
Genome-Wide Characterization of PIN Auxin Efflux Carrier Gene Family in Mikania micrantha
Abstract
:1. Introduction
2. Results
2.1. Identification of the PIN Gene Family of M. micrantha
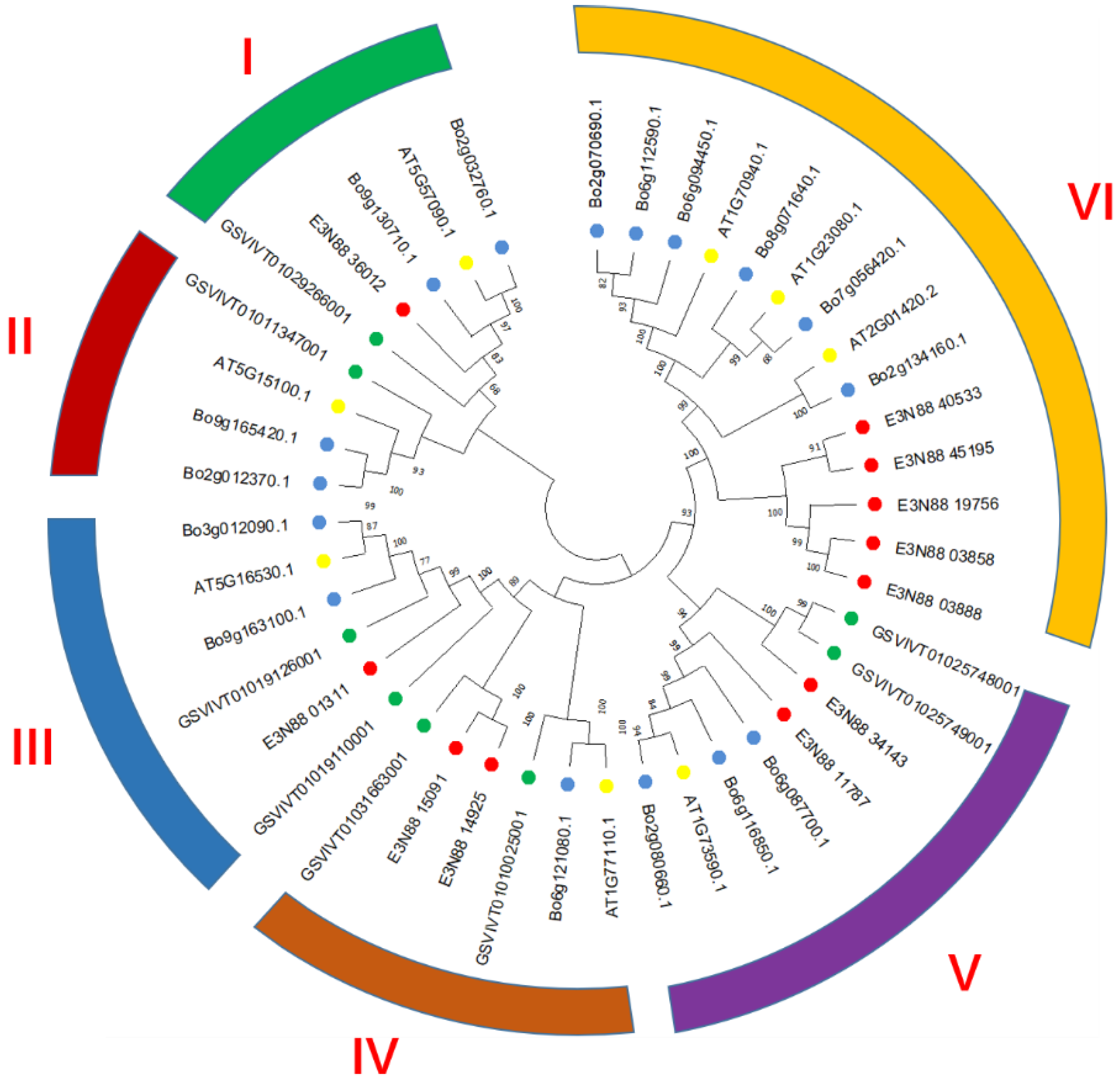
2.2. Phylogenetic Analysis of the PIN Gene Family of M. micrantha
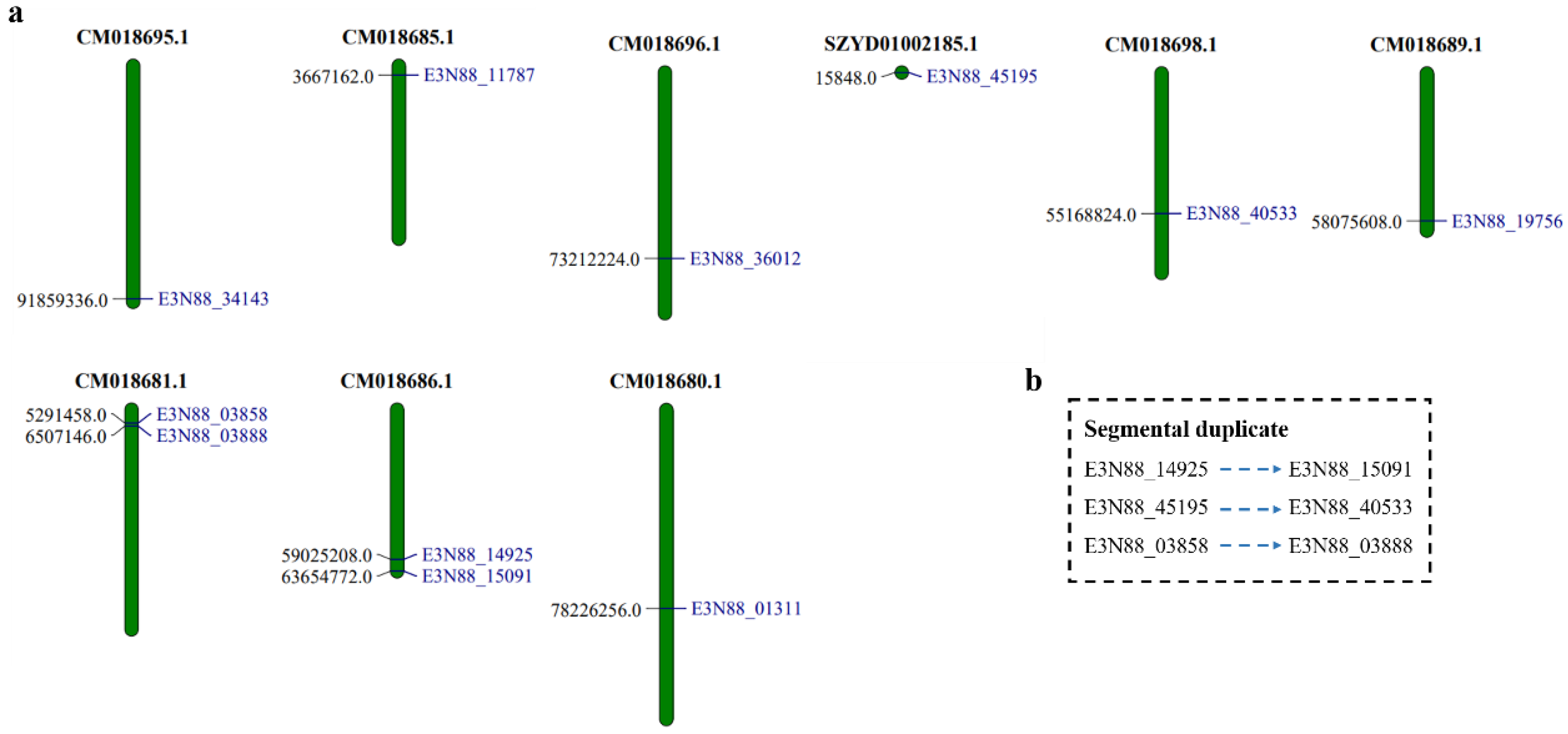
2.3. Chromosome Localization of the PIN Gene Family of M. micrantha
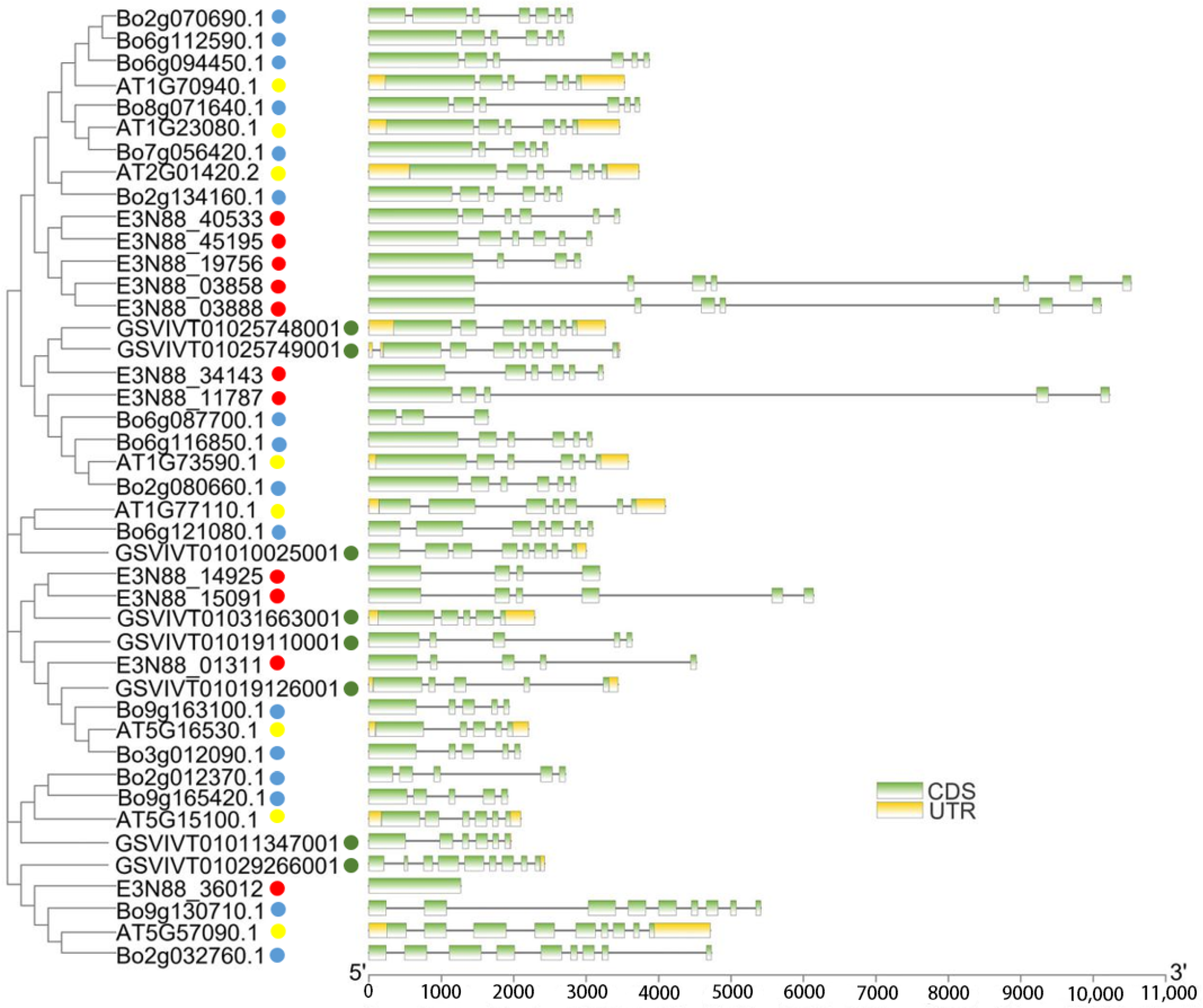
2.4. Analysis of PIN Gene Structure of M. micrantha
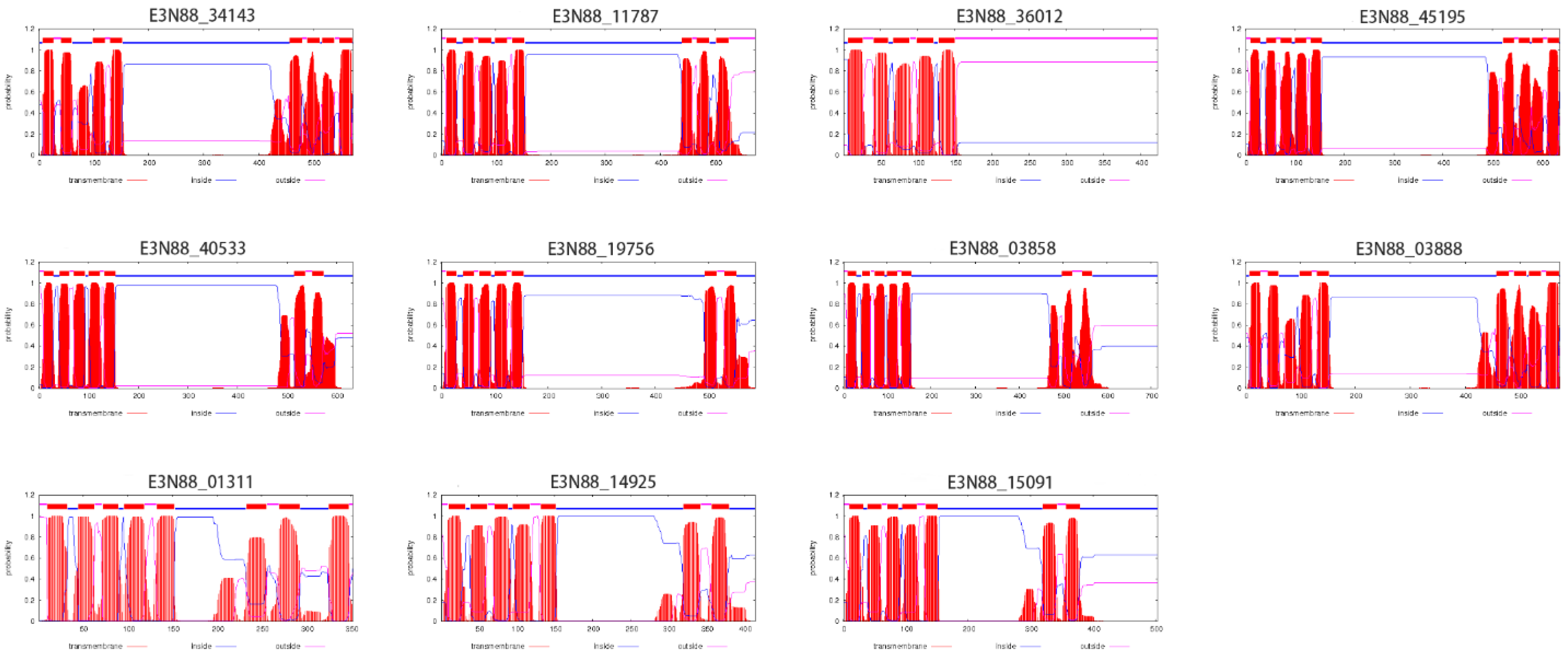
2.5. Transmembrane Region Prediction, Conserved Motifs and Multiple Sequence Alignment Analysis
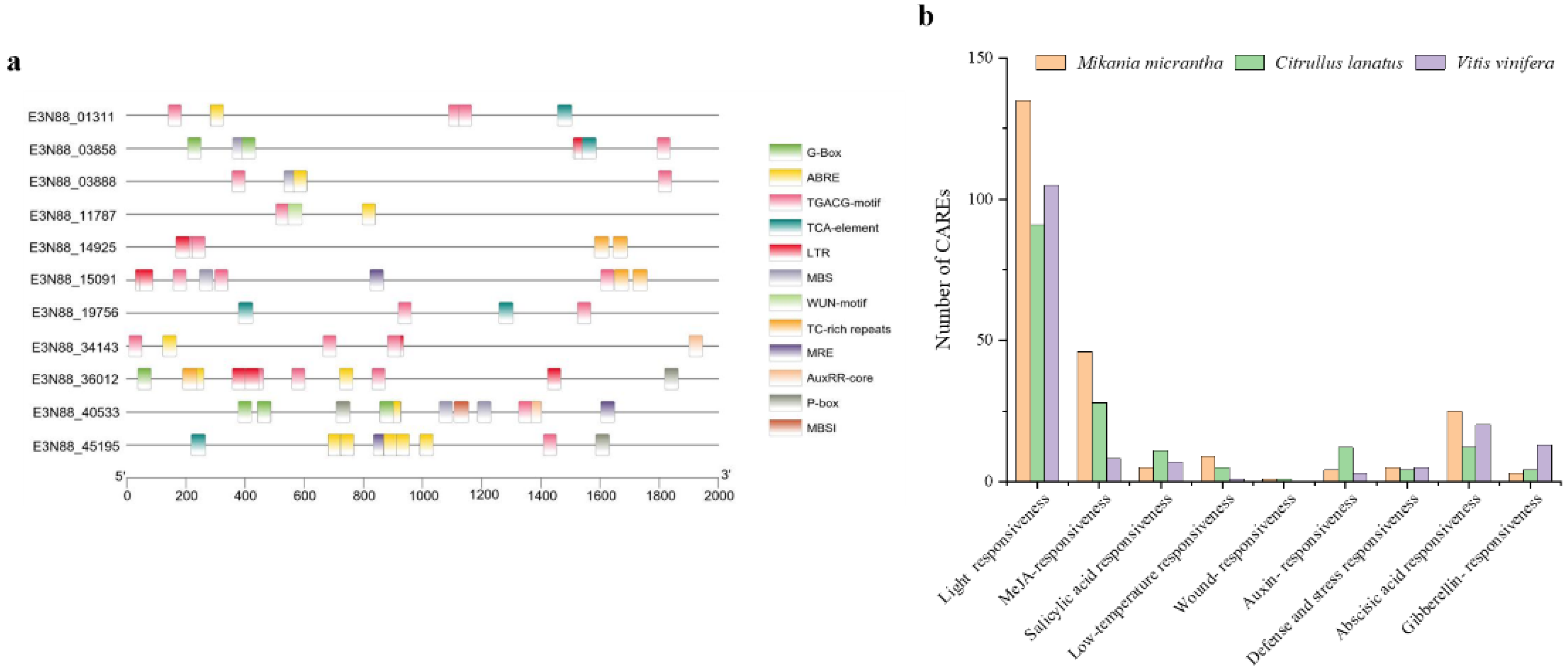
2.6. Analysis of Cis-Acting Regulatory Elements (CAREs) of the PIN Gene Family of M. micrantha
2.7. Tissue Expression Profile of the PIN Gene Family of M. micrantha
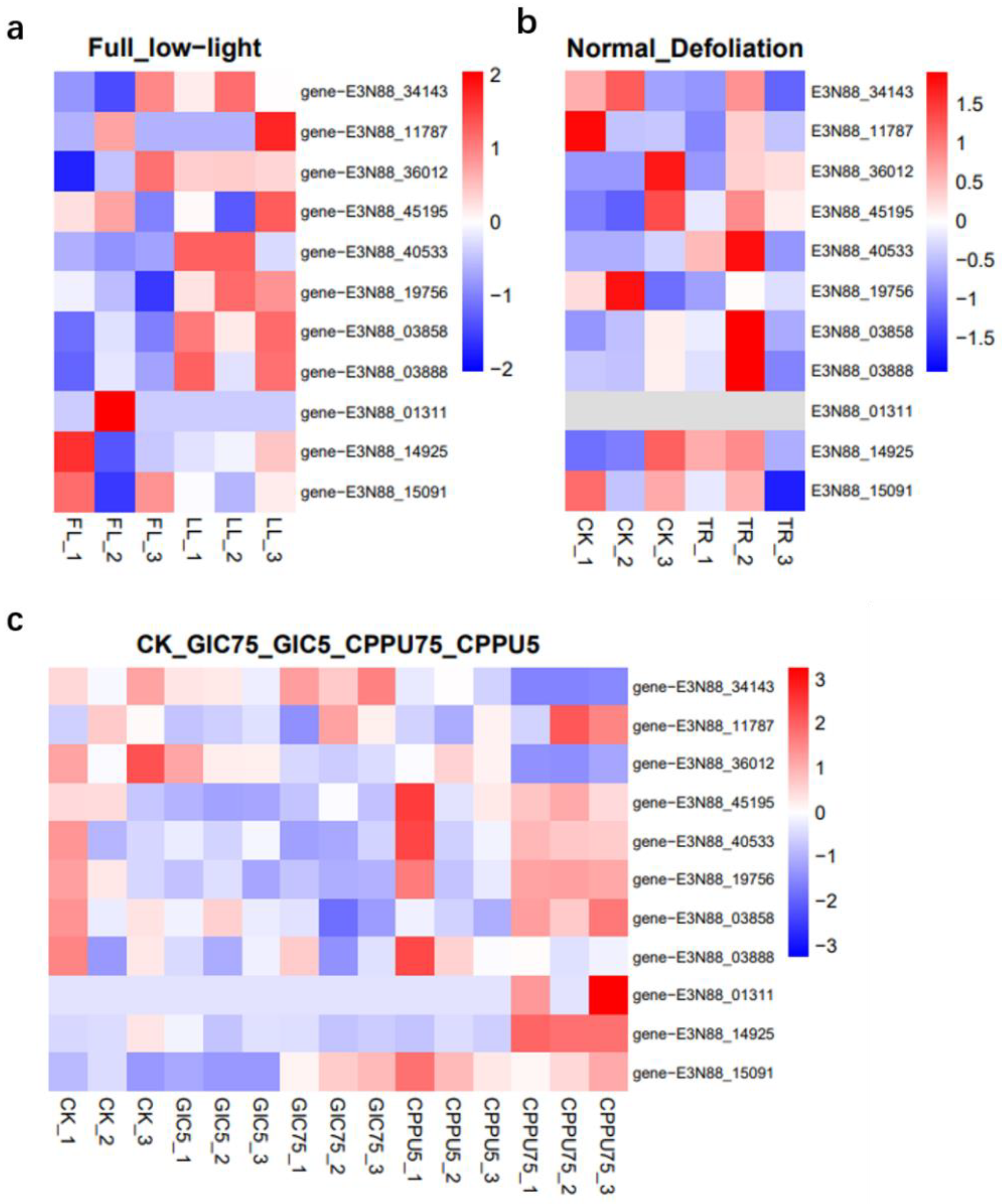
2.8. Expression Patterns Analysis of M. micrantha PINs under Abiotic Stress and Hormone Treatment
3. Discussion
3.1. Identification and Evolution of the PIN Gene Family of M. mikrantha
3.2. The Expression Profile of PIN Genes in Different Tissues Predicts Its Role in the Growth and Development of M. micrantha
3.3. Members of the PIN Gene Family of M. micrantha Have Different Response Modes to Abiotic Stress and Hormone Treatment
4. Materials and Methods
4.1. Screening and Identification of Gene Family
4.2. Chromosome Localization, Phylogenetic Analysis and Gene Duplication
4.3. Analysis of Gene Structure, Conserved Motif and Multiple Sequence Alignment
4.4. Analysis of Cis-Acting Regulatory Elements (CAREs)
4.5. Analysis of Gene Expression Pattern
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Song, L.Z.; Wang, Y.; Yang, Q.H.; Cheng, Y.F. Role of auxin in early embryogenesis of plants. Chin. Bull. Bot. 2013, 48, 371–380. [Google Scholar]
- Yu, C.L.; Dong, W.Q.; Zhan, Y.H.; Huang, Z.A.; Li, Z.M.; Kim, I.S.; Zhang, C.H. Genome-wide identification and expression analysis of ClLAX, ClPIN and ClABCB genes families in Citrullus lanatus under various abiotic stresses and grafting. BMC Genet. 2017, 18, 33. [Google Scholar] [CrossRef]
- Shibasaki, K.; Uemura, M.; Tsurumi, S.; Rahman, A. Auxin response in Arabidopsis under cold stress: Underlying molecular mechanisms. Plant Cell 2009, 21, 3823–3838. [Google Scholar] [CrossRef]
- Sharma, E.; Sharma, R.; Borah, P.; Jain, M.; Khurana, J.P. Emerging Roles of Auxin in Abiotic Stress Responses. In Elucidation of Abiotic Stress Signaling in Plants; Springer: New York, NY, USA, 2015; pp. 299–328. [Google Scholar]
- Ribba, T.; Garrido-Vargas, F.; O’Brien, J.A. Auxin-mediated responses under salt stress: From developmental regulation to biotechnological applications. J. Exp. Bot. 2020, 71, 3843–3853. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, A.P.; Liu, Y.K.; He, C.Z. The Polar Transport and Regulatory Mechanism of Auxin in Plant. J. Yunnan Agric. Univ. 2013, 28, 878–884. [Google Scholar]
- Swarup, R.; Peret, B. AUX/LAX family of auxin influx carriers-an overview. Front. Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef] [PubMed]
- Adamowski, M.; Friml, J. PIN-Dependent Auxin Transport: Action, Regulation, and Evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Murphy, A.S. The ABC of auxin transport: The role of p-glycoproteins in plant development. FEBS Lett. 2006, 580, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, Efflux-Dependent Auxin Gradients as a Common Module for Plant Organ Formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef]
- Scarpella, E.; Marcos, D.; Friml, J.; Berleth, T. Control of leaf vascular patterning by polar auxin transport. Genes Dev. 2006, 20, 1015–1027. [Google Scholar] [CrossRef]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [Green Version]
- Swarup, R.; Bennett, M. Auxin transport: The fountain of life in plants? Dev. Cell 2003, 5, 824–826. [Google Scholar] [CrossRef]
- Paponov, I.A.; Teale, W.D.; Trebar, M.; Blilou, I.; Palme, K. The PIN auxin efflux facilitators: Evolutionary and functional perspectives. Trends Plant Sci. 2005, 10, 170–177. [Google Scholar] [CrossRef]
- Roumeliotis, E.; Kloosterman, B.; Oortwijn, M.; Visser, R.G.; Bachem, C.W. The PIN family of proteins in potato and their putative role in tuberization. Front. Plant Sci. 2013, 4, 524. [Google Scholar] [CrossRef]
- Zhou, J.J.; Luo, J. The PIN-FORMED Auxin Efflux Carriers in Plants. Int. J. Mol. Sci. 2018, 19, 2759. [Google Scholar] [CrossRef] [PubMed]
- Křeček, P.; Skůpa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zažímalová, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef]
- Simon, S.; Skůpa, P.; Viaene, T.; Zwiewka, M.; Tejos, R.; Klíma, P.; Čarná, M.; Rolčík, J.; Rycke, R.D.; Moreno, I.; et al. PIN6 auxin transporter at endoplasmic reticulum and plasma membrane mediates auxin homeostasis and organogenesis in Arabidopsis. New Phytol. 2016, 211, 65–74. [Google Scholar] [CrossRef]
- Ditengou, F.A.; Gomes, D.; Nziengui, H.; Kochersperger, P.; Lasok, H.; Medeiros, V.; Paponov, I.A.; Nagy, S.K.; Nádai, T.V.; Mészáros, T.; et al. Characterization of auxin transporter PIN6 plasma membrane targeting reveals a function for PIN6 in plant bolting. New Phytol. 2018, 217, 1610–1624. [Google Scholar] [CrossRef]
- Bennett, T.; Brockington, S.F.; Rothfels, C.; Graham, S.W.; Stevenson, D.; Kutchan, T.; Rolf, M.; Thomas, P.; Wong, G.K.S.; Leyser, O.; et al. Paralogous radiations of PIN proteins with multiple origins of noncanonical PIN structure. Mol. Biol. Evol. 2014, 31, 2042–2060. [Google Scholar] [CrossRef]
- Friml, J.; Benková, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jürgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef]
- Reinhardt, D.; Pesce, E.R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Traas, J.; Friml, J.; Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Ottenschläger, I.; Wolff, P.; Wolverton, C.; Bhalerao, R.P.; Sandberg, G.; Ishikawa, H.; Evans, M.; Palme, K. Gravity-regulated differential auxin transport from columella to lateral root cap cells. Proc. Natl. Acad. Sci. USA 2003, 100, 2987–2991. [Google Scholar] [CrossRef]
- Blakeslee, J.J.; Bandyopadhyay, A.; Peer, W.A.; Makam, S.N.; Murphy, A.S. Relocalization of the PIN1 Auxin Efflux Facilitator Plays a Role in Phototropic Responses. Plant Physiol. 2004, 134, 28–31. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, C.J.; Wang, W.J. Bioinformatics Analysis of PIN-formed Family in Oryza sativa. J. Anhui Agric. Sci. 2012, 40, 13238–13242. [Google Scholar]
- Dal Bosco, C.; Dovzhenko, A.; Palme, K. Intracellular auxin transport in pollen: PIN8, PIN5 and PILS5. Plant Signal. Behav. 2012, 7, 1504–1505. [Google Scholar] [CrossRef]
- Zheng, J.L.; Luo, Q.; Li, Z.T.; Men, S.Z. Bioinformatics Analysis of the Auxin Efflux Carrier PIN Proteins of Ricinus communis L. J. Liaocheng Univ. (Nat. Sci.) 2021, 34, 88–99. [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds: Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Huang, F.F.; Peng, S.L. Intraspecific competitive ability declines towards the edge of the expanding range of the invasive vine Mikania micrantha. Oecologia 2016, 181, 115–123. [Google Scholar] [CrossRef]
- Hu, L.; Li, M.G. Climbing capacity of the invasive vine Mikania micrantha Kunth on vertical artificial poles. Biol. Invasions 2013, 16, 295–302. [Google Scholar] [CrossRef]
- Waterhouse, D.F. Biological Control of Weeds: Southeast Asian Prospects; ACIAR: Canberra, Australia, 1994. [Google Scholar]
- Zhang, L.Y.; Ye, W.H.; Cao, H.L.; Feng, H.L. Mikania micrantha H.B.K. in China—An overview. Weed Res. 2004, 44, 42–49. [Google Scholar] [CrossRef]
- Yang, T.; Davies, P.J.; Reid, J.B. Genetic Dissection of the Relative Roles of Auxin and Gibberellin in the Regulation of Stem Elongation in Intact Light-Grown Peas. Plant Physiol. 1996, 110, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yan, J.; Li, W.H.; Yin, L.; Li, P.; Yu, H.; Xing, L.S.; Cai, M.L.; Wang, H.C.; Zhao, M.X.; et al. Mikania micrantha genome provides insights into the molecular mechanism of rapid growth. Nat. Commun. 2020, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Wang, Y.T.; Zheng, Y.P.; Cai, M.L.; Peng, C.L.; Li, W.H. Physiological and transcriptomic responses of Mikania micrantha stem to shading yield novel insights into its invasiveness. Biol. Invasions 2021, 23, 2927–2943. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, Y.; Ge, D.; Wang, Z.; Song, W.; Gu, R.; Che, G.; Cheng, Z.H.; Liu, R.Y.; Zhang, X.L. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar]
- Wang, H.; Jiang, W.W.; Guo, Y.C.; Si, J.P.; Chen, D.H. Genome-wide Identification and Expression Analysis of PIN Gene Family in Dendrobium catenatum. J. Agric. Biotechnol. 2021, 29, 1649–1664. [Google Scholar]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Zhao, N.; Ze, S.Z.; Liu, N.Y.; Hu, L.R.; Ji, M.; Li, Q.; Yang, B. Exogenous phytohormone application and transcriptome analysis of Mikania micrantha provides insights for a potential control strategy. Genomics 2021, 113, 964–975. [Google Scholar] [CrossRef]
- Michniewicz, M.; Brewer, P.B.; Friml, J. Polar Auxin Transport and Asymmetric Auxin Distribution. Arab. Book 2007, 5, e108. [Google Scholar]
- Habets, M.E.J.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377. [Google Scholar] [CrossRef]
- Kumar, M.; Kherawat, B.S.; Dey, P.; Saha, D.; Singh, A.; Bhatia, S.K.; Ghodake, G.S.; Kadam, A.A.; Kim, H.-U.; Manorama; et al. Genome-Wide Identification and Characterization of PIN-FORMED (PIN) Gene Family Reveals Role in Developmental and Various Stress Conditions in Triticum aestivum L. Int. J. Mol. Sci. 2021, 22, 7396. [Google Scholar] [CrossRef]
- Yang, C.H.; Wang, D.D.; Zhang, C.; Kong, N.N.; Ma, H.L.; Chen, Q. Comparative Analysis of the PIN Auxin Transporter Gene Family in Different Plant Species: A Focus on Structural and Expression Profiling of PINs in Solanum tuberosum. Int. J. Mol. Sci. 2019, 20, 3270. [Google Scholar] [CrossRef]
- Cao, J.; Li, X.; Lv, Y.Q.; Ding, L.N. Comparative analysis of the phytocyanin gene family in 10 plant species: A focus on Zea mays. Front. Plant Sci. 2015, 6, 515. [Google Scholar] [CrossRef]
- Hu, L.; Li, M.G.; Wei, P.P. Salt tolerance of the invasive vine Mikania micrantha Kunth. Ecol. Environ. Sci. 2014, 23, 7–15. [Google Scholar]
- Zhang, Q.L.; Zhai, J.J.; Chen, G.X.; Lin, W.; Peng, C.L. The Changing Distribution of Anthocyanin in Mikania micrantha Leaves as an Adaption to Low-Temperature Environments. Plants 2019, 8, 456. [Google Scholar] [CrossRef]
- Yue, M.F.; Yu, H.X.; Li, W.H.; Yin, A.G.; Cui, Y.; Tian, X.S. Flooding with shallow water promotes the invasiveness of Mikania micrantha. Ecol. Evol. 2019, 9, 9177–9184. [Google Scholar] [CrossRef]
- Deng, X. Morphological and physiological plasticity responding to different light environments of the invasive plant, Mikania micrantha H.B. Kunth. Ecol. Environ. Sci. 2010, 19, 1170–1175. [Google Scholar]
- Liang, H.L.; Zheng, Y.P.; Jiang, Z.Y.; Yue, M.F.; Li, W.H. Stem Elongation Characteristics of Mikania micrantha and Its Physiological Basis under Low Light Condition. J. Trop. Subtrop. Bot. 2022, 30, 70–78. [Google Scholar]
- Hu, L.F.; Wang, P.K.; Long, X.F.; Wu, W.H.; Zhang, J.J.; Pan, Y.; Cheng, Y.L.; Shi, J.S.; Chen, J.H. The PIN gene family in relic plant L. chinense: Genome-wide identification and gene expression profiling in different organizations and abiotic stress responses. Plant Physiol. Biochem. 2021, 162, 634–646. [Google Scholar] [CrossRef]
- Shang, H.G.; Zhang, N.; Xie, Z.L.; Deng, S.Y.; Yi, L.C.; Huang, X. Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus). Agriculture 2021, 11, 447. [Google Scholar] [CrossRef]
- Wang, Y.; Chai, C.; Valliyodan, B.; Maupin, C.; Annen, B.; Nguyen, H.T. Genome-wide analysis and expression profiling of the PIN auxin transporter gene family in soybean (Glycine max). BMC Genom. 2015, 16, 951. [Google Scholar] [CrossRef] [Green Version]
- Zwiewka, M.; Bilanovičová, V.; Seifu, Y.W.; Nodzyński, T. The Nuts and Bolts of PIN Auxin Efflux Carriers. Front. Plant Sci. 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- Galweiler, L.; Guan, C.; Muller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, J.S.; Wu, L.L.; Shao, Y.L.; Wu, Y.R.; Mao, C.Z. Functional Divergence of PIN1 Paralogous Genes in Rice. Plant Cell Physiol. 2019, 60, 2720–2732. [Google Scholar] [CrossRef] [PubMed]
- Mravec, J.; Skůpa, P.; Bailly, A.; Hoyerová, K.; Křeček, P.; Bielach, A.; Petrášek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.D.; et al. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef]
- Müller, A.; Guan, C.; Gälweiler, L.; Tänzler, P.; Huijser, P.; Marchant, A.; Parry, G.; Bennett, M.; Wisman, E.; Palme, K. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 1998, 17, 6903–6911. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Wang, B.S.; Li, M.G.; Zan, Q.J.; Wang, Y.J. The Branching Pattern and Biomass of Mikania micrantha Shoot Modules in Acacia confusa Community and Miscanthus sinensis Community. Chin. J. Plant Ecol. 2002, 26, 346. [Google Scholar]
- Rakusová, H.; Abbas, M.; Han, H.B.; Song, S.Y.; Robert, H.S.; Friml, J. Termination of Shoot Gravitropic Responses by Auxin Feedback on PIN3 Polarity. Curr. Biol. 2016, 26, 3026–3032. [Google Scholar] [CrossRef]
- Ruiz Rosquete, M.; Waidmann, S.; Kleine-Vehn, J. PIN7 Auxin Carrier Has a Preferential Role in Terminating Radial Root Expansion in Arabidopsis thaliana. Int. J. Mol. Sci. 2018, 19, 1238. [Google Scholar] [CrossRef]
- Pattison, R.J.; Catalá, C. Evaluating auxin distribution in tomato (Solanum lycopersicum) through an analysis of the PIN and AUX/LAX gene families. Plant J. 2012, 70, 585–598. [Google Scholar] [CrossRef]
- Xie, X.D.; Qin, G.Y.; Si, P.; Luo, Z.P.; Gao, J.P.; Chen, X.; Zhang, J.F.; Wei, P.; Xia, Q.Y.; Lin, F.C.; et al. Analysis of Nicotiana tabacum PIN genes identifies NtPIN4 as a key regulator of axillary bud growth. Physiol. Plant. 2017, 160, 222–239. [Google Scholar] [CrossRef]
- Waldie, T.; Leyser, O. Cytokinin targets auxin transport to promote shoot branching. Plant Physiol. 2018, 177, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.G.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar demand, not auxin, is the initial regulator of apical dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, L.; Wang, H.F.; Gu, Z.Q.; Liu, Y.F.; Wang, M.M.; Wang, M.; Xu, Y.T.; Shi, Q.B.; Li, G.; et al. MtPIN1 and MtPIN3 Play Dual Roles in Regulation of Shade Avoidance Response under Different Environments in Medicago truncatula. Int. J. Mol. Sci. 2020, 21, 8742. [Google Scholar] [CrossRef]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.; Peeters, A.J.; Pierik, R. Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc. Natl. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef]
- Voorips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gu, Z.L.; Cavalcanti, A.; Chen, F.C.; Bouman, P.; Li, W.H. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef]
- Ozyigit, I.I.; Filiz, E.; Vatansever, R.; Kurtoglu, K.Y.; Koc, I.; Öztürk, M.X.; Anjum, N.A. Identification and comparative analysis of H2O2-scavenging enzymes (ascorbate peroxidase and glutathione peroxidase) in selected plants employing bioinformatics approaches. Front. Plant Sci. 2016, 7, 301. [Google Scholar] [CrossRef]
- Wang, D.P.; Zhang, Y.B.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, X.X.; Rui, H.Y.; Zhang, F.Q.; Hu, Z.B.; Xia, Y.; Shen, Z.G. Overexpression of a functional Vicia sativa PCS1 homolog increases cadmium tolerance and phytochelatins synthesis in Arabidopsis. Front. Plant Sci. 2018, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Ginestet, C. ggplot2: Elegant graphics for data analysis. J. R. Stat. Soc. Ser. A-Stat. Soc. 2011, 174, 245. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Chromosome Location (bp) | No.of Extrons | Deducted Polypeptid | GRAVY | No. of Transmembrane | Subcellular Localization | ||
|---|---|---|---|---|---|---|---|---|
| Length (aa) | MI wt (Da) | pI | ||||||
| E3N88_34143 | CM018695.1: 91859336-91862569 (+) | 6 | 572 | 61,745.42 | 8.71 | 0.255 | 8 | Plasma membrane |
| E3N88_11787 | CM018685.1: 3667162-3677387 (+) | 5 | 573 | 62,411.30 | 9.34 | 0.154 | 8 | Plasma membrane |
| E3N88_36012 | CM018696.1: 73212228-73213496 (+) | 1 | 422 | 45,883.24 | 9.12 | −0.093 | 5 | Plasma membrane |
| E3N88_45195 | SZYD01002185.1: 15848-18924 (−) | 6 | 636 | 69,032.71 | 6.43 | 0.164 | 9 | Plasma membrane |
| E3N88_40533 | CM018698.1: 55168829-55172292 (−) | 6 | 633 | 69,241.85 | 6.07 | 0.111 | 7 | Plasma membrane |
| E3N88_19756 | CM018689.1: 58075607-58078526 (+) | 4 | 587 | 63,755.81 | 7.75 | 0.164 | 7 | Plasma membrane |
| E3N88_03858 | CM018681.1: 5291458-5301981 (−) | 7 | 715 | 77,840.88 | 8.01 | 0.040 | 7 | Plasma membrane |
| E3N88_03888 | CM018681.1: 6507146-6517256 (+) | 7 | 715 | 77,912.95 | 7.64 | 0.049 | 7 | Plasma membrane |
| E3N88_01311 | CM018680.1: 78226254-78230774 (+) | 5 | 352 | 38,726.75 | 6.51 | 0.725 | 8 | Vacular membrane |
| E3N88_14925 | CM018686.1: 59025212-59028402 (−) | 4 | 414 | 45,963.12 | 7.89 | 0.353 | 7 | Plasma membrane |
| E3N88_15091 | CM018686.1: 63654766-63660902 (+) | 6 | 503 | 56,134.80 | 9.10 | 0.122 | 7 | Plasma membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Cai, M.; Chen, M.; Ke, W.; Pan, Y.; Huang, J.; Zhang, J.; Peng, C. Genome-Wide Characterization of PIN Auxin Efflux Carrier Gene Family in Mikania micrantha. Int. J. Mol. Sci. 2022, 23, 10183. https://doi.org/10.3390/ijms231710183
Chen L, Cai M, Chen M, Ke W, Pan Y, Huang J, Zhang J, Peng C. Genome-Wide Characterization of PIN Auxin Efflux Carrier Gene Family in Mikania micrantha. International Journal of Molecular Sciences. 2022; 23(17):10183. https://doi.org/10.3390/ijms231710183
Chicago/Turabian StyleChen, Lihua, Minling Cai, Minghao Chen, Weiqian Ke, Yanru Pan, Jundong Huang, Junjie Zhang, and Changlian Peng. 2022. "Genome-Wide Characterization of PIN Auxin Efflux Carrier Gene Family in Mikania micrantha" International Journal of Molecular Sciences 23, no. 17: 10183. https://doi.org/10.3390/ijms231710183