
Targeting Persistent Neuroinflammation after Hypoxic-Ischemic Encephalopathy—Is Exendin-4 the Answer?

{kind=link}
{kind=link}
Abstract
:1. Introduction
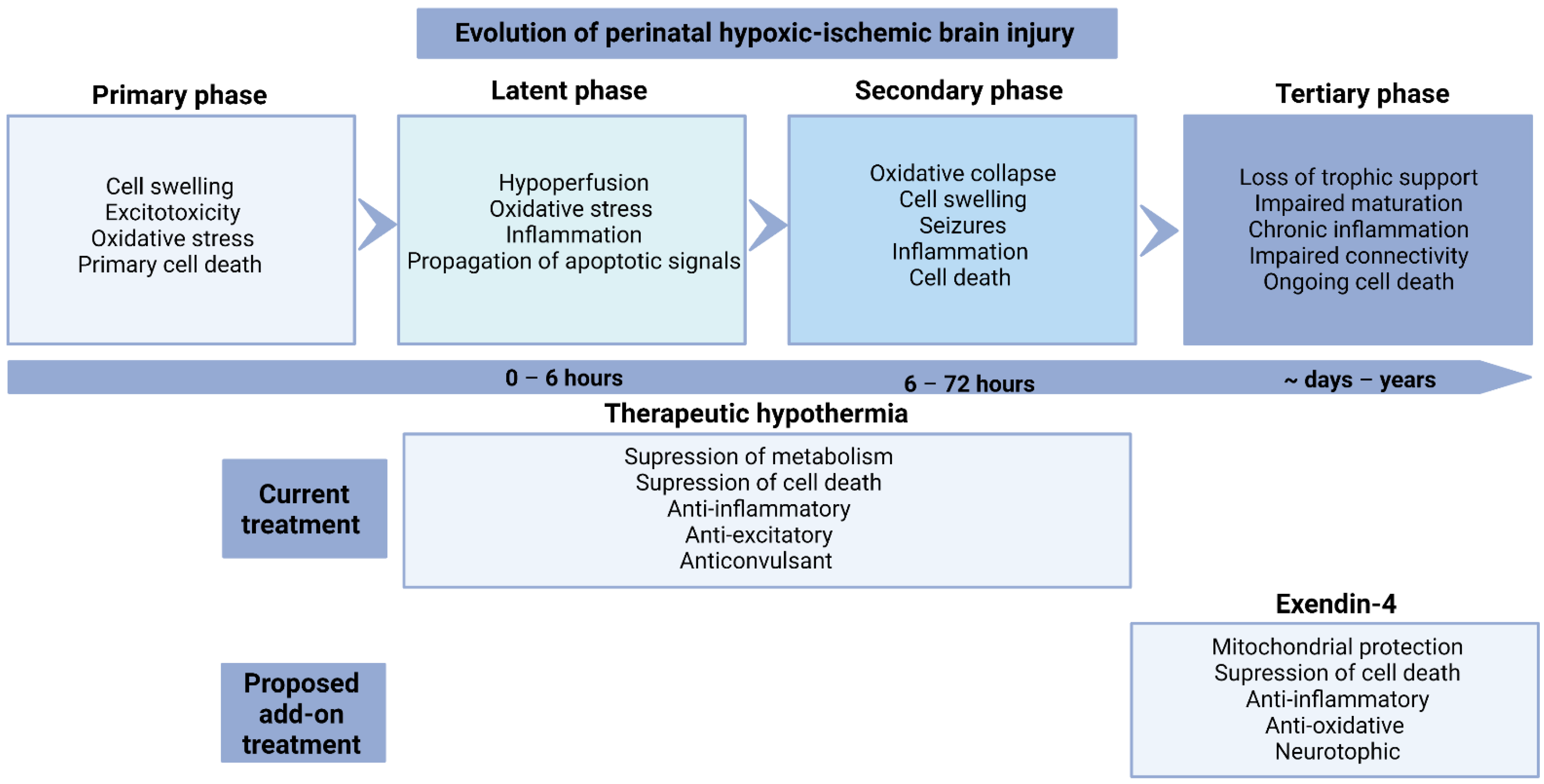
2. Evolution of Injury
3. Inflammation after HIE
3.1. Role of Microglia
3.2. Role of Astrocytes
4. Mechanisms of Therapeutic Hypothermia
Anti-Inflammatory Effects
5. Is Chronic Inflammation after Hypothermia Deleterious?
6. Exendin-4
6.1. Background
6.2. Preclinical Studies for HIE
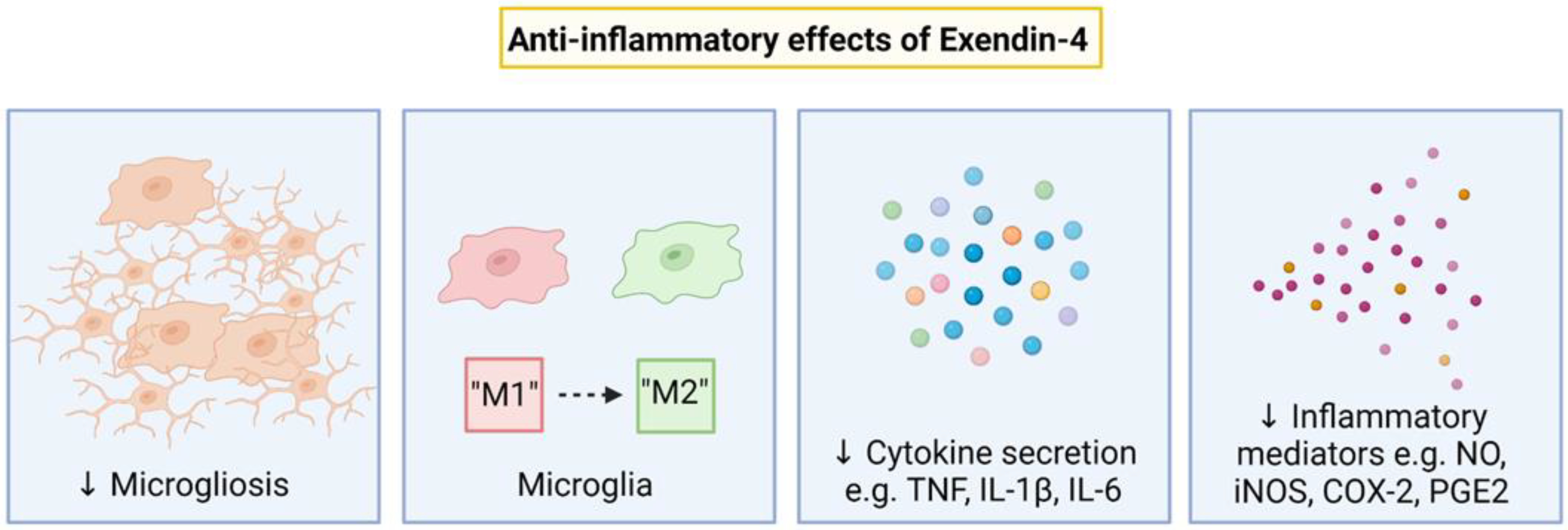
7. Anti-Inflammatory Effects of Exendin-4
Inflammatory Pathways Targetted by Exendin-4
8. Other Effects of Exendin-4
8.1. Mitochondrial Protection
8.2. Anti-Apoptotic
8.3. Anti-Oxidation
8.4. Neurotrophic
9. Timing of Exendin-4 Administration
10. Safety of Exendin-4 in Infants
11. Other GLP-1R Agonists
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischaemic encephalopathy. Cochrane Database Syst. Rev. 2013, 1, CD003311. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, G.; Laptook, A.; Shankaran, S. Therapeutic hypothermia: How can we optimize this therapy to further improve outcomes? Clin. Perinatol. 2018, 45, 241–255. [Google Scholar] [CrossRef]
- Azzopardi, D.; Strohm, B.; Marlow, N.; Brocklehurst, P.; Deierl, A.; Eddama, O.; Goodwin, J.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; et al. Effects of hypothermia for perinatal asphyxia on childhood outcomes. N. Engl. J. Med. 2014, 371, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, S.; Laptook, A.R.; Pappas, A.; McDonald, S.A.; Das, A.; Tyson, J.E.; Poindexter, B.B.; Schibler, K.; Bell, E.F.; Heyne, R.J.; et al. Effect of Depth and Duration of Cooling on Death or Disability at Age 18 Months Among Neonates with Hypoxic-Ischemic Encephalopathy: A Randomized Clinical Trial. JAMA 2017, 318, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Tonks, J.; Cloke, G.; Lee-Kelland, R.; Jary, S.; Thoresen, M.; Cowan, F.M.; Chakkarapani, E. Attention and visuo-spatial function in children without cerebral palsy who were cooled for neonatal encephalopathy: A case-control study. Brain Inj. 2019, 33, 894–898. [Google Scholar] [CrossRef]
- Lee-Kelland, R.; Jary, S.; Tonks, J.; Cowan, F.M.; Thoresen, M.; Chakkarapani, E. School-age outcomes of children without cerebral palsy cooled for neonatal hypoxic-ischaemic encephalopathy in 2008–2010. Arch. Dis. Child.-Fetal Neonatal Ed. 2019, 105, 8–13. [Google Scholar] [CrossRef]
- Spencer, A.P.C.; Brooks, J.C.W.; Masuda, N.; Byrne, H.; Lee-Kelland, R.; Jary, S.; Thoresen, M.; Goodfellow, M.; Cowan, F.M.; Chakkarapani, E. Motor function and white matter connectivity in children cooled for neonatal encephalopathy. NeuroImage Clin. 2021, 32, 102872. [Google Scholar] [CrossRef]
- Thayyil, S.; Pant, S.; Montaldo, P.; Shukla, D.; Oliveira, V.; Ivain, P.; Bassett, P.; Swamy, R.; Mendoza, J.; Moreno-Morales, M.; et al. Hypothermia for moderate or severe neonatal encephalopathy in low-income and middle-income countries (HELIX): A randomised controlled trial in India, Sri Lanka, and Bangladesh. Lancet Glob Health 2021, 9, e1273–e1285. [Google Scholar] [CrossRef]
- Lee, A.C.; Kozuki, N.; Blencowe, H.; Vos, T.; Bahalim, A.; Darmstadt, G.L.; Niermeyer, S.; Ellis, M.; Robertson, N.J.; Cousens, S.; et al. Intrapartum-related neonatal encephalopathy incidence and impairment at regional and global levels for 2010 with trends from 1990. Pediatr. Res. 2013, 74, 50–72. [Google Scholar] [CrossRef]
- Draghi, V.; Wassink, G.; Zhou, K.Q.; Bennet, L.; Gunn, A.J.; Davidson, J.O. Differential effects of slow rewarming after cerebral hypothermia on white matter recovery after global cerebral ischemia in near-term fetal sheep. Sci. Rep. 2019, 9, 10142. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.Q.; Bennet, L.; Wassink, G.; McDouall, A.; Curtis, M.A.; Highet, B.; Stevenson, T.J.; Gunn, A.J.; Davidson, J.O. Persistent cortical and white matter inflammation after therapeutic hypothermia for ischemia in near-term fetal sheep. J. Neuroinflammation. 2022, 19, 139. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ferreira, E.; Poupon, L.; Zelco, A.; Leverin, A.L.; Nair, S.; Jonsdotter, A.; Carlsson, Y.; Thornton, C.; Hagberg, H.; Rahim, A.A. Neuroprotective exendin-4 enhances hypothermia therapy in a model of hypoxic-ischaemic encephalopathy. Brain 2018, 141, 2925–2942. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.Q.; Davidson, J.O.; Bennet, L.; Gunn, A.J. Combination treatments with therapeutic hypothermia for hypoxic-ischemic neuroprotection. Dev. Med. Child Neurol. 2020, 62, 1131–1137. [Google Scholar] [CrossRef]
- Wassink, G.; Davidson, J.O.; Lear, C.A.; Juul, S.E.; Northington, F.; Bennet, L.; Gunn, A.J. A working model for hypothermic neuroprotection. J. Physiol. 2018, 596, 5641–5654. [Google Scholar] [CrossRef]
- Iwata, O.; Iwata, S.; Bainbridge, A.; De Vita, E.; Matsuishi, T.; Cady, E.B.; Robertson, N.J. Supra- and sub-baseline phosphocreatine recovery in developing brain after transient hypoxia-ischaemia: Relation to baseline energetics, insult severity and outcome. Brain 2008, 131, 2220–2226. [Google Scholar] [CrossRef]
- Davidson, J.O.; Dean, J.M.; Fraser, M.; Wassink, G.; Andelius, T.C.; Dhillon, S.K.; Bennet, L.; Gunn, A.J. Perinatal brain injury: Mechanisms and therapeutic approaches. Front. Biosci. 2018, 23, 2204–2226. [Google Scholar] [CrossRef]
- Davidson, J.O.; Wassink, G.; Yuill, C.A.; Zhang, F.G.; Bennet, L.; Gunn, A.J. How long is too long for cerebral cooling after ischemia in fetal sheep? J. Cereb. Blood Flow Metab. 2015, 35, 751–758. [Google Scholar] [CrossRef]
- Bennet, L.; Roelfsema, V.; Pathipati, P.; Quaedackers, J.; Gunn, A.J. Relationship between evolving epileptiform activity and delayed loss of mitochondrial activity after asphyxia measured by near-infrared spectroscopy in preterm fetal sheep. J. Physiol. 2006, 572, 141–154. [Google Scholar] [CrossRef]
- Baburamani, A.A.; Lo, C.; Castillo-Melendez, M.; Walker, D.W. Morphological evaluation of the cerebral blood vessels in the late gestation fetal sheep following hypoxia in utero. Microvasc. Res. 2013, 85, 1–9. [Google Scholar] [CrossRef]
- Northington, F.J.; Zelaya, M.E.; O’Riordan, D.P.; Blomgren, K.; Flock, D.L.; Hagberg, H.; Ferriero, D.M.; Martin, L.J. Failure to complete apoptosis following neonatal hypoxia-ischemia manifests as “continuum” phenotype of cell death and occurs with multiple manifestations of mitochondrial dysfunction in rodent forebrain. Neuroscience 2007, 149, 822–833. [Google Scholar] [CrossRef] [Green Version]
- Northington, F.J.; Chavez-Valdez, R.; Martin, L.J. Neuronal cell death in neonatal hypoxia-ischemia. Ann. Neurol. 2011, 69, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Ness, J.K.; Romanko, M.J.; Rothstein, R.P.; Wood, T.L.; Levison, S.W. Perinatal hypoxia-ischemia induces apoptotic and excitotoxic death of periventricular white matter oligodendrocyte progenitors. Dev. Neurosci. 2001, 23, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Romanko, M.J.; Rothstein, R.P.; Levison, S.W. Neural stem cells in the subventricular zone are resilient to hypoxia/ischemia whereas progenitors are vulnerable. J. Cereb. Blood Flow Metab. 2004, 24, 814–825. [Google Scholar] [CrossRef]
- Ziemka-Nalecz, M.; Jaworska, J.; Zalewska, T. Insights into the Neuroinflammatory Responses After Neonatal Hypoxia-Ischemia. J. Neuropathol. Exp. Neurol. 2017, 76, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Foster-Barber, A.; Dickens, B.; Ferriero, D.M. Human perinatal asphyxia: Correlation of neonatal cytokines with MRI and outcome. Dev. Neurosci. 2001, 23, 213–218. [Google Scholar] [CrossRef]
- Bartha, A.I.; Foster-Barber, A.; Miller, S.P.; Vigneron, D.B.; Glidden, D.V.; Barkovich, A.J.; Ferriero, D.M. Neonatal encephalopathy: Association of cytokines with MR spectroscopy and outcome. Pediatr. Res. 2004, 56, 960–966. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chang, Y.C.; Wang, S.T.; Lee, T.Y.; Lin, C.F.; Huang, C.C. Altered inflammatory responses in preterm children with cerebral palsy. Ann. Neurol. 2010, 68, 204–212. [Google Scholar] [CrossRef]
- Barakat, R.; Redzic, Z. Differential cytokine expression by brain microglia/macrophages in primary culture after oxygen glucose deprivation and their protective effects on astrocytes during anoxia. Fluids Barriers CNS 2015, 12, 6. [Google Scholar] [CrossRef]
- Fumagalli, S.; Perego, C.; Pischiutta, F.; Zanier, E.R.; De Simoni, M.G. The ischemic environment drives microglia and macrophage function. Front. Neurol. 2015, 6, 81. [Google Scholar] [CrossRef]
- Czeh, M.; Gressens, P.; Kaindl, A.M. The yin and yang of microglia. Dev. Neurosci. 2011, 33, 199–209. [Google Scholar] [CrossRef]
- Denker, S.P.; Ji, S.; Dingman, A.; Lee, S.Y.; Derugin, N.; Wendland, M.F.; Vexler, Z.S. Macrophages are comprised of resident brain microglia not infiltrating peripheral monocytes acutely after neonatal stroke. J. Neurochem. 2007, 100, 893–904. [Google Scholar] [CrossRef] [PubMed]
- McRae, A.; Gilland, E.; Bona, E.; Hagberg, H. Microglia activation after neonatal hypoxic-ischemia. Brain Res. Dev. Brain Res. 1995, 84, 245–252. [Google Scholar] [CrossRef]
- Fleiss, B.; Van Steenwinckel, J.; Bokobza, C.; Shearer, I.K.; Ross-Munro, E.; Gressens, P. Microglia-mediated neurodegeneration in perinatal brain injuries. Biomolecules 2021, 11, 99. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, J.; Sivakumar, V.; Ling, E.A.; Kaur, C. Amoeboid microglia in the periventricular white matter induce oligodendrocyte damage through expression of proinflammatory cytokines via MAP kinase signaling pathway in hypoxic neonatal rats. Brain Pathol. 2008, 18, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Sun, D.; Hu, Y.; Cui, D. White matter injury in the neonatal hypoxic-ischemic brain and potential therapies targeting microglia. J. Neurosci. Res. 2021, 99, 991–1008. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Baburamani, A.A.; Supramaniam, V.G.; Hagberg, H.; Mallard, C. Microglia toxicity in preterm brain injury. Reprod. Toxicol. 2014, 48, 106–112. [Google Scholar] [CrossRef]
- Hu, X. Microglia/macrophage polarization: Fantasy or evidence of functional diversity? J. Cereb. Blood Flow Metab. 2020, 40, S134–S136. [Google Scholar] [CrossRef] [PubMed]
- Bonestroo, H.J.; Nijboer, C.H.; van Velthoven, C.T.; Kavelaars, A.; Hack, C.E.; van Bel, F.; Heijnen, C.J. Cerebral and hepatic inflammatory response after neonatal hypoxia-ischemia in newborn rats. Dev. Neurosci. 2013, 35, 197–211. [Google Scholar] [CrossRef]
- Jaworska, J.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The potential neuroprotective role of a histone deacetylase inhibitor, sodium butyrate, after neonatal hypoxia-ischemia. J. Neuroinflammation 2017, 14, 34. [Google Scholar] [CrossRef] [Green Version]
- Hellstrom, E.N.; Smith, P.L.; Fleiss, B.; Nair, S.; Svedin, P.; Wang, W.; Bostrom, M.; Gressens, P.; Hagberg, H.; Brown, K.L.; et al. Temporal characterization of microglia/macrophage phenotypes in a mouse model of neonatal hypoxic-ischemic brain injury. Front. Cell Neurosci 2016, 10, 286. [Google Scholar] [CrossRef]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflammation 2014, 11, 98. [Google Scholar] [CrossRef]
- Ferrazzano, P.; Chanana, V.; Uluc, K.; Fidan, E.; Akture, E.; Kintner, D.B.; Cengiz, P.; Sun, D. Age-dependent microglial activation in immature brains after hypoxia- ischemia. CNS Neurol. Disord. Drug Targets 2013, 12, 338–349. [Google Scholar] [CrossRef]
- Cikla, U.; Chanana, V.; Kintner, D.B.; Covert, L.; Dewall, T.; Waldman, A.; Rowley, P.; Cengiz, P.; Ferrazzano, P. Suppression of microglia activation after hypoxia-ischemia results in age-dependent improvements in neurologic injury. J. Neuroimmunol. 2016, 291, 18–27. [Google Scholar] [CrossRef]
- Winerdal, M.; Winerdal, M.E.; Kinn, J.; Urmaliya, V.; Winqvist, O.; Aden, U. Long lasting local and systemic inflammation after cerebral hypoxic ischemia in newborn mice. PLoS ONE 2012, 7, e36422. [Google Scholar] [CrossRef]
- Davidson, J.O.; Yuill, C.A.; Zhang, F.G.; Wassink, G.; Bennet, L.; Gunn, A.J. Extending the duration of hypothermia does not further improve white matter protection after ischemia in term-equivalent fetal sheep. Sci. Rep. 2016, 6, 25178. [Google Scholar] [CrossRef] [PubMed]
- van den Heuij, L.G.; Fraser, M.; Miller, S.L.; Jenkin, G.; Wallace, E.M.; Davidson, J.O.; Lear, C.A.; Lim, R.; Wassink, G.; Gunn, A.J.; et al. Delayed intranasal infusion of human amnion epithelial cells improves white matter maturation after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2019, 39, 223–239. [Google Scholar] [CrossRef]
- Bona, E.; Andersson, A.L.; Blomgren, K.; Gilland, E.; Puka-Sundvall, M.; Gustafson, K.; Hagberg, H. Chemokine and inflammatory cell response to hypoxia-ischemia in immature rats. Pediatr. Res. 1999, 45, 500–509. [Google Scholar] [CrossRef]
- Romero, J.; Muniz, J.; Logica Tornatore, T.; Holubiec, M.; Gonzalez, J.; Barreto, G.E.; Guelman, L.; Lillig, C.H.; Blanco, E.; Capani, F. Dual role of astrocytes in perinatal asphyxia injury and neuroprotection. Neurosci. Lett. 2014, 565, 42–46. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrocyte barriers to neurotoxic inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [Green Version]
- McKeon, R.J.; Schreiber, R.C.; Rudge, J.S.; Silver, J. Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J. Neurosci. 1991, 11, 3398–3411. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, F.; Quintana, F.J. The Role of Astrocytes in CNS Inflammation. Trends Immunol. 2020, 41, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Odorcyk, F.K.; Nicola, F.; Duran-Carabali, L.E.; Figueiro, F.; Kolling, J.; Vizuete, A.; Konrath, E.L.; Goncalves, C.A.; Wyse, A.T.S.; Netto, C.A. Galantamine administration reduces reactive astrogliosis and upregulates the anti-oxidant enzyme catalase in rats submitted to neonatal hypoxia ischemia. Int. J. Dev. Neurosci. 2017, 62, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Wassink, G.; Davidson, J.O.; Dhillon, S.K.; Fraser, M.; Galinsky, R.; Bennet, L.; Gunn, A.J. Partial white and grey matter protection with prolonged infusion of recombinant human erythropoietin after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2017, 37, 1080–1094. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.M.; Bjorkman, S.T.; Miller, S.M.; Colditz, P.B.; Pow, D.V. Morphological changes in white matter astrocytes in response to hypoxia/ischemia in the neonatal pig. Brain Res. 2010, 1319, 164–174. [Google Scholar] [CrossRef]
- Wassink, G.; Davidson, J.O.; Dhillon, S.K.; Zhou, K.; Bennet, L.; Thoresen, M.; Gunn, A.J. Therapeutic hypothermia in neonatal hypoxic-ischemic encephalopathy. Curr. Neurol. Neurosci. Rep. 2019, 19, 2. [Google Scholar] [CrossRef]
- Seo, J.W.; Kim, J.H.; Seo, M.; Han, H.S.; Park, J.; Suk, K. Time-dependent effects of hypothermia on microglial activation and migration. J. Neuroinflammation 2012, 9, 164. [Google Scholar] [CrossRef]
- Han, H.S.; Karabiyikoglu, M.; Kelly, S.; Sobel, R.A.; Yenari, M.A. Mild hypothermia inhibits nuclear factor-kappaB translocation in experimental stroke. J. Cereb. Blood Flow Metab. 2003, 23, 589–598. [Google Scholar] [CrossRef]
- Han, H.S.; Qiao, Y.; Karabiyikoglu, M.; Giffard, R.G.; Yenari, M.A. Influence of mild hypothermia on inducible nitric oxide synthase expression and reactive nitrogen production in experimental stroke and inflammation. J. Neurosci. 2002, 22, 3921–3928. [Google Scholar] [CrossRef]
- Wang, G.J.; Deng, H.Y.; Maier, C.M.; Sun, G.H.; Yenari, M.A. Mild hypothermia reduces ICAM-1 expression, neutrophil infiltration and microglia/monocyte accumulation following experimental stroke. Neuroscience 2002, 114, 1081–1090. [Google Scholar] [CrossRef]
- Zhou, K.Q.; Draghi, V.; Lear, C.A.; Dean, J.M.; Ashton, J.L.; Hou, Y.; Bennet, L.; Gunn, A.J.; Davidson, J.O. Protection of axonal integrity with 48 or 72 h of cerebral hypothermia in near-term fetal sheep. Pediatr. Res. 2020, 88, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Köster, C.; Dzietko, M.; Sabir, H.; Serdar, M.; Felderhoff-Müser, U.; Bendix, I.; Herz, J. Hypothermia modulates myeloid cell polarization in neonatal hypoxic-ischemic brain injury. J. Neuroinflammation 2021, 18, 266. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, S.; Di Martino, E.; Mukai, T.; Tsuji, S.; Murakami, T.; Harris, R.A.; Blomgren, K.; Åden, U. Aggravated brain injury after neonatal hypoxic ischemia in microglia-depleted mice. J. Neuroinflammation 2020, 17, 111. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Wei, D.; Yi, L.; Tao, X.; Ma, Y. Expression and purification of exendin-4, a GLP-1 receptor agonist, in Escherichia coli. Protein Expr. Purif. 2005, 41, 259–265. [Google Scholar] [CrossRef]
- Yap, M.K.K.; Misuan, N. Exendin-4 from Heloderma suspectum venom: From discovery to its latest application as type II diabetes combatant. Basic Clin. Pharmacol. Toxicol. 2019, 124, 513–527. [Google Scholar] [CrossRef]
- Ai, G.; Chen, Z.; Shan, C.; Che, J.; Hou, Y.; Cheng, Y. Single- and multiple-dose pharmacokinetics of exendin-4 in rhesus monkeys. Int. J. Pharm. 2008, 353, 56–64. [Google Scholar] [CrossRef]
- Deacon, C.F.; Pridal, L.; Klarskov, L.; Olesen, M.; Holst, J.J. Glucagon-like peptide 1 undergoes differential tissue-specific metabolism in the anesthetized pig. Am. J. Physiol. 1996, 271, E458–E464. [Google Scholar] [CrossRef]
- Grant, P.; Lipscomb, D.; Quin, J. Psychological and quality of life changes in patients using GLP-1 analogues. J. Diabetes Complicat. 2011, 25, 244–246. [Google Scholar] [CrossRef]
- Jin, J.; Kang, H.M.; Jung, J.; Jeong, J.W.; Park, C. Related expressional change of HIF-1α to the neuroprotective activity of exendin-4 in transient global ischemia. Neuroreport 2014, 25, 65–70. [Google Scholar] [CrossRef]
- Briyal, S.; Gulati, K.; Gulati, A. Repeated administration of exendin-4 reduces focal cerebral ischemia-induced infarction in rats. Brain Res. 2012, 1427, 23–34. [Google Scholar] [CrossRef]
- Zhang, H.; Meng, J.; Zhou, S.; Liu, Y.; Qu, D.; Wang, L.; Li, X.; Wang, N.; Luo, X.; Ma, X. Intranasal Delivery of Exendin-4 Confers Neuroprotective Effect Against Cerebral Ischemia in Mice. AAPS J. 2016, 18, 385–394. [Google Scholar] [CrossRef]
- Garabadu, D.; Verma, J. Exendin-4 attenuates brain mitochondrial toxicity through PI3K/Akt-dependent pathway in amyloid beta (1-42)-induced cognitive deficit rats. Neurochem. Int. 2019, 128, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.J.; Chen, S.; Yang, Y.Y.; Glotfelty, E.J.; Jung, J.; Kim, H.K.; Choi, H.I.; Choi, D.S.; Hoffer, B.J.; Greig, N.H.; et al. PT320, Sustained-Release Exendin-4, Mitigates L-DOPA-Induced Dyskinesia in a Rat 6-Hydroxydopamine Model of Parkinson’s Disease. Front. Neurosci. 2020, 14, 785. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Bu, R.; Yang, Q.; Jia, J.; Li, T.; Wang, Q.; Chen, Y. Exendin-4 Protects against Hyperglycemia-Induced Cardiomyocyte Pyroptosis via the AMPK-TXNIP Pathway. J. Diabetes Res. 2019, 2019, 8905917. [Google Scholar] [CrossRef]
- Garcia-Flores, V.; Romero, R.; Miller, D.; Xu, Y.; Done, B.; Veerapaneni, C.; Leng, Y.; Arenas-Hernandez, M.; Khan, N.; Panaitescu, B.; et al. Inflammation-Induced Adverse Pregnancy and Neonatal Outcomes Can Be Improved by the Immunomodulatory Peptide Exendin-4. Front. Immunol. 2018, 9, 1291. [Google Scholar] [CrossRef]
- Teramoto, S.; Miyamoto, N.; Yatomi, K.; Tanaka, Y.; Oishi, H.; Arai, H.; Hattori, N.; Urabe, T. Exendin-4, a glucagon-like peptide-1 receptor agonist, provides neuroprotection in mice transient focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2011, 31, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Jeon, S.J.; Cho, K.S.; Moon, E.; Sapkota, A.; Jun, H.S.; Ryu, J.H.; Choi, J.W. Activation of Glucagon-Like Peptide-1 Receptor Promotes Neuroprotection in Experimental Autoimmune Encephalomyelitis by Reducing Neuroinflammatory Responses. Mol. Neurobiol. 2018, 55, 3007–3020. [Google Scholar] [CrossRef]
- Darsalia, V.; Hua, S.; Larsson, M.; Mallard, C.; Nathanson, D.; Nyström, T.; Sjöholm, Å.; Johansson, M.E.; Patrone, C. Exendin-4 reduces ischemic brain injury in normal and aged type 2 diabetic mice and promotes microglial M2 polarization. PLoS ONE 2014, 9, e103114. [Google Scholar] [CrossRef]
- Wu, H.Y.; Tang, X.Q.; Liu, H.; Mao, X.F.; Wang, Y.X. Both classic Gs-cAMP/PKA/CREB and alternative Gs-cAMP/PKA/p38β/CREB signal pathways mediate exenatide-stimulated expression of M2 microglial markers. J. Neuroimmunol. 2018, 316, 17–22. [Google Scholar] [CrossRef]
- Bułdak, Ł.; Machnik, G.; Bułdak, R.J.; Łabuzek, K.; Bołdys, A.; Belowski, D.; Basiak, M.; Okopień, B. Exenatide (a GLP-1 agonist) expresses anti-inflammatory properties in cultured human monocytes/macrophages in a protein kinase A and B/Akt manner. Pharmacol. Rep. 2016, 68, 329–337. [Google Scholar] [CrossRef]
- He, L.; Wong, C.K.; Cheung, K.K.; Yau, H.C.; Fu, A.; Zhao, H.L.; Leung, K.M.; Kong, A.P.; Wong, G.W.; Chan, P.K.; et al. Anti-inflammatory effects of exendin-4, a glucagon-like peptide-1 analog, on human peripheral lymphocytes in patients with type 2 diabetes. J. Diabetes Investig. 2013, 4, 382–392. [Google Scholar] [CrossRef]
- Kim, S.; Jeong, J.; Jung, H.S.; Kim, B.; Kim, Y.E.; Lim, D.S.; Kim, S.D.; Song, Y.S. Anti-inflammatory Effect of Glucagon Like Peptide-1 Receptor Agonist, Exendin-4, through Modulation of IB1/JIP1 Expression and JNK Signaling in Stroke. Exp. Neurobiol. 2017, 26, 227–239. [Google Scholar] [CrossRef]
- Lu, C.; Xie, T.; Guo, X.; Wu, D.; Li, S.; Li, X.; Lu, Y.; Wang, X. Glucagon-like peptide-1 receptor agonist exendin-4 mitigates lipopolysaccharide-induced inflammatory responses in RAW264.7 macrophages. Int. Immunopharmacol. 2019, 77, 105969. [Google Scholar] [CrossRef]
- An, J.; Zhou, Y.; Zhang, M.; Xie, Y.; Ke, S.; Liu, L.; Pan, X.; Chen, Z. Exenatide alleviates mitochondrial dysfunction and cognitive impairment in the 5×FAD mouse model of Alzheimer’s disease. Behav. Brain Res. 2019, 370, 111932. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jia, Z.; Li, G.; Zhao, X.; Sun, P.; Wang, J.; Fan, Z.; Lv, G. Neuroprotective effects of exendin-4 in rat model of spinal cord injury via inhibiting mitochondrial apoptotic pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 4837–4843. [Google Scholar] [PubMed]
- Tews, D.; Lehr, S.; Hartwig, S.; Osmers, A.; Paslack, W.; Eckel, J. Anti-apoptotic action of exendin-4 in INS-1 beta cells: Comparative protein pattern analysis of isolated mitochondria. Horm. Metab. Res. 2009, 41, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, Z.; Huang, G.; Hu, F.; Xiang, Y.; He, L. Exendin-4 protects mitochondria from reactive oxygen species induced apoptosis in pancreatic Beta cells. PLoS ONE 2013, 8, e76172. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, K. CREB and cAMP response element-mediated gene expression in the ischemic brain. FEBS J. 2007, 274, 3210–3217. [Google Scholar] [CrossRef] [PubMed]
- Gumuslu, E.; Mutlu, O.; Celikyurt, I.K.; Ulak, G.; Akar, F.; Erden, F.; Ertan, M. Exenatide enhances cognitive performance and upregulates neurotrophic factor gene expression levels in diabetic mice. Fundam. Clin. Pharmacol. 2016, 30, 376–384. [Google Scholar] [CrossRef]
- Tsukamoto, M.; Niimi, N.; Sango, K.; Takaku, S.; Kanazawa, Y.; Utsunomiya, K. Neurotrophic and neuroprotective properties of exendin-4 in adult rat dorsal root ganglion neurons: Involvement of insulin and RhoA. Histochem. Cell Biol. 2015, 144, 249–259. [Google Scholar] [CrossRef]
- Li, H.; Lee, C.H.; Yoo, K.Y.; Choi, J.H.; Park, O.K.; Yan, B.C.; Byun, K.; Lee, B.; Hwang, I.K.; Won, M.H. Chronic treatment of exendin-4 affects cell proliferation and neuroblast differentiation in the adult mouse hippocampal dentate gyrus. Neurosci. Lett. 2010, 486, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Montaldo, P.; Caredda, E.; Pugliese, U.; Zanfardino, A.; Delehaye, C.; Inserra, E.; Capozzi, L.; Chello, G.; Capristo, C.; Miraglia Del Giudice, E.; et al. Continuous glucose monitoring profile during therapeutic hypothermia in encephalopathic infants with unfavorable outcome. Pediatr. Res. 2020, 88, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, L.B.; Nielsen, P.F.; Huusfeldt, P.O.; Johansen, N.L.; Madsen, K.; Pedersen, F.Z.; Thøgersen, H.; Wilken, M.; Agersø, H. Potent derivatives of glucagon-like peptide-1 with pharmacokinetic properties suitable for once daily administration. J. Med. Chem. 2000, 43, 1664–1669. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.S.; Bai, J.J.; Jiang, H.; Zhu, J.J.; Fu, C.C.; He, M.Z.; Zhu, J.H.; Chen, S.Q.; Li, P.J.; Fu, X.Q.; et al. Treatment with Liraglutide Exerts Neuroprotection After Hypoxic-Ischemic Brain Injury in Neonatal Rats via the PI3K/AKT/GSK3β Pathway. Front. Cell. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef]
- Lau, J.; Bloch, P.; Schäffer, L.; Pettersson, I.; Spetzler, J.; Kofoed, J.; Madsen, K.; Knudsen, L.B.; McGuire, J.; Steensgaard, D.B.; et al. Discovery of the Once-Weekly Glucagon-Like Peptide-1 (GLP-1) Analogue Semaglutide. J. Med. Chem. 2015, 58, 7370–7380. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, K.Q.; Dhillon, S.K.; Bennet, L.; Gunn, A.J.; Davidson, J.O. Targeting Persistent Neuroinflammation after Hypoxic-Ischemic Encephalopathy—Is Exendin-4 the Answer? Int. J. Mol. Sci. 2022, 23, 10191. https://doi.org/10.3390/ijms231710191
Zhou KQ, Dhillon SK, Bennet L, Gunn AJ, Davidson JO. Targeting Persistent Neuroinflammation after Hypoxic-Ischemic Encephalopathy—Is Exendin-4 the Answer? International Journal of Molecular Sciences. 2022; 23(17):10191. https://doi.org/10.3390/ijms231710191
Chicago/Turabian StyleZhou, Kelly Q., Simerdeep K. Dhillon, Laura Bennet, Alistair J. Gunn, and Joanne O. Davidson. 2022. "Targeting Persistent Neuroinflammation after Hypoxic-Ischemic Encephalopathy—Is Exendin-4 the Answer?" International Journal of Molecular Sciences 23, no. 17: 10191. https://doi.org/10.3390/ijms231710191