
- Okada, K.; Wada, M.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Nakayasu, M.; Mizutani, M.; Nakajima, M.; Moriya, S.; Shimizu, T.; et al. Columnar growth phenotype in apple results from gibberellin deficiency by ectopic expression of a dioxygenase gene. Tree Physiol. 2020, 40, 1205–1216. [Google Scholar] [CrossRef]
- Schmidt, R.; Schippers, J.H.M.; Mieulet, D.; Obata, T.; Fernie, A.R.; Guiderdoni, E.; Mueller-Roeber, B. MULTIPASS, a rice R2R3-type MYB transcription factor, regulates adaptive growth by integrating multiple hormonal pathways. Plant J. 2013, 76, 258–273. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.F.; Liu, X.Y.; Du, H.; Dai, M.Q.; Zhou, D.X.; Yang, W.J.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.H.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; Ureshino, A.; Tanaka, N.; Komori, S.; Takahashi, S.; Kudo, K.; Bessho, H. Anatomical analysis by two approaches ensure the promoter activities of apple AFL genes. J. Jpn. Soc. Hortic. Sci. 2009, 78, 32–39. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, L.N.; Zeng, Z.R.; Wu, C.Z.; Gong, Y.Y.; Liu, L.H.; Cao, F.Q. Molecular identification of a root apical cell-specific and stress-responsive enhancer from an Arabidopsis enhancer trap line. Plant Methods 2019, 15, 8. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.P.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, T.; Yu, J.; Zhuang, J.; Wang, Z.; Sun, X.; Zhang, Y. The Characterization of Columnar Apple Gene MdCoL Promoter and Its Response to Abscisic Acid, Brassinosteroid and Gibberellic Acid. Int. J. Mol. Sci. 2022, 23, 10781. https://doi.org/10.3390/ijms231810781
Han T, Yu J, Zhuang J, Wang Z, Sun X, Zhang Y. The Characterization of Columnar Apple Gene MdCoL Promoter and Its Response to Abscisic Acid, Brassinosteroid and Gibberellic Acid. International Journal of Molecular Sciences. 2022; 23(18):10781. https://doi.org/10.3390/ijms231810781
Chicago/Turabian StyleHan, Tingting, Jiahui Yu, Jie Zhuang, Ziyu Wang, Xin Sun, and Yugang Zhang. 2022. "The Characterization of Columnar Apple Gene MdCoL Promoter and Its Response to Abscisic Acid, Brassinosteroid and Gibberellic Acid" International Journal of Molecular Sciences 23, no. 18: 10781. https://doi.org/10.3390/ijms231810781