
Antioxidant and Antibacterial Effects of Potential Probiotics Isolated from Korean Fermented Foods
 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
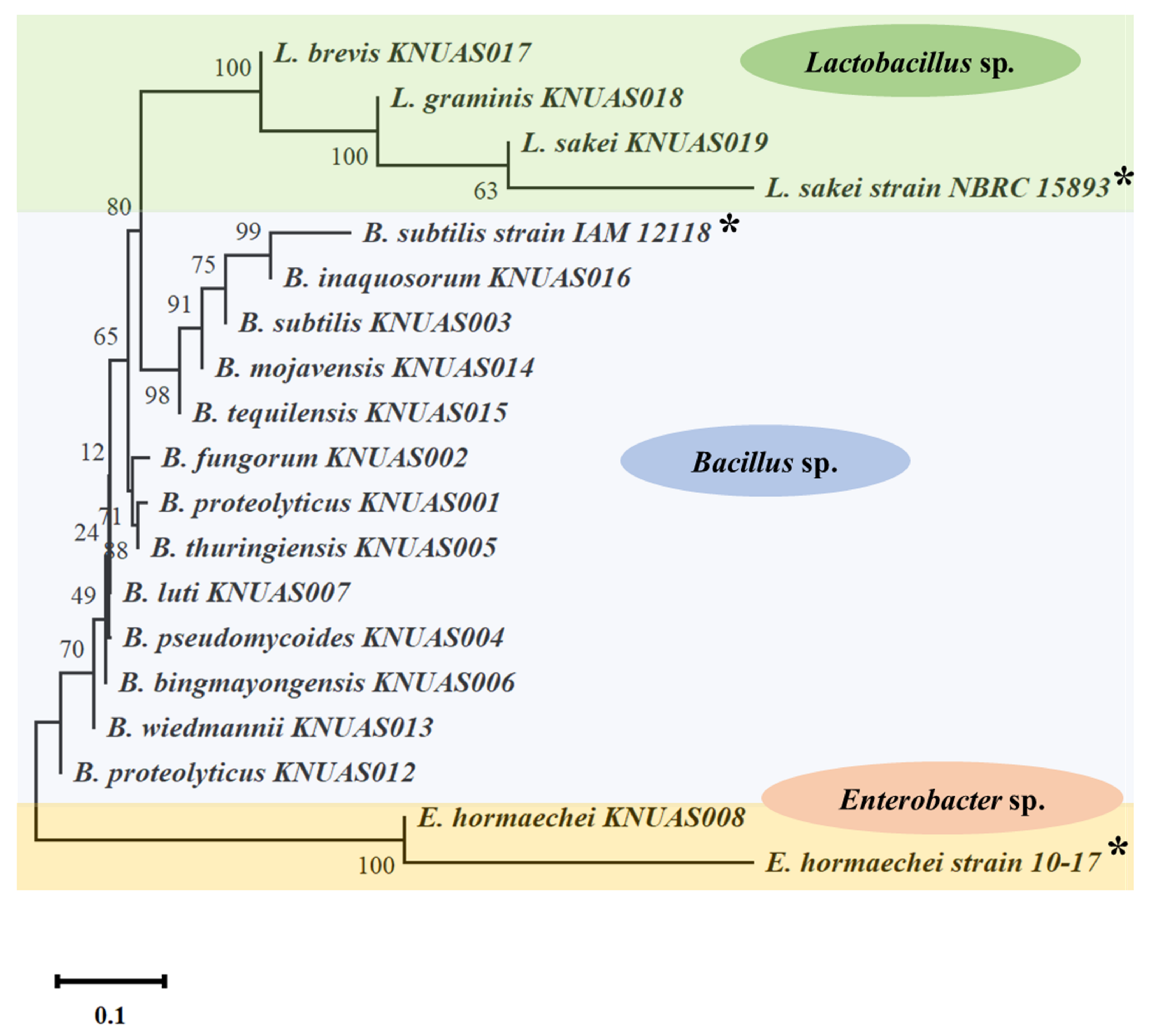
2.1. Identification of Probiotic Bacterial Strains
2.2. Resistance to Biological Barriers
2.2.1. Tolerance in Gastric and Intestinal Fluids
2.2.2. Autoaggregation, Coaggregation, and Hydrophobicity Properties of Bacterial Strains
2.3. Safety Assessment
2.3.1. Hemolytic Property
2.3.2. Antibiotic Susceptibility
2.4. Characterization of Probiotic Potential
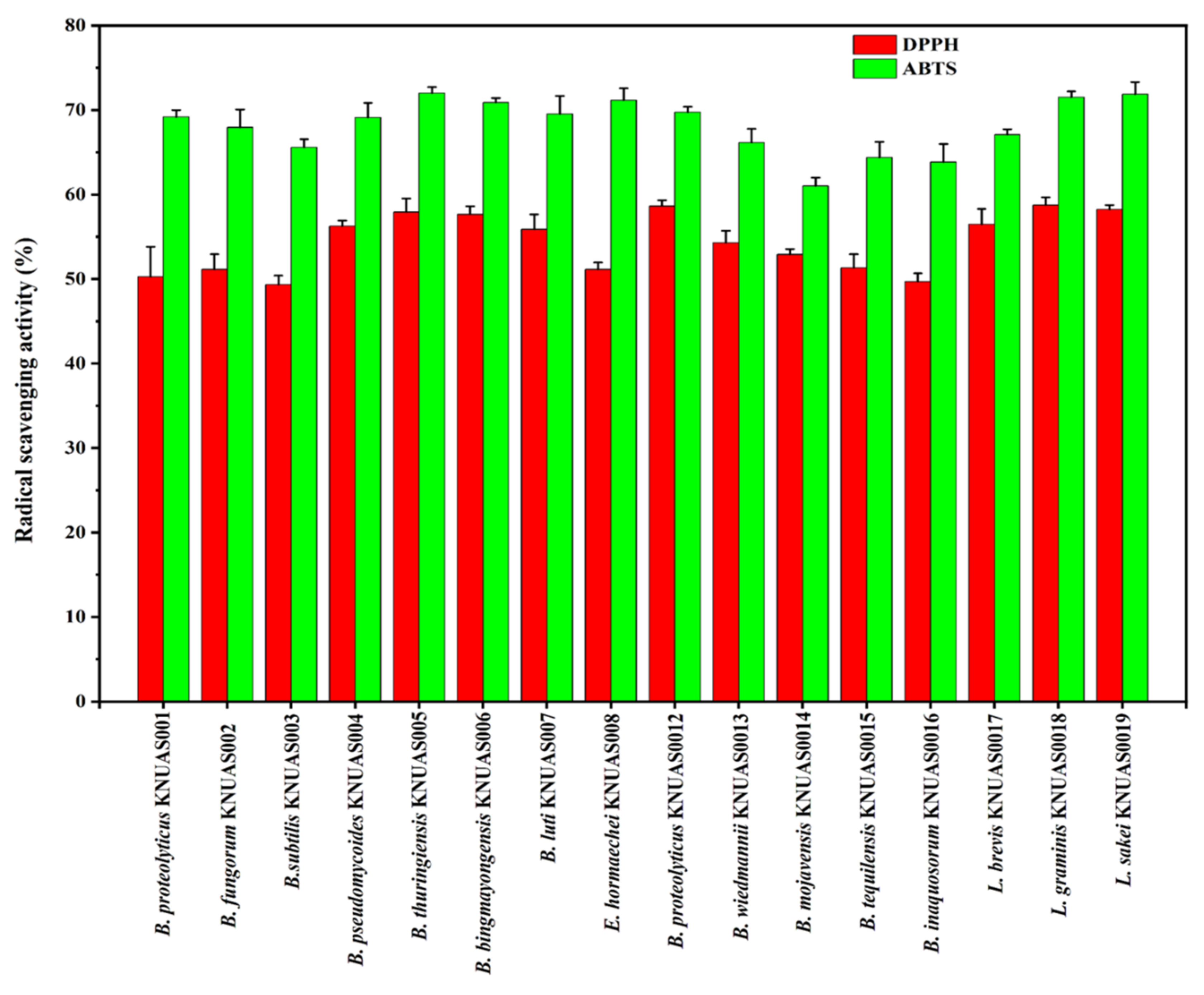
2.4.1. Antioxidant Activity
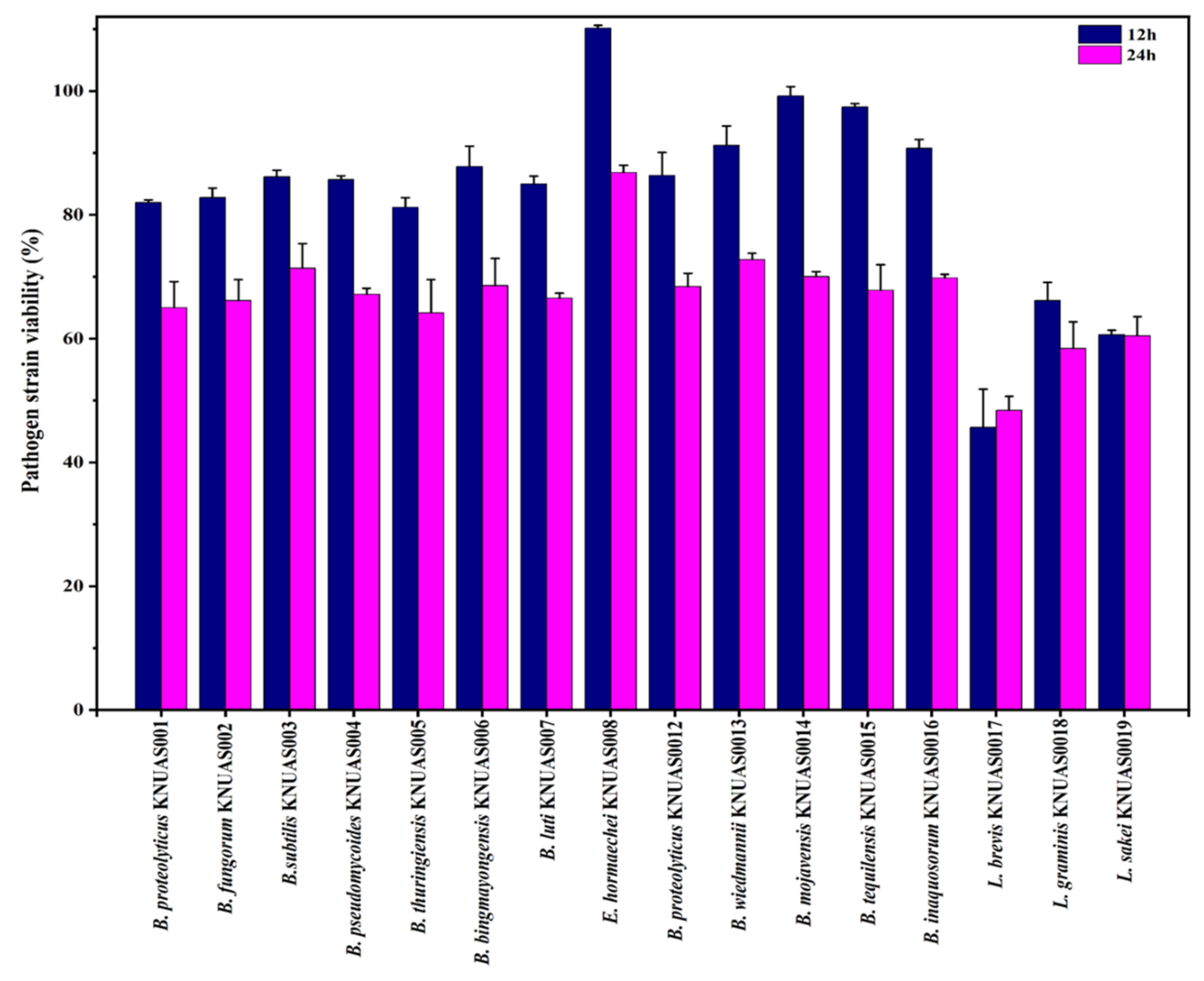
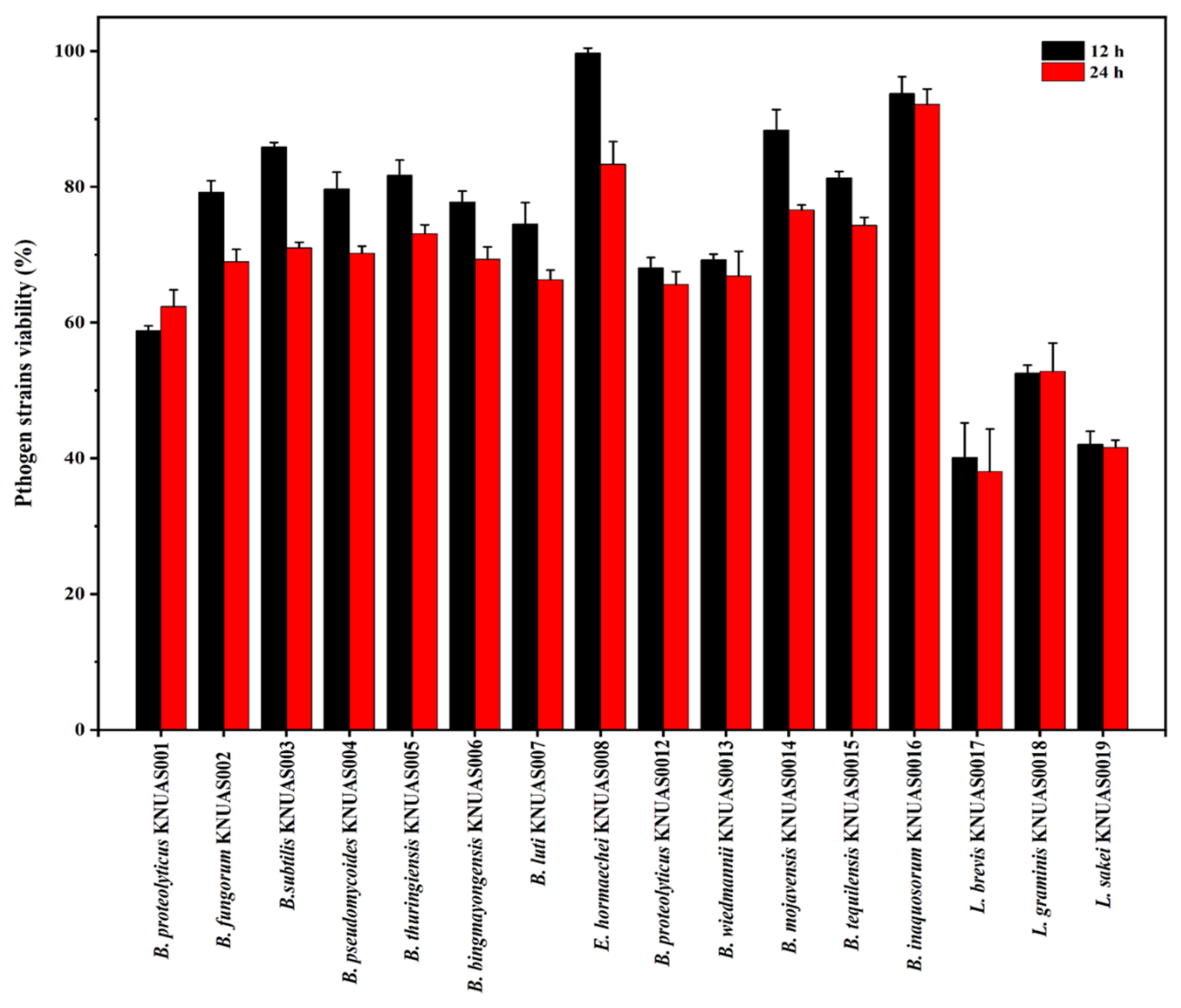
2.4.2. Antibacterial Properties
3. Materials and Methods
3.1. Isolation of Probiotic Bacterial Strains and Culture Condition
3.2. Identification of Bacterial Strains from Fermented Korean Foods
3.3. Resistance to Biological Barriers
3.3.1. Tolerance in Gastric Juice and Intestinal Fluids
3.3.2. Autoaggregation and Coaggregation Ability
3.3.3. Hydrophobicity
3.4. Safety Assessment
3.4.1. Hemolytic Property
3.4.2. Antibiotic Susceptibility
3.5. Characterization of Probiotic Potential
3.5.1. Antioxidant Activity
3.5.2. Preparation of Culture Free Supernatant
3.5.3. Antibacterial Properties
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and Host Nutrition across Plant and Animal Kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.B.; Hsiao, E.Y. Microbiomes as Sources of Emergent Host Phenotypes. Science 2019, 365, 1405–1409. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3, 4. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, K.; Wu, Y.; Yang, Y.; Tso, P.; Wu, Z. Interactions between Intestinal Microbiota and Host Immune Response in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 942. [Google Scholar] [CrossRef]
- Valitutti, F.; Cucchiara, S.; Fasano, A. Celiac Disease and the Microbiome. Nutrients 2019, 11, 2403. [Google Scholar] [CrossRef]
- Maeda, Y.; Takeda, K. Host-Microbiota Interactions in Rheumatoid Arthritis. Exp. Mol. Med. 2019, 51, 1–6. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut Microbiome Dysbiosis and Immunometabolism: New Frontiers for Treatment of Metabolic Diseases. Mediat. Inflamm. 2018, 2018, 2037838. [Google Scholar] [CrossRef]
- Main, B.S.; Minter, M.R. Microbial Immuno-Communication in Neurodegenerative Diseases. Front. Neurosci. 2017, 11, 151. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Useros, N.; Nova, E.; González-Zancada, N.; Díaz, L.E.; Gómez-Martínez, S.; Marcos, A. Microbiota and Lifestyle: A Special Focus on Diet. Nutrients 2020, 12, 1776. [Google Scholar] [CrossRef]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Ross, R.P.; Stanton, C. Impact of Antibiotics on the Human Microbiome and Consequences for Host Health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121. [Google Scholar] [CrossRef]
- Rostami, F.M.; Mousavi, H.; Mousavi, M.R.N.; Shahsafi, M. Efficacy of Probiotics in Prevention and Treatment of Infectious Diseases. Clin. Microbiol. Newsl. 2018, 40, 97–103. [Google Scholar] [CrossRef]
- Meurman, J.H.; Stamatova, I. Probiotics: Contributions to Oral Health. Oral Dis. 2007, 13, 443–451. [Google Scholar] [CrossRef]
- Morrow, L.E.; Gogineni, V.; Malesker, M.A. Probiotics in the Intensive Care Unit. Nutr. Clin. Pract. 2012, 27, 235–241. [Google Scholar] [CrossRef]
- Fontana, L.; Bermudez-Brito, M.; Plaza-Diaz, J.; Muñoz-Quezada, S.; Gil, A. Sources, Isolation, Characterisation and Evaluation of Probiotics. Br. J. Nutr. 2013, 109, S35–S50. [Google Scholar] [CrossRef]
- Dekaboruah, E.; Suryavanshi, M.V.; Chettri, D.; Verma, A.K. Human Microbiome: An Academic Update on Human Body Site Specific Surveillance and Its Possible Role. Arch. Microbiol. 2020, 202, 2147–2167. [Google Scholar] [CrossRef] [PubMed]
- Belizário, J.E.; Napolitano, M. Human Microbiomes and Their Roles in Dysbiosis, Common Diseases, and Novel Therapeutic Approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef]
- Dubourg, G.; Lagier, J.C.; Armougom, F.; Robert, C.; Audoly, G.; Papazian, L.; Raoult, D. High-Level Colonisation of the Human Gut by Verrucomicrobia Following Broad-Spectrum Antibiotic Treatment. Int. J. Antimicrob. Agents 2013, 41, 149–155. [Google Scholar] [CrossRef]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic Isolates from Unconventional Sources: A Review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [PubMed]
- Roobab, U.; Batool, Z.; Manzoor, M.F.; Shabbir, M.A.; Khan, M.R.; Aadil, R.M. Sources, Formulations, Advanced Delivery and Health Benefits of Probiotics. Curr. Opin. Food Sci. 2020, 32, 17–28. [Google Scholar] [CrossRef]
- Verna, E.C.; Lucak, S. Use of Probiotics in Gastrointestinal Disorders: What to Recommend? Ther. Adv. Gastroenterol. 2010, 3, 307. [Google Scholar] [CrossRef] [PubMed]
- Culligan, E.P.; Hill, C.; Sleator, R.D. Probiotics and Gastrointestinal Disease: Successes, Problems and Future Prospects. Gut Pathog. 2009, 1, 19. [Google Scholar] [CrossRef]
- Montassier, E.; Valdés-Mas, R.; Batard, E.; Zmora, N.; Dori-Bachash, M.; Suez, J.; Elinav, E. Probiotics Impact the Antibiotic Resistance Gene Reservoir along the Human GI Tract in a Person-Specific and Antibiotic-Dependent Manner. Nat. Microbiol. 2021, 6, 1043–1054. [Google Scholar] [CrossRef]
- Lee, J.H.; Jin, Y.H.; Park, Y.K.; Yun, S.J.; Mah, J.H. Formation of Biogenic Amines in Pa (Green Onion) Kimchi and Gat (Mustard Leaf) Kimchi. Foods 2019, 8, 109. [Google Scholar] [CrossRef]
- Lee, Y.C. Kimchi: The Famous Fermented Vegetable Product in Korea. Food Rev. Int. 2009, 7, 399–415. [Google Scholar] [CrossRef]
- Song, H.S.; Lee, S.H.; Ahn, S.W.; Kim, J.Y.; Rhee, J.K.; Roh, S.W. Effects of the Main Ingredients of the Fermented Food, Kimchi, on Bacterial Composition and Metabolite Profile. Food Res. Int. 2021, 149, 110668. [Google Scholar] [CrossRef]
- Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients 2022, 14, 2038. [Google Scholar] [CrossRef]
- Jeong, D.W.; Lee, J.H. Antibiotic Resistance, Hemolysis and Biogenic Amine Production Assessments of Leuconostoc and Weissella Isolates for Kimchi Starter Development. LWT-Food Sci. Technol. 2015, 64, 1078–1084. [Google Scholar] [CrossRef]
- Park, S.; Saravanakumar, K.; Sathiyaseelan, A.; Park, S.J.; Hu, X.; Wang, M.H. Cellular Antioxidant Properties of Nontoxic Exopolysaccharide Extracted from Lactobacillales (Weissella Cibaria) Isolated from Korean Kimchi. LWT 2022, 154, 112727. [Google Scholar] [CrossRef]
- Song, M.W.; Chung, Y.; Kim, K.T.; Hong, W.S.; Chang, H.J.; Paik, H.D. Probiotic Characteristics of Lactobacillus Brevis B13-2 Isolated from Kimchi and Investigation of Antioxidant and Immune-Modulating Abilities of Its Heat-Killed Cells. LWT 2020, 128, 109452. [Google Scholar] [CrossRef]
- Cutting, S.M. Bacillus Probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Casula, G.; Cutting, S.M. Bacillus Probiotics: Spore Germination in the Gastrointestinal Tract. Appl. Environ. Microbiol. 2002, 68, 2344–2352. [Google Scholar] [CrossRef]
- Mazkour, S.; Shekarforoush, S.S.; Basiri, S.; Nazifi, S.; Yektaseresht, A.; Honarmand, M. Effects of Two Probiotic Spores of Bacillus Species on Hematological, Biochemical, and Inflammatory Parameters in Salmonella Typhimurium Infected Rats. Sci. Rep. 2020, 10, 8035. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus Probiotics: An Alternative to Antibiotics for Livestock Production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Koehler, T.M.; Lereclus, D. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7, GPP3-0032-2018. [Google Scholar] [CrossRef]
- Pilo, P.; Frey, J. Pathogenicity, Population Genetics and Dissemination of Bacillus Anthracis. Infect. Genet. Evol. 2018, 64, 115–125. [Google Scholar] [CrossRef]
- McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Strain-Specificity and Disease-Specificity of Probiotic Efficacy: A Systematic Review and Meta-Analysis. Front. Med. (Lausanne) 2018, 5, 124. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [Green Version]
- Jasim, B.; Sreelakshmi, S.; Mathew, J.; Radhakrishnan, E.K. Identification of Endophytic Bacillus Mojavensis with Highly Specialized Broad Spectrum Antibacterial Activity. 3 Biotech 2016, 6, 187. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Zeigler, D.R. Promotion of Bacillus Subtilis Subsp. Inaquosorum, Bacillus Subtilis Subsp. Spizizenii and Bacillus Subtilis Subsp. Stercoris to Species Status. Antonie Van Leeuwenhoek 2019, 113, 1–12. [Google Scholar] [CrossRef]
- Won, S.M.; Chen, S.; Park, K.W.; Yoon, J.H. Isolation of Lactic Acid Bacteria from Kimchi and Screening of Lactobacillus Sakei ADM14 with Anti-Adipogenic Effect and Potential Probiotic Properties. LWT 2020, 126, 109296. [Google Scholar] [CrossRef]
- Shaker, R.; Osaili, T.; Al-Omary, W.; Jaradat, Z.; Al-Zuby, M. Isolation of Enterobacter Sakazakii and Other Enterobacter Sp. from Food and Food Production Environments. Food Control 2007, 18, 1241–1245. [Google Scholar] [CrossRef]
- Riethorst, D.; Mols, R.; Duchateau, G.; Tack, J.; Brouwers, J.; Augustijns, P. Characterization of Human Duodenal Fluids in Fasted and Fed State Conditions. J. Pharm. Sci. 2016, 105, 673–681. [Google Scholar] [CrossRef]
- Evans, D.F.; Pye, G.; Bramley, R.; Clark, A.G.; Dyson, T.J.; Hardcastle, J.D. Measurement of Gastrointestinal PH Profiles in Normal Ambulant Human Subjects. Gut 1988, 29, 1035. [Google Scholar] [CrossRef]
- Hutt, C.A.; Keefe, D.M. GRAS Notice (GRN) No. 831. 2018. Available online: https://www.fda.gov/food/generally-recognized-safe-gras/gras-notice-inventory (accessed on 21 January 2022).
- Jang, H.J.; Lee, N.K.; Paik, H.D. Probiotic Characterization of Lactobacillus Brevis KU15153 Showing Antimicrobial and Antioxidant Effect Isolated from Kimchi. Food Sci. Biotechnol. 2019, 28, 1521. [Google Scholar] [CrossRef]
- Peres, C.M.; Alves, M.; Hernandez-Mendoza, A.; Moreira, L.; Silva, S.; Bronze, M.R.; Vilas-Boas, L.; Peres, C.; Malcata, F.X. Novel Isolates of Lactobacilli from Fermented Portuguese Olive as Potential Probiotics. LWT-Food Sci. Technol. 2014, 59, 234–246. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut Microbiota: Role in Pathogen Colonization, Immune Responses and Inflammatory Disease. Immunol. Rev. 2017, 279, 70. [Google Scholar] [CrossRef]
- Farid, W.; Masud, T.; Sohail, A.; Ahmad, N.; Naqvi, S.M.S.; Khan, S.; Ali, A.; Khalifa, S.A.; Hussain, A.; Ali, S.; et al. Gastrointestinal Transit Tolerance, Cell Surface Hydrophobicity, and Functional Attributes of Lactobacillus Acidophilus Strains Isolated from Indigenous Dahi. Food Sci. Nutr. 2021, 9, 5092. [Google Scholar] [CrossRef]
- Lakra, A.K.; Domdi, L.; Hanjon, G.; Tilwani, Y.M.; Arul, V. Some Probiotic Potential of Weissella Confusa MD1 and Weissella Cibaria MD2 Isolated from Fermented Batter. LWT 2020, 125, 109261. [Google Scholar] [CrossRef]
- Zeng, Z.; He, X.; Li, F.; Zhang, Y.; Huang, Z.; Wang, Y.; Li, K.; Bao, Y.; Iqbal, M.; Fakhar-e-Alam Kulyar, M.; et al. Probiotic Properties of Bacillus proteolyticus Isolated From Tibetan Yaks, China. Front. Microbiol. 2021, 12, 649207. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Lavigne, J.P.; Pagès, J.M. Enterobacter Spp.: Update on Taxonomy, Clinical Aspects, and Emerging Antimicrobial Resistance. Clin. Microbiol. Rev. 2019, 32, e00002-19. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, H.; Stindl, S.; Ludwig, W.; Stumpf, A.; Mehlen, A.; Monget, D.; Pierard, D.; Ziesing, S.; Heesemann, J.; Roggenkamp, A.; et al. Enterobacter Hormaechei Subsp. Oharae Subsp. Nov., E. Hormaechei Subsp. Hormaechei Comb. Nov., and E. Hormaechei Subsp. Steigerwaltii Subsp. Nov., Three New Subspecies of Clinical Importance. J. Clin. Microbiol. 2005, 43, 3297. [Google Scholar] [CrossRef]
- Erginkaya, Z.; Turhan, E.U.; Tatli, D. Determination of Antibiotic Resistance of Lactic Acid Bacteria Isolated from Traditional Turkish Fermented Dairy Products. Iran. J. Vet. Res. 2018, 19, 53. [Google Scholar] [CrossRef]
- Szutowska, J.; Gwiazdowska, D. Probiotic Potential of Lactic Acid Bacteria Obtained from Fermented Curly Kale Juice. Arch. Microbiol. 2021, 203, 975–988. [Google Scholar] [CrossRef]
- Sornsenee, P.; Singkhamanan, K.; Sangkhathat, S.; Saengsuwan, P.; Romyasamit, C. Probiotic Properties of Lactobacillus Species Isolated from Fermented Palm Sap in Thailand. Probiotics Antimicrob. Proteins 2021, 13, 957–969. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, R.; Tian, X.; Zhou, X.; Pan, X.; Wong, A. Assessing the Risk of Probiotic Dietary Supplements in the Context of Antibiotic Resistance. Front. Microbiol. 2017, 8, 908. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef]
- Kostelac, D.; Gerić, M.; Gajski, G.; Markov, K.; Domijan, A.M.; Čanak, I.; Jakopović, Ž.; Svetec, I.K.; Žunar, B.; Frece, J. Lactic Acid Bacteria Isolated from Equid Milk and Their Extracellular Metabolites Show Great Probiotic Properties and Anti-Inflammatory Potential. Int. Dairy J. 2021, 112, 104828. [Google Scholar] [CrossRef]
- Ge, Q.; Yang, B.; Liu, R.; Jiang, D.; Yu, H.; Wu, M.; Zhang, W. Antioxidant Activity of Lactobacillus Plantarum NJAU-01 in an Animal Model of Aging. BMC Microbiol. 2021, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Alkalbani, N.S.; Turner, M.S.; Ayyash, M.M. Isolation, Identification, and Potential Probiotic Characterization of Isolated Lactic Acid Bacteria and in Vitro Investigation of the Cytotoxicity, Antioxidant, and Antidiabetic Activities in Fermented Sausage. Microb. Cell Fact. 2019, 18, 188. [Google Scholar] [CrossRef]
- Avery, S.E.; Ruzbarsky, S.P.; Hise, A.M.; Schreier, H.J. Antibacterial Activity of Bacillus Inaquosorum Strain T1 against PirABVp-Bearing Vibrio Parahaemolyticus: Genetic and Physiological Characterization. Appl. Environ. Microbiol. 2020, 86, e01950-20. [Google Scholar] [CrossRef]
- Knight, C.A.; Bowman, M.J.; Frederick, L.; Day, A.; Lee, C.; Dunlap, C.A. The First Report of Antifungal Lipopeptide Production by a Bacillus Subtilis Subsp. Inaquosorum Strain. Microbiol. Res. 2018, 216, 40–46. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus Subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Park, J.H.; Saravanakumar, G.; Kim, K.; Kwon, I.C. Targeted Delivery of Low Molecular Drugs Using Chitosan and Its Derivatives. Adv. Drug Deliv. Rev. 2010, 62, 28–41. [Google Scholar] [CrossRef]
- Jung, W.J.; Mabood, F.; Souleimanov, A.; Zhou, X.; Jaoua, S.; Kamoun, F.; Smith, D.L. Stability and Antibacterial Activity of Bacteriocins Produced by Bacillus Thuringiensis and Bacillus Thuringiensis Ssp. Kurstaki. J. Microbiol. Biotechnol. 2008, 18, 1836–1840. [Google Scholar] [CrossRef]
- Rani, R.P.; Anandharaj, M.; Hema, S.; Deepika, R.; Ravindran, A.D. Purification of Antilisterial Peptide (Subtilosin A) from Novel Bacillus Tequilensis FR9 and Demonstrate Their Pathogen Invasion Protection Ability Using Human Carcinoma Cell Line. Front. Microbiol. 2016, 7, 1910. [Google Scholar] [CrossRef]
- Argentini, C.; Fontana, F.; Alessandri, G.; Lugli, G.A.; Mancabelli, L.; Ossiprandi, M.C.; van Sinderen, D.; Ventura, M.; Milani, C.; Turroni, F. Evaluation of Modulatory Activities of Lactobacillus Crispatus Strains in the Context of the Vaginal Microbiota. Microbiol. Spectr. 2022, 10, e02733-21. [Google Scholar] [CrossRef]
- Somashekaraiah, R.; Shruthi, B.; Deepthi, B.V.; Sreenivasa, M.Y. Probiotic Properties of Lactic Acid Bacteria Isolated from Neera: A Naturally Fermenting Coconut Palm Nectar. Front. Microbiol. 2019, 10, 1382. [Google Scholar] [CrossRef]
- Mohammed, S.; Çon, A.H. Isolation and Characterization of Potential Probiotic Lactic Acid Bacteria from Traditional Cheese. LWT 2021, 152, 112319. [Google Scholar] [CrossRef]
- Albayrak, Ç.B.; Duran, M. Isolation and Characterization of Aroma Producing Lactic Acid Bacteria from Artisanal White Cheese for Multifunctional Properties. LWT 2021, 150, 112053. [Google Scholar] [CrossRef]
- Vasiee, A.; Falah, F.; Behbahani, B.A.; Tabatabaee-yazdi, F. Probiotic Characterization of Pediococcus Strains Isolated from Iranian Cereal-Dairy Fermented Product: Interaction with Pathogenic Bacteria and the Enteric Cell Line Caco-2. J. Biosci. Bioeng. 2020, 130, 471–479. [Google Scholar] [CrossRef]
- Xia, A.N.; Meng, X.S.; Tang, X.J.; Zhang, Y.Z.; Lei, S.M.; Liu, Y.G. Probiotic and Related Properties of a Novel Lactic Acid Bacteria Strain Isolated from Fermented Rose Jam. LWT 2021, 136, 110327. [Google Scholar] [CrossRef]
- Sathiyaseelan, A.; Park, S.; Saravanakumar, K.; Mariadoss, A.V.A.; Wang, M.-H. Evaluation of Phytochemicals, Antioxidants, and Antidiabetic Efficacy of Various Solvent Fractions of Gynura procumbens (Lour.) Merr. Process Biochem. 2021, 111, 51–62. [Google Scholar] [CrossRef]
- Sathiyaseelan, A.; Saravanakumar, K.; Mariadoss, A.V.A.; Ramachandran, C.; Hu, X.; Oh, D.H.; Wang, M.H. Chitosan-Tea Tree Oil Nanoemulsion and Calcium Chloride Tailored Edible Coating Increase the Shelf Life of Fresh Cut Red Bell Pepper. Prog. Org. Coat. 2020, 151, 106010. [Google Scholar] [CrossRef]
- Estilarte, M.L.; Tymczyszyn, E.E.; de los Ángeles Serradell, M.; Carasi, P. Freeze-Drying of Enterococcus Durans: Effect on Their Probiotics and Biopreservative Properties. LWT 2021, 137, 110496. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Organism | Isolation Source | GenBank Accession Number |
|---|---|---|---|
| KNUAS001 | Bacillus proteolyticus | Fruit juice of Prunus domestica | OM327557 |
| KNUAS002 | Bacillus fungorum | Rhizome juice of Zingiber officinale | OM327558 |
| KNUAS003 | Bacillus subtilis | OM327559 | |
| KNUAS004 | Bacillus pseudomycoides | Commercial soybean paste | OM327560 |
| KNUAS005 | Bacillus thuringiensis | OM327561 | |
| KNUAS006 | Bacillus bingmayongensis | OM327562 | |
| KNUAS007 | Bacillus luti | OM327563 | |
| KNUAS008 | Enterobacter hormaechei | Fruit juice of red Prunus mume | OM327564 |
| KNUAS012 | Bacillus proteolyticus | Homemade soybean paste | OM327568 |
| KNUAS013 | Bacillus wiedmannii | OM327569 | |
| KNUAS014 | Bacillus mojavensis | Green onion Kimchi | OM327570 |
| KNUAS015 | Bacillus tequilensis | OM327571 | |
| KNUAS016 | Bacillus inaquosorum | Mustard leaf Kimchi | OM327572 |
| KNUAS017 | Lactobacillu brevis (Levilactobacillus brevis) | OM327573 | |
| KNUAS018 | Lacotbacillus graminis (Latilactobacillus graminis) | Cabbage Kimchi | OM327574 |
| KNUAS019 | Lactobacillus sakei (Latilactobacillus sakei) | OM327575 |
| Bacterial Isolate | Cell Viability (log10 CFU/mL) | ||||
|---|---|---|---|---|---|
| Simulated Gastric Juice at pH 3.0 | Intestinal Fluid (Bile Salts (1%) and Pancreatin (0.1%)) | ||||
| 1 h | 3 h | 1 h | 2 h | 4 h | |
| B. proteolyticus KNUAS001 | 6.72 ± 0.04 b | 7.42 ± 0.05 b | 7.95 ± 0.02 a | 7.54 ± 0.02 a | 6.46 ± 0.06 a |
| B. fungorum KNUAS002 | 6.93 ± 0.07 b | 7.89 ± 0.08 b | 6.96 ± 0.03 b | 6.47 ± 0.04 b | 6.19 ± 0.07 a |
| B. subtilis KNUAS003 | 5.84 ± 0.02 c | 5.95 ± 0.02 d | 6.73 ± 0.02 b | 6.55 ± 0.03 b | 6.21 ± 0.05 a |
| B. pseudomycoides KNUAS004 | 7.26 ± 0.05 a | 8.12 ± 0.08 a | 7.45 ± 0.04 a | 7.24 ± 0.05 a | 6.34 ± 0.02 a |
| B. thuringiensis KNUAS005 | 6.21 ± 0.03 b | 6.76 ± 0.03 c | 6.21 ± 0.03 b | 6.12 ± 0.03 b | 6.02 ± 0.07 a |
| B. bingmayongensis KNUAS006 | 7.21 ± 0.05 a | 8.14 ± 0.07 a | 7.45 ± 0.05 a | 7.08 ± 0.02 a | 6.28 ± 0.05 a |
| B. luti KNUAS007 | 6.58 ± 0.02 b | 6.97 ± 0.03 c | 7.87 ± 0.02 a | 7.52 ± 0.04 a | 5.84 ± 0.04 b |
| E. hormaechei KNUAS008 | 6.78 ± 0.04 b | 7.05 ± 0.05 b | 6.38 ± 0.04 b | 6.21 ± 0.08 b | 5.97 ± 0.03 b |
| B. proteolyticus KNUAS012 | 7.46 ± 0.07 a | 6.59 ± 0.04 c | 7.46 ± 0.07 a | 7.07 ± 0.06 a | 6.54 ± 0.07 a |
| B. wiedmannii KNUAS013 | 7.25 ± 0.08 a | 5.37 ± 0.01 d | 6.28 ± 0.04 b | 6.94 ± 0.02 b | 6.24 ± 0.08 a |
| B. mojavensis KNUAS014 | 5.87 ± 0.04 c | 6.64 ± 0.05 c | 6.36 ± 0.02 b | 6.22 ± 0.07 b | 5.65 ± 0.05 b |
| B. tequilensis KNUAS015 | 6.79 ± 0.02 b | 3.47 ± 0.02 e | 5.48 ± 0.05 c | 5.25 ± 0.05 c | 5.15 ± 0.04 b |
| B. inaquosorum KNUAS016 | 7.86 ± 0.04 a | 8.25 ± 0.08 a | 7.98 ± 0.03 a | 7.74 ± 0.01 a | 6.86 ± 0.02 a |
| L. brevis KNUAS017 | 6.31 ± 0.09 b | 6.17 ± 0.04 c | 6.24 ± 0.02 b | 6.09 ± 0.07 b | 5.37 ± 0.01 b |
| L. graminis KNUAS018 | 2.48 ± 0.04 e | 1.56 ± 0.05 f | 2.81 ± 0.07 e | 2.61 ± 0.08 d | 2.21 ± 0.08 c |
| L. sakei KNUAS019 | 3.48 ± 0.03 d | 0.8 ± 0.02 g | 3.65 ± 0.04 d | 2.82 ± 0.04 d | 2.14 ± 0.05 c |
| Bacterial Isolates | Autoaggregation (%) | Coaggregation (%) | Hydrophobicity (%) | ||||
|---|---|---|---|---|---|---|---|
| 1 h | 3 h | 6 h | 1 h | 3 h | 6 h | Xylene | |
| B. proteolyticus KNUAS001 | 3.87 ± 0.42 g | 13.83 ± 0.21 e | 34.44 ± 0.59 e | 0.62 ± 5.73 h | 9.61 ± 0.24 f | 30.02 ± 0.33 d | 7.44 ± 0.75 d |
| B. fungorum KNUAS002 | 19.40 ± 0.55 b | 30.66 ± 0.25 b | 46.30 ± 0.14 d | 4.69 ± 0.40 f | 19.85 ± 0.08 e | 50.67 ± 0.26 b | 5.26 ± 1.28 e |
| B. subtilis KNUAS003 | 18.71 ± 0.38 c | 33.67 ± 0.26 a | 52.27 ± 0.23 c | 36.13 ± 0.46 a | 39.75 ± 0.23 b | 63.84 ± 0.19 a | 1.64 ± 0.38 f |
| B. pseudomycoides KNUAS004 | 18.60 ± 0.30 c | 24.89 ± 0.21 c | 49.08 ± 0.16 d | 5.82 ± 0.13 f | 22.37 ± 0.39 d | 46.09 ± 0.26 | 15.41 ± 0.40 b |
| B. thuringiensis KNUAS005 | 10.62 ± 0.39 d | 15.69 ± 0.41 e | 49.32 ± 0.04 d | 1.25 ± 0.24 g | 24.25 ± 0.22 c | 46.44 ± 0.45 c | 11.96 ± 0.73 c |
| B. bingmayongensis KNUAS006 | 6.62 ± 0.08 f | 8.85 ± 0.12 g | 49.73 ± 0.10 d | 1.61 ± 0.44 g | 24.34 ± 0.72 c | 51.63 ± 0.35 b | 0.09 ± 0.83 g |
| B. luti KNUAS007 | 2.54 ± 0.63 h | 20.41 ± 0.34 d | 48.86 ± 0.14 d | 2.68 ± 0.28 g | 20.82 ± 0.06 | 47.67 ± 0.27 c | 13.17 ± 0.76 c |
| E. hormaechei KNUAS008 | 19.45 ± 0.53 b | 26.19 ± 0.38 c | 56.85 ± 0.18 c | 26.02 ± 0.49 b | 48.46 ± 0.33 a | 62.58 ± 0.55 a | 0.24 ± 0.10 g |
| B. proteolyticus KNUAS012 | 3.08 ± 0.30 g | 10.75 ± 0.31 f | 33.73 ± 0.12 e | 8.76 e | 16.03 e | 49.12 ± 0.34 c | 12. 46 ± 0.06 c |
| B. wiedmannii KNUAS013 | 1.52 ± 0.22 i | 11.04 ± 0.33 f | 51.35 ± 0.27 c | 0.39 ± 6.04 | 9.68 ± 0.29 f | 50.39 ± 0.36 | 4.67 ± 0.17 e |
| B. mojavensis KNUAS014 | 7.24 ± 0.44 e | 8.58 ± 0.27 g | 56.98 ± 0.10 c | 14.76 ± 0.12 d | 20.11 ± 0.40 e | 61.67 ± 0.39 a | 1.78 ± 1.00 f |
| B. tequilensis KNUAS015 | 22.63 ± 0.43 a | 24.71 ± 0.18 c | 64.56 ± 0.14 a | 21.21 ± 0.29 c | 34.15 ± 0.55 b | 63.64 ± 0.35 a | 7.29 ± 0.63 d |
| B. inaquosorum KNUAS016 | 8.51 ± 0.20 e | 10.82 ± 0.18 f | 61.63 ± 0.19 b | 5.83 ± 0.19 f | 7.36 ± 0.42 f | 57.63 ± 0.48 b | 16.76 ± 0.16 b |
| L. brevis KNUAS017 | 8.58 ± 0.18 e | 21.58 ± 0.18 d | 29.98 ± 0.09 e | 0.75 ± 0.21 h | 27.45 ± 0.26 c | 33.86 ± 0.12 d | 25.64 ± 0.28 a |
| L. graminis KNUAS018 | 0.95 ± 0.17 i | 3.06 ± 0.33 h | 31.11 ± 0.19 e | 5.56 f | 12.14 ± 0.29 f | 45.71 ± 0.23 c | 18.81 ± 1.36 b |
| L. sakei KNUAS019 | 7.01 ± 0.23 e | 26.06 ± 0.11 c | 66.55 ± 0.33 | 0.66 ± 0.29 h | 24.71 ± 0.21 c | 55.69 ± 0.30 b | 14.91 ± 0.48 c |
| Zone of Inhibition (mm) | |||||
|---|---|---|---|---|---|
| Bacterial Isolates | TCH | VAN | ERY | GEN | AMP |
| B. proteolyticus KNUAS001 | 22.2 ± 0.4 b | 10.2 ± 0.3 c | 23.2 ± 0.3 c | 14.1 ± 0.1 b | 9.2 ± 0.3 d |
| B. fungorum KNUAS002 | 23.7 ± 0.4 b | 14.9 ± 0.1 b | 22.9 ± 0.1 c | 14.3 ± 0.4 b | 8.2 ± 0.4 d |
| B. subtilis KNUAS003 | 18.7 ± 1.1 c | 11.3 ± 0.4 b | 19.3 ± 0.5 d | 13.3 ± 0.5 b | 12.1 ± 0.1 d |
| B. pseudomycoides KNUAS004 | 25.9 ± 0.1 b | 17.7 ± 0.4 a | 25.8 ± 0.3 a | 18.2 ± 0.3 a | 39.7 ± 0.4 a |
| B. thuringiensis KNUAS005 | 22.2 ± 0.4 b | 17.8 ± 0.3 a | 27.1 ± 0.1 a | 12.2 ± 0.3 c | 21.8 ± 1.1 b |
| B. bingmayongensis KNUAS006 | 12.2 ± 0.3 d | 13.1 ± 0.1 b | 28.1 ± 0.1 a | 21.5 ± 0.7 a | 10.1 ± 0.2 d |
| B. luti KNUAS007 | 31.7 ± 0.4 a | 19.1 ± 0.2 a | 10.1 ± 0.1 e | 17.2 ± 0.3 a | 20.9 ± 0.1 b |
| E. hormaechei KNUAS008 | 18.3 ± 0.4 c | 12.2 ± 0.4 c | 22.2 ± 0.3 c | 12.3 ± 0.5 c | 22.1 ± 0.1 b |
| B. proteolyticus KNUAS012 | 28.8 ± 0.2 a | 0 | 22.3 ± 0.4 c | 17.4 ± 0.6 a | 39.4 ± 0.9 a |
| B. wiedmannii KNUAS013 | 25.3 ± 0.4 b | 18.2 ± 0.4 a | 23.1 ± 0.1 c | 0 | 41.3 ± 0.9 a |
| B. mojavensis KNUAS014 | 0 | 0 | 0 | 0 | 0 |
| B. tequilensis KNUAS015 | 10.9 ± 0.1 d | 14.1 ± 0.2 b | 16.2 ± 0.3 d | 12.1 ± 0.1 c | 18.1 ± 0.1 c |
| B. inaquosorum KNUAS016 | 0 | 0 | 0 | 0 | 0 |
| L. brevis KNUAS017 | 0 | 0 | 0 | 0 | 0 |
| L. graminis KNUAS018 | 24.8 ± 0.2 b | 0 | 24.9 ± 0.1 c | 0 | 23.2 ± 0.3 b |
| L. sakei KNUAS019 | 21.4 ± 0.6 b | 0 | 17.1 ± 0.2 d | 0 | 18.0 ± 0.0 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sathiyaseelan, A.; Saravanakumar, K.; Han, K.; Naveen, K.V.; Wang, M.-H. Antioxidant and Antibacterial Effects of Potential Probiotics Isolated from Korean Fermented Foods. Int. J. Mol. Sci. 2022, 23, 10062. https://doi.org/10.3390/ijms231710062
Sathiyaseelan A, Saravanakumar K, Han K, Naveen KV, Wang M-H. Antioxidant and Antibacterial Effects of Potential Probiotics Isolated from Korean Fermented Foods. International Journal of Molecular Sciences. 2022; 23(17):10062. https://doi.org/10.3390/ijms231710062
Chicago/Turabian StyleSathiyaseelan, Anbazhagan, Kandasamy Saravanakumar, Kiseok Han, Kumar Vishven Naveen, and Myeong-Hyeon Wang. 2022. "Antioxidant and Antibacterial Effects of Potential Probiotics Isolated from Korean Fermented Foods" International Journal of Molecular Sciences 23, no. 17: 10062. https://doi.org/10.3390/ijms231710062