
Epigenetic Dynamics and Regulation of Plant Male Reproduction


Abstract
:1. Introduction
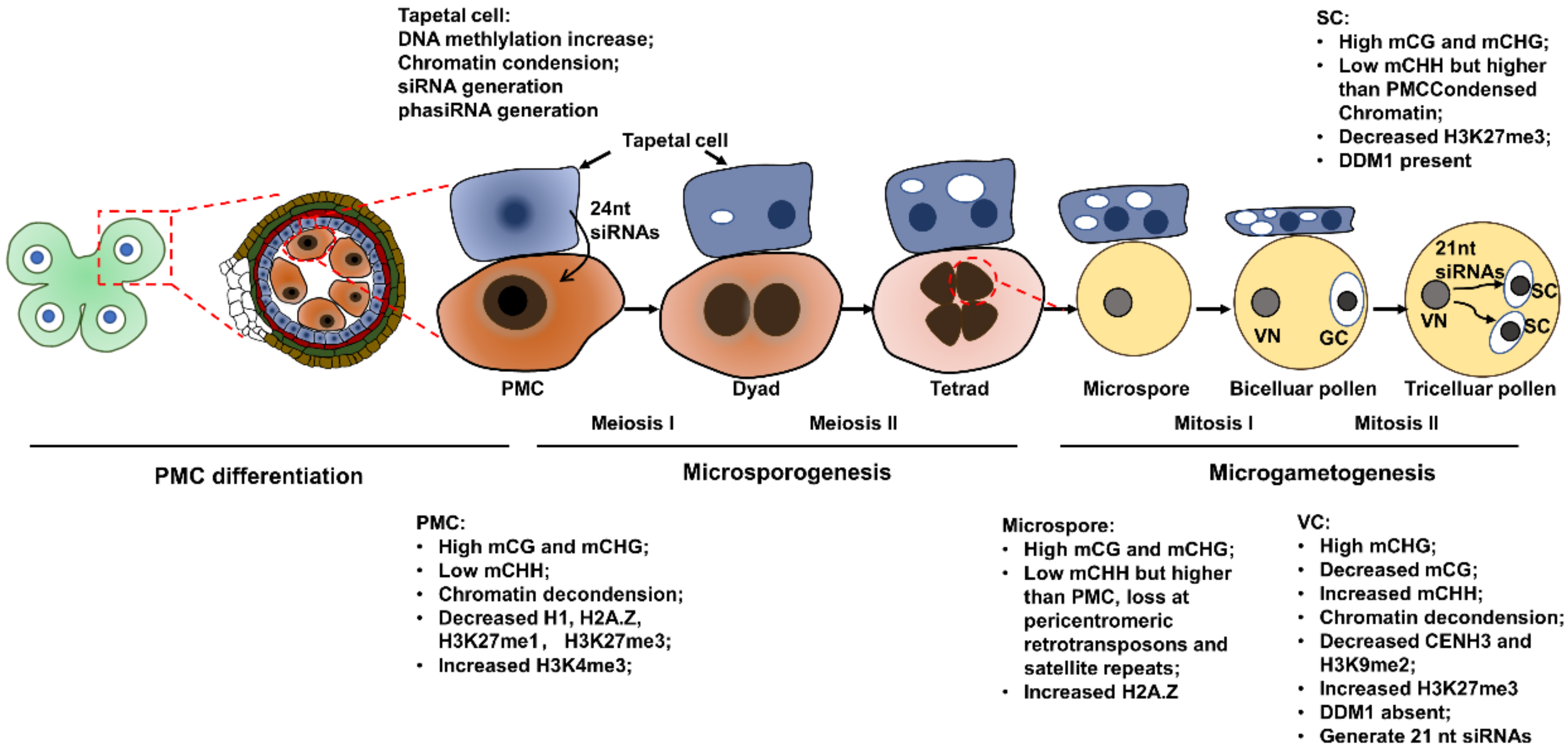
2. Epigenetic Dynamics during Pollen Mother Cell Differentiation
3. Epigenetic Dynamics during Microsporogenesis
4. Epigenetic Dynamics at Microgametogenesis
5. Epigenetic Dynamics in Tapetal Cell Development
6. Epigenetic Variations between Male Fertile and Sterile Lines
7. Epigenetic Regulation-Related Genes That Are Involved in Pollen Development
8. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, X.; Dickinson, H.G. Tapetal cell fate, lineage and proliferation in the Arabidopsis anther. Development 2010, 137, 2409–2416. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.; Taylor, M.L.; O’Meara, B.C. Repeated evolution of tricellular (and bicellular) pollen. Am. J. Bot. 2014, 101, 559–571. [Google Scholar] [CrossRef]
- Liu, L.; Fan, X.D. Tapetum: Regulation and role in sporopollenin biosynthesis in Arabidopsis. Plant Mol. Biol. 2013, 83, 165–175. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Zhao, T.; Zhan, Z.; Jiang, D. Histone modifications and their regulatory roles in plant development and environmental memory. J. Genet. Genom. 2019, 46, 467–476. [Google Scholar] [CrossRef]
- Ito, T.; Wellmer, F.; Yu, H.; Das, P.; Ito, N.; Alves-Ferreira, M.; Riechmann, J.L.; Meyerowitz, E.M. The homeotic protein AGAMOUS controls microsporogenesis by regulation of SPOROCYTELESS. Nature 2004, 430, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Schiefthaler, U.; Balasubramanian, S.; Sieber, P.; Chevalier, D.; Wisman, E.; Schneitz, K. Molecular analysis of NOZZLE, a gene involved in pattern formation and early sporogenesis during sex organ development in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1999, 96, 11664–11669. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.C.; Ye, D.; Xu, J.; Sundaresan, V. The SPOROCYTELESS gene of Arabidopsis is required for initiation of sporogenesis and encodes a novel nuclear protein. Genes Dev. 1999, 13, 2108–2117. [Google Scholar] [CrossRef] [PubMed]
- Hord, C.L.; Chen, C.; Deyoung, B.J.; Clark, S.E.; Ma, H. The BAM1/BAM2 receptor-like kinases are important regulators of Arabidopsis early anther development. Plant Cell 2006, 18, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, C.; Russinova, E.; Hecht, V.; Baaijens, E.; de Vries, S. The Arabidopsis thaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES1 and 2 control male sporogenesis. Plant Cell 2005, 17, 3337–3349. [Google Scholar] [CrossRef] [Green Version]
- Colcombet, J.; Boisson-Dernier, A.; Ros-Palau, R.; Vera, C.E.; Schroeder, J.I. Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASES1 and 2 are essential for tapetum development and microspore maturation. Plant Cell 2005, 17, 3350–3361. [Google Scholar] [CrossRef]
- Yang, S.L.; Jiang, L.; Puah, C.S.; Xie, L.F.; Zhang, X.Q.; Chen, L.Q.; Yang, W.C.; Ye, D. Overexpression of TAPETUM DETERMINANT1 alters the cell fates in the Arabidopsis carpel and tapetum via genetic interaction with excess microsporocytes1/extra sporogenous cells. Plant Physiol. 2005, 139, 186–191. [Google Scholar] [CrossRef]
- Lou, Y.; Zhou, H.S.; Han, Y.; Zeng, Q.Y.; Zhu, J.; Yang, Z.N. Positive regulation of AMS by TDF1 and the formation of a TDF1-AMS complex are required for anther development in Arabidopsis thaliana. New Phytol. 2018, 217, 378–391. [Google Scholar] [CrossRef]
- Han, Y.; Zhou, S.D.; Fan, J.J.; Zhou, L.; Shi, Q.S.; Zhang, Y.F.; Liu, X.L.; Chen, X.; Zhu, J.; Yang, Z.N. OsMS188 Is a Key Regulator of Tapetum Development and Sporopollenin Synthesis in Rice. Rice 2021, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Park, K.S.; Twell, D.; Park, S.K. The Sidecar Pollen gene encodes a microspore-specific LOB/AS2 domain protein required for the correct timing and orientation of asymmetric cell division. Plant J. 2010, 64, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Hoai, T.N.T.; Park, H.J.; Zhao, M.; Twell, D.; Honys, D.; Park, S.K. MYB81, a microspore-specific GAMYB transcription factor, promotes pollen mitosis I and cell lineage formation in Arabidopsis. Plant J. Cell Mol. Biol. 2020, 101, 590–603. [Google Scholar] [CrossRef]
- Huang, J.; Dong, J.; Qu, L.-J. From birth to function: Male gametophyte development in flowering plants. Curr. Opin. Plant Biol. 2021, 63, 102118. [Google Scholar] [CrossRef] [PubMed]
- Twell, D. Male gametogenesis and germline specification in flowering plants. Sex Plant Reprod. 2011, 24, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Walbot, V.; Egger, R.L. Pre-Meiotic Anther Development: Cell Fate Specification and Differentiation. Annu. Rev. Plant Biol. 2016, 67, 365–395. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Dickinson, H.G. Packaging the male germline in plants. Trends Genet. 2007, 23, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, X.; Yuan, C.; Harrison, A.P.; Chen, M. The roles of cross-talk epigenetic patterns in Arabidopsis thaliana. Brief. Funct. Genom. 2016, 15, 278–287. [Google Scholar] [CrossRef]
- Hou, Q.; Wan, X. Epigenome and Epitranscriptome: Potential Resources for Crop Improvement. Int. J. Mol. Sci. 2021, 22, 12912. [Google Scholar] [CrossRef]
- Yang, H.; Lu, P.; Wang, Y.; Ma, H. The transcriptome landscape of Arabidopsis male meiocytes from high-throughput sequencing: The complexity and evolution of the meiotic process. Plant J. 2011, 65, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Farmer, A.D.; Langley, R.J.; Mudge, J.; Crow, J.A.; May, G.D.; Huntley, J.; Smith, A.G.; Retzel, E.F. Meiosis-specific gene discovery in plants: RNA-Seq applied to isolated Arabidopsis male meiocytes. BMC Plant Biol. 2010, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Hood, L.; Galas, D. The digital code of DNA. Nature 2003, 421, 444–448. [Google Scholar] [CrossRef] [PubMed]
- March-Díaz, R.; Reyes, J.C. The beauty of being a variant: H2A.Z and the SWR1 complex in plants. Mol. Plant 2009, 2, 565–577. [Google Scholar] [CrossRef] [PubMed]
- She, W.; Baroux, C. Chromatin dynamics in pollen mother cells underpin a common scenario at the somatic-to-reproductive fate transition of both the male and female lineages in Arabidopsis. Front. Plant Sci. 2015, 6, 294. [Google Scholar] [CrossRef]
- Walker, J.; Gao, H.; Zhang, J.; Aldridge, B.; Vickers, M.; Higgins, J.D.; Feng, X. Sexual-lineage-specific DNA methylation regulates meiosis in Arabidopsis. Nat. Genet. 2018, 50, 130–137. [Google Scholar] [CrossRef]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nat. Rev. Genet. 2014, 15, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Walker, J.; She, W.; Aldridge, B.; Gao, H.; Deans, S.; Vickers, M.; Feng, X. Nurse cell—Derived small RNAs define paternal epigenetic inheritance in Arabidopsis. Science 2021, 373, abh0556. [Google Scholar] [CrossRef]
- Huang, J.; Wang, C.; Wang, H.; Lu, P.; Zheng, B.; Ma, H.; Copenhaver, G.P.; Wang, Y. Meiocyte-Specific and AtSPO11-1-Dependent Small RNAs and Their Association with Meiotic Gene Expression and Recombination. Plant Cell 2019, 31, 444–464. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Fukushima, S.; Takatsuji, H. RNA-directed DNA methylation induces transcriptional activation in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 1660–1665. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.G. 11—Plant Embryology. In Plant Systematics, 3rd ed.; Simpson, M.G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 567–582. [Google Scholar]
- Nelms, B.; Walbot, V. Gametophyte genome activation occurs at pollen mitosis I in maize. Science 2022, 375, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.D.; Osman, K.; Jones, G.H.; Franklin, F.C. Factors underlying restricted crossover localization in barley meiosis. Annu. Rev. Genet. 2014, 48, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Gore, M.A.; Chia, J.M.; Elshire, R.J.; Sun, Q.; Ersoz, E.S.; Hurwitz, B.L.; Peiffer, J.A.; McMullen, M.D.; Grills, G.S.; Ross-Ibarra, J.; et al. A first-generation haplotype map of maize. Science 2009, 326, 1115–1117. [Google Scholar] [CrossRef]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Hellsten, U.; Wright, K.M.; Jenkins, J.; Shu, S.; Yuan, Y.; Wessler, S.R.; Schmutz, J.; Willis, J.H.; Rokhsar, D.S. Fine-scale variation in meiotic recombination in Mimulus inferred from population shotgun sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 19478–19482. [Google Scholar] [CrossRef]
- Wijnker, E.; Velikkakam James, G.; Ding, J.; Becker, F.; Klasen, J.R.; Rawat, V.; Rowan, B.A.; de Jong, D.F.; de Snoo, C.B.; Zapata, L.; et al. The genomic landscape of meiotic crossovers and gene conversions in Arabidopsis thaliana. eLife 2013, 2, e01426. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Zhao, X.; Kelly, K.A.; Venn, O.; Higgins, J.D.; Yelina, N.E.; Hardcastle, T.J.; Ziolkowski, P.A.; Copenhaver, G.P.; Franklin, F.C.H.; et al. Arabidopsis meiotic crossover hot spots overlap with H2A.Z nucleosomes at gene promoters. Nat. Genet. 2013, 45, 1327–1336. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, F.F.; Wu, Y.T.; Zhou, X.; Huang, X.Y.; Zhu, J.; Gao, J.F.; Dong, R.B.; Cao, K.M.; Yang, Z.N. MULTIPOLAR SPINDLE 1 (MPS1), a novel coiled-coil protein of Arabidopsis thaliana, is required for meiotic spindle organization. Plant J. Cell Mol. Biol. 2009, 59, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Pina, C.; Pinto, F.; Feijó, J.A.; Becker, J.D. Gene family analysis of the Arabidopsis pollen transcriptome reveals biological implications for cell growth, division control, and gene expression regulation. Plant Physiol. 2005, 138, 744–756. [Google Scholar] [CrossRef]
- Houben, A.; Kumke, K.; Nagaki, K.; Hause, G. CENH3 distribution and differential chromatin modifications during pollen development in rye (Secale cereale L.). Chromosome Res. 2011, 19, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.D.; Gou, X.; Wong, C.E.; Wang, X.; Yuan, T.; Wei, X.; Bhalla, P.L.; Singh, M.B. Genomic profiling of rice sperm cell transcripts reveals conserved and distinct elements in the flowering plant male germ lineage. New Phytol. 2012, 195, 560–573. [Google Scholar] [CrossRef]
- Borges, F.; Gomes, G.; Gardner, R.; Moreno, N.; McCormick, S.; Feijó, J.A.; Becker, D., Jr. Comparative Transcriptomics of Arabidopsis Sperm Cells. Plant Physiol. 2008, 148, 1168–1181. [Google Scholar] [CrossRef] [PubMed]
- Calarco, J.P.; Borges, F.; Donoghue, M.T.; Van Ex, F.; Jullien, P.E.; Lopes, T.; Gardner, R.; Berger, F.; Feijo, J.A.; Becker, J.D.; et al. Reprogramming of DNA methylation in pollen guides epigenetic inheritance via small RNA. Cell 2012, 151, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Schoft, V.K.; Chumak, N.; Mosiolek, M.; Slusarz, L.; Komnenovic, V.; Brownfield, L.; Twell, D.; Kakutani, T.; Tamaru, H. Induction of RNA-directed DNA methylation upon decondensation of constitutive heterochromatin. EMBO Rep. 2009, 10, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Vaughn, M.; Borges, F.; Tanurdzic, M.; Becker, J.D.; Feijo, J.A.; Martienssen, R.A. Epigenetic reprogramming and small RNA silencing of transposable elements in pollen. Cell 2009, 136, 461–472. [Google Scholar] [CrossRef]
- Huang, X.; Sun, M.X. H3K27 methylation regulates the fate of two cell lineages in male gametophytes. Plant Cell 2022, 34, 2989–3005. [Google Scholar] [CrossRef] [PubMed]
- Ingouff, M.; Rademacher, S.; Holec, S.; Šoljić, L.; Xin, N.; Readshaw, A.; Foo, S.H.; Lahouze, B.; Sprunck, S.; Berger, F. Zygotic Resetting of the HISTONE 3 Variant Repertoire Participates in Epigenetic Reprogramming in Arabidopsis. Curr. Biol. 2010, 20, 2137–2143. [Google Scholar] [CrossRef] [PubMed]
- Honys, D.; Twell, D. Transcriptome analysis of haploid male gametophyte development in Arabidopsis. Genome Biol. 2004, 5, R85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, G.; Panda, K.; Köhler, C.; Slotkin, R.K. Silencing in sperm cells is directed by RNA movement from the surrounding nurse cell. Nat. Plants 2016, 2, 16030. [Google Scholar] [CrossRef] [PubMed]
- Vaillant, I.; Schubert, I.; Tourmente, S.; Mathieu, O. MOM1 mediates DNA-methylation-independent silencing of repetitive sequences in Arabidopsis. EMBO Rep. 2006, 7, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Liu, B. Tapetum-Dependent Male Meiosis Progression in Plants: Increasing Evidence Emerges. Front. Plant Sci. 2020, 10, 1667. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.J.; Spielman, M.; Dickinson, H.G. Stamen structure and function. Plant Cell 2004, 16, S46–S60. [Google Scholar] [CrossRef]
- Solís, M.-T.; Chakrabarti, N.; Corredor, E.; Cortés-Eslava, J.; Rodríguez-Serrano, M.; Biggiogera, M.; Risueño, M.C.; Testillano, P.S. Epigenetic Changes Accompany Developmental Programmed Cell Death in Tapetum Cells. Plant Cell Physiol. 2013, 55, 16–29. [Google Scholar] [CrossRef]
- Axtell, M.J. Classification and Comparison of Small RNAs from Plants. Annu. Rev. Plant Biol. 2013, 64, 137–159. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Li, P.; Zhai, J.; Zhou, M.; Ma, L.; Liu, B.; Jeong, D.H.; Nakano, M.; Cao, S.; Liu, C.; et al. Roles of DCL4 and DCL3b in rice phased small RNA biogenesis. Plant J. Cell Mol. Biol. 2012, 69, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Zhang, H.; Arikit, S.; Huang, K.; Nan, G.L.; Walbot, V.; Meyers, B.C. Spatiotemporally dynamic, cell-type-dependent premeiotic and meiotic phasiRNAs in maize anthers. Proc. Natl. Acad. Sci. USA 2015, 112, 3146–3151. [Google Scholar] [CrossRef]
- Fei, Q.; Yang, L.; Liang, W.; Zhang, D.; Meyers, B.C. Dynamic changes of small RNAs in rice spikelet development reveal specialized reproductive phasiRNA pathways. J. Exp. Bot. 2016, 67, 6037–6049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Ma, X.; Wang, C.; Li, Q.; Meyers, B.C.; Springer, N.M.; Walbot, V. CHH DNA methylation increases at 24-PHAS loci depend on 24-nt phased small interfering RNAs in maize meiotic anthers. New Phytol. 2021, 229, 2984–2997. [Google Scholar] [CrossRef] [PubMed]
- Dukowic-Schulze, S.; Sundararajan, A.; Ramaraj, T.; Kianian, S.; Pawlowski, W.P.; Mudge, J.; Chen, C. Novel Meiotic miRNAs and Indications for a Role of PhasiRNAs in Meiosis. Front. Plant Sci. 2016, 7, 762. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, K.; Morohoshi, A.; Nakano, M.; Eiguchi, M.; Miyao, A.; Hirochika, H.; Kurata, N. A germ cell specific gene of the ARGONAUTE family is essential for the progression of premeiotic mitosis and meiosis during sporogenesis in rice. Plant Cell 2007, 19, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Komiya, R.; Ohyanagi, H.; Niihama, M.; Watanabe, T.; Nakano, M.; Kurata, N.; Nonomura, K. Rice germline-specific Argonaute MEL1 protein binds to phasiRNAs generated from more than 700 lincRNAs. Plant J. Cell Mol. Biol. 2014, 78, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Liu, H.; Tsuda, K.; Fukai, E.; Tanaka, K.; Sasaki, T.; Nonomura, K.I. EAT1 transcription factor, a non-cell-autonomous regulator of pollen production, activates meiotic small RNA biogenesis in rice anther tapetum. PLoS Genet. 2018, 14, e1007238. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Luo, D.; Tang, M.; Cao, S.; Pan, J.; Zhang, W.; Hu, Y.; Yue, J.; Huang, Z.; Li, R.; et al. Integrated Methylome and Transcriptome Analysis Provides Insights into the DNA Methylation Underlying the Mechanism of Cytoplasmic Male Sterility in Kenaf (Hibiscus cannabinus L.). Int. J. Mol. Sci. 2022, 23, 6864. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ding, X.; Wang, X.; He, T.; Zhang, H.; Yang, L.; Wang, T.; Chen, L.; Gai, J.; Yang, S. Genome-wide comparative analysis of DNA methylation between soybean cytoplasmic male-sterile line NJCMS5A and its maintainer NJCMS5B. BMC Genom. 2017, 18, 596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, X.; Wu, Z.; An, H.; Yi, B.; Wen, J.; Ma, C.; Shen, J.; Fu, T.; Tu, J. Genome-Wide DNA Methylation Comparison between Brassica napus Genic Male Sterile Line and Restorer Line. Int. J. Mol. Sci. 2018, 19, 2689. [Google Scholar] [CrossRef]
- Hu, J.; Chen, X.; Zhang, H.; Ding, Y. Genome-wide analysis of DNA methylation in photoperiod- and thermo-sensitive male sterile rice Peiai 64S. BMC Genom. 2015, 16, 102. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Min, L.; Wang, M.; Wang, C.; Zhao, Y.; Li, Y.; Fang, Q.; Wu, Y.; Xie, S.; Ding, Y.; et al. Disrupted Genome Methylation in Response to High Temperature Has Distinct Affects on Microspore Abortion and Anther Indehiscence. Plant Cell 2018, 30, 1387–1403. [Google Scholar] [CrossRef] [Green Version]
- Cartagena, J.A.; Matsunaga, S.; Seki, M.; Kurihara, D.; Yokoyama, M.; Shinozaki, K.; Fujimoto, S.; Azumi, Y.; Uchiyama, S.; Fukui, K. The Arabidopsis SDG4 contributes to the regulation of pollen tube growth by methylation of histone H3 lysines 4 and 36 in mature pollen. Dev. Biol. 2008, 315, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Thorstensen, T.; Grini, P.E.; Mercy, I.S.; Alm, V.; Erdal, S.; Aasland, R.; Aalen, R.B. The Arabidopsis SET-domain protein ASHR3 is involved in stamen development and interacts with the bHLH transcription factor ABORTED MICROSPORES (AMS). Plant Mol. Biol. 2008, 66, 47–59. [Google Scholar] [CrossRef]
- Pinon, V.; Yao, X.; Dong, A.; Shen, W.H. SDG2-Mediated H3K4me3 Is Crucial for Chromatin Condensation and Mitotic Division during Male Gametogenesis in Arabidopsis. Plant Physiol. 2017, 174, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Verdone, L.; Caserta, M.; Di Mauro, E. Role of histone acetylation in the control of gene expression. Biochem. Cell Biol. 2005, 83, 344–353. [Google Scholar] [CrossRef]
- Sarnowski, T.J.; Ríos, G.; Jásik, J.; Swiezewski, S.; Kaczanowski, S.; Li, Y.; Kwiatkowska, A.; Pawlikowska, K.; Koźbiał, M.; Koźbiał, P.; et al. SWI3 subunits of putative SWI/SNF chromatin-remodeling complexes play distinct roles during Arabidopsis development. Plant Cell 2005, 17, 2454–2472. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, C.; Bergounioux, C.; Domenichini, S.; Delarue, M.; Zhou, D.X. Arabidopsis histone acetyltransferase AtGCN5 regulates the floral meristem activity through the WUSCHEL/AGAMOUS pathway. J. Biol. Chem. 2003, 278, 28246–28251. [Google Scholar] [CrossRef] [PubMed]
- Vlachonasios, K.E.; Thomashow, M.F.; Triezenberg, S.J. Disruption mutations of ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis growth, development, and gene expression. Plant Cell 2003, 15, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Genau, A.C.; Li, Z.; Renzaglia, K.S.; Fernandez Pozo, N.; Nogué, F.; Haas, F.B.; Wilhelmsson, P.K.I.; Ullrich, K.K.; Schreiber, M.; Meyberg, R.; et al. HAG1 and SWI3A/B control of male germ line development in P. patens suggests conservation of epigenetic reproductive control across land plants. Plant Reprod. 2021, 34, 149–173. [Google Scholar] [CrossRef]
- Cao, H.; Li, X.; Wang, Z.; Ding, M.; Sun, Y.; Dong, F.; Chen, F.; Liu, L.a.; Doughty, J.; Li, Y.; et al. Histone H2B Monoubiquitination Mediated by HISTONE MONOUBIQUITINATION1 and HISTONE MONOUBIQUITINATION2 Is Involved in Anther Development by Regulating Tapetum Degradation-Related Genes in Rice. Plant Physiol. 2015, 168, 1389–1405. [Google Scholar] [CrossRef]
- Ravi, M.; Shibata, F.; Ramahi, J.S.; Nagaki, K.; Chen, C.; Murata, M.; Chan, S.W. Meiosis-specific loading of the centromere-specific histone CENH3 in Arabidopsis thaliana. PLoS Genet. 2011, 7, e1002121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, L.Y.; Lin, J.; Xu, H.W.; Chen, C.X.; Chen, J.Y.; Zhang, J.; Zhang, J.; Li, Y.X.; Ye, C.; Zhang, H.; et al. Intragenic heterochromatin-mediated alternative polyadenylation modulates miRNA and pollen development in rice. New Phytol. 2021, 232, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Tang, Y.; Liu, L.; Xu, Y.; Wang, T. The methyl-CpG-binding domain family member PEM1 is essential for Ubisch body formation and pollen exine development in rice. Plant J. Cell Mol. Biol. 2022, 111, 1283–1295. [Google Scholar] [CrossRef]
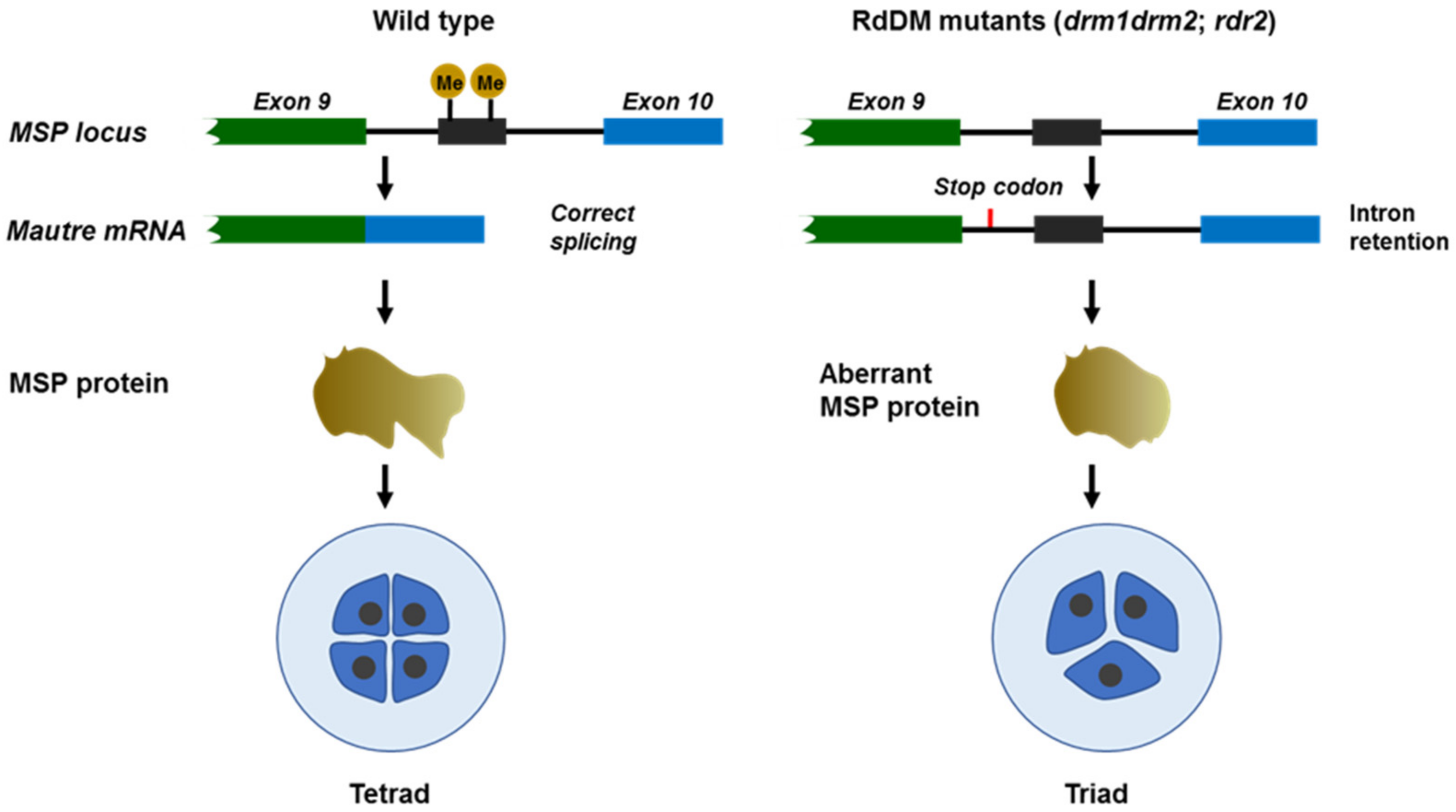
- Wang, Z.; Butel, N.; Santos-González, J.; Borges, F.; Yi, J.; Martienssen, R.A.; Martinez, G.; Köhler, C. Polymerase IV Plays a Crucial Role in Pollen Development in Capsella. Plant Cell 2020, 32, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zheng, K.; Zeng, L.; Xu, D.; Zhu, T.; Yin, Y.; Zhan, H.; Wu, Y.; Yang, D.L. Reinforcement of CHH methylation through RNA-directed DNA methylation ensures sexual reproduction in rice. Plant Physiol. 2022, 188, 1189–1209. [Google Scholar] [CrossRef]
- Oliver, C.; Annacondia, M.L.; Wang, Z.; Jullien, P.E.; Slotkin, R.K.; Köhler, C.; Martinez, G. The miRNome function transitions from regulating developmental genes to transposable elements during pollen maturation. Plant Cell 2022, 34, 784–801. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, Y.; Pan, L.; Zhao, Y.; Huang, W.; Jin, W. Male sterile 28 encodes an ARGONAUTE family protein essential for male fertility in maize. Chromosome Res. 2021, 29, 189–201. [Google Scholar] [CrossRef]
- Zheng, S.; Li, J.; Ma, L.; Wang, H.; Zhou, H.; Ni, E.; Jiang, D.; Liu, Z.; Zhuang, C. OsAGO2 controls ROS production and the initiation of tapetal PCD by epigenetically regulating OsHXK1 expression in rice anthers. Proc. Natl. Acad. Sci. USA 2019, 116, 7549–7558. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-C.; He, R.-R.; Lian, J.-P.; Zhou, Y.-F.; Zhang, F.; Li, Q.-F.; Yu, Y.; Feng, Y.-Z.; Yang, Y.-W.; Lei, M.-Q.; et al. OsmiR528 regulates rice-pollen intine formation by targeting an uclacyanin to influence flavonoid metabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Peral, M.M.; Li, J.; Li, Y.; Allen, R.S.; Schnippenkoetter, W.; Ohms, S.; White, R.G.; Millar, A.A. The microRNA159-regulated GAMYB-like genes inhibit growth and promote programmed cell death in Arabidopsis. Plant Physiol. 2010, 154, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Sun, G.; Shi, C.; Sun, D. Transcriptome analysis reveals new microRNAs-mediated pathway involved in anther development in male sterile wheat. BMC Genom. 2018, 19, 333. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, T.; Eulgem, T. An alternative polyadenylation mechanism coopted to the Arabidopsis RPP7 gene through intronic retrotransposon domestication. Proc. Natl. Acad. Sci. USA 2013, 110, E3535–E3543. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.T.; Cocklin, R.; Mayampurath, A.; Lister, R.; Rowley, M.J.; Gregory, B.D.; Ecker, J.R.; Tang, H.; Pikaard, C.S. Spatial and functional relationships among Pol V-associated loci, Pol IV-dependent siRNAs, and cytosine methylation in the Arabidopsis epigenome. Genes Dev. 2012, 26, 1825–1836. [Google Scholar] [CrossRef] [PubMed]
- Meister, G. Argonaute proteins: Functional insights and emerging roles. Nat. Rev. Genet. 2013, 14, 447–459. [Google Scholar] [CrossRef]
- Mallory, A.; Vaucheret, H. Form, function, and regulation of ARGONAUTE proteins. Plant Cell 2010, 22, 3879–3889. [Google Scholar] [CrossRef]
- McCue, A.D.; Nuthikattu, S.; Reeder, S.H.; Slotkin, R.K. Gene expression and stress response mediated by the epigenetic regulation of a transposable element small RNA. PLoS Genet. 2012, 8, e1002474. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Pereira, P.A.; Slotkin, R.K.; Martienssen, R.A.; Becker, J.D. MicroRNA activity in the Arabidopsis male germline. J. Exp. Bot. 2011, 62, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Kidner, C.A.; Martienssen, R.A. The role of ARGONAUTE1 (AGO1) in meristem formation and identity. Dev. Biol. 2005, 280, 504–517. [Google Scholar] [CrossRef]
- Oliver, C.; Pradillo, M.; Jover-Gil, S.; Cuñado, N.; Ponce, M.R.; Santos, J.L. Loss of function of Arabidopsis microRNA-machinery genes impairs fertility, and has effects on homologous recombination and meiotic chromatin dynamics. Sci. Rep. 2017, 7, 9280. [Google Scholar] [CrossRef]
- Araki, S.; Le, N.T.; Koizumi, K.; Villar-Briones, A.; Nonomura, K.I.; Endo, M.; Inoue, H.; Saze, H.; Komiya, R. miR2118-dependent U-rich phasiRNA production in rice anther wall development. Nat. Commun. 2020, 11, 3115. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.Q.; Ali, I.; Tang, J.; Yang, W.C. New Insights into 5hmC DNA Modification: Generation, Distribution and Function. Front. Genet. 2017, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karanthamalai, J.; Chodon, A.; Chauhan, S.; Pandi, G. DNA N(6)-Methyladenine Modification in Plant Genomes-A Glimpse into Emerging Epigenetic Code. Plants 2020, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Sun, H.X.; Li, J.; Li, J.; Zhao, Y.; Chen, L.; Qin, C.; Chen, R.; Chen, Z.; Liu, G.; et al. The single-cell stereo-seq reveals region-specific cell subtypes and transcriptome profiling in Arabidopsis leaves. Dev. Cell 2022, 57, 1299–1310.e1294. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Guillaume-Gentil, O.; Rainer, P.Y.; Gäbelein, C.G.; Saelens, W.; Gardeux, V.; Klaeger, A.; Dainese, R.; Zachara, M.; Zambelli, T.; et al. Live-seq enables temporal transcriptomic recording of single cells. Nature 2022, 608, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Kaya-Okur, H.S.; Janssens, D.H.; Henikoff, J.G.; Ahmad, K.; Henikoff, S. Efficient low-cost chromatin profiling with CUT&Tag. Nat. Protoc. 2020, 15, 3264–3283. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Gene Product | Function | Species | References |
|---|---|---|---|---|
| AtASHR3(AtSDG4) | SET-domain protein histone methyltransferase | Histone modification | Arabidopsis thaliana | [72,73] |
| AtSDG2 | Histone methyltransferases SET DOMAIN GROUP2 | Histone modification | Arabidopsis thaliana | [74] |
| AtSWI3B/3C/3D | Component of SWITCH/SUCROSE NONFERMENTING (SWI/SNF) chromatin-remodeling complexes | Chromatin remodeling | Arabidopsis thaliana | [76] |
| AtADA2b | Histone acetyltransferase | Chromatin remodeling | Arabidopsis thaliana | [78] |
| AtGCN5 | Histone acetyltransferase | Chromatin remodeling | Arabidopsis thaliana | [78] |
| PpHAG1 | Histone acetyltransferase of the GNAT family 1 | Histone modification | Physcomitrella patens | [79] |
| PpSWI3A/B | Component of SWITCH/SUCROSE NONFERMENTING (SWI/SNF) chromatin-remodeling complexes | Chromatin remodeling | Physcomitrella patens | [79] |
| OsHUB1/2 | Histone monoubiquitination 1/2 | Histone modification | Rice | [80] |
| AtCENH3 | Histone variant | Chromatin remodeling | Arabidopsis thaliana | [81] |
| OsASI1 | Antisilencing 1, intragenic heterochromatin reader | Alternative polyadenylation regulation | Rice | [82] |
| OsPEM1 | methyl-CpG-binding domain containing protein | DNA methylation reader | Rice | [83] |
| CrNRPD1 | The largest subunit of Polymerase IV | RdDM | Capsella rubella | [84] |
| OsRDR2 | RNA-dependent RNA polymerase 2 | RdDM | Rice | [85] |
| AtDCL1 | Dicer-like protein | miRNA generation | Arabidopsis thaliana | [86] |
| AtAGO1/5 | Argonaute | Associate with miRNA | Arabidopsis thaliana | [86] |
| ZmAGO5c | Argonaute | Associate with siRNA | Maize | [87] |
| OsMEL1 | Argonaute | Associate with siRNA | Rice | [64] |
| OsAGO2 | Argonaute | Associate with siRNA | Rice | [88] |
| OsmiR528 | microRNA | Gene silencing | Rice | [89] |
| AtmiR159 | microRNA | Gene silencing | Arabidopsis thaliana | [90] |
| TamiR2275 | microRNA | Gene silencing | Wheat | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Q.; Zhang, T.; Qi, Y.; Dong, Z.; Wan, X. Epigenetic Dynamics and Regulation of Plant Male Reproduction. Int. J. Mol. Sci. 2022, 23, 10420. https://doi.org/10.3390/ijms231810420
Hou Q, Zhang T, Qi Y, Dong Z, Wan X. Epigenetic Dynamics and Regulation of Plant Male Reproduction. International Journal of Molecular Sciences. 2022; 23(18):10420. https://doi.org/10.3390/ijms231810420
Chicago/Turabian StyleHou, Quancan, Tianye Zhang, Yuchen Qi, Zhenying Dong, and Xiangyuan Wan. 2022. "Epigenetic Dynamics and Regulation of Plant Male Reproduction" International Journal of Molecular Sciences 23, no. 18: 10420. https://doi.org/10.3390/ijms231810420