
Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice

 ,
,
Abstract
:1. Introduction
2. Results
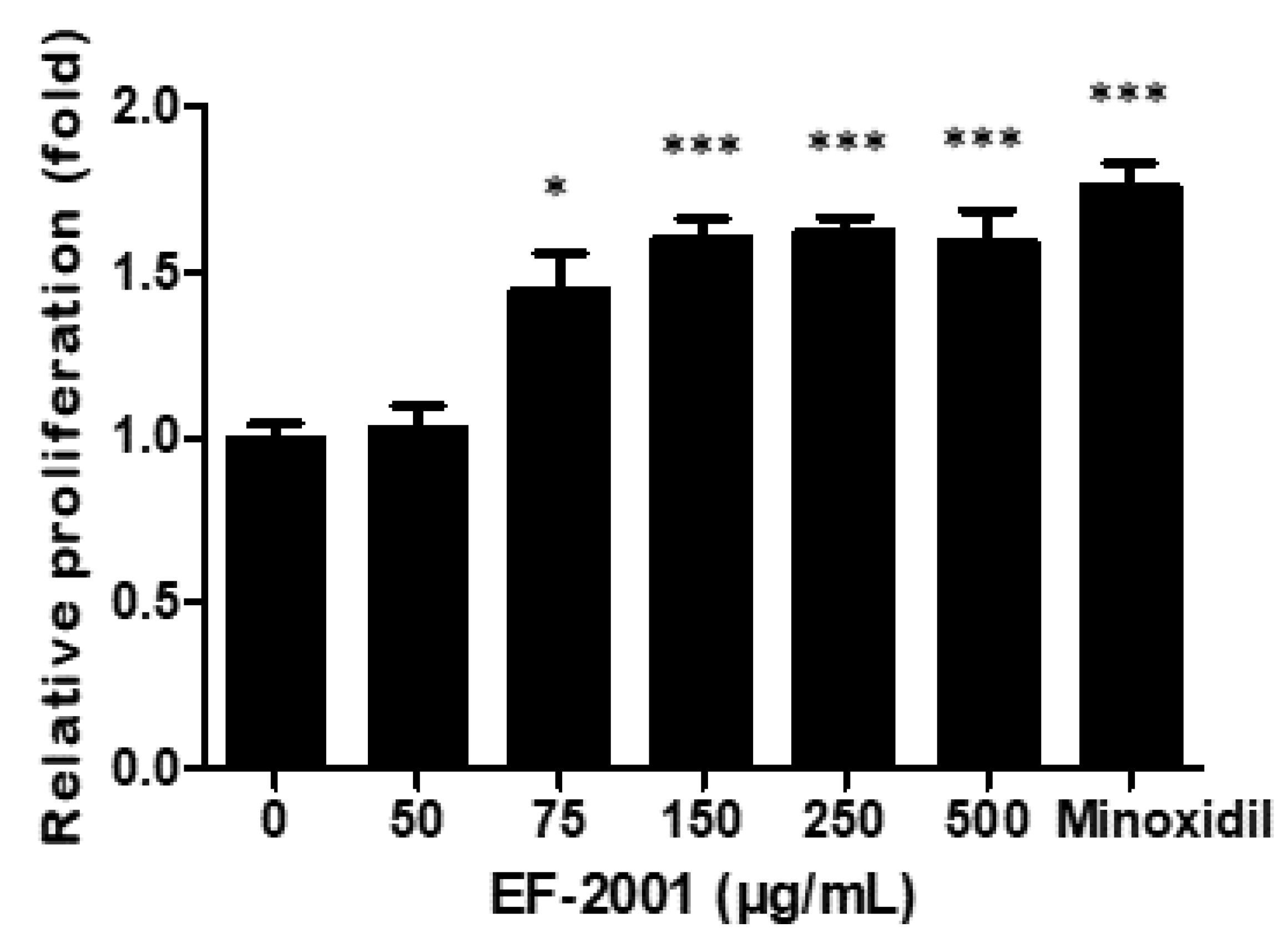
2.1. Effect of EF-2001 on HDPC Proliferation
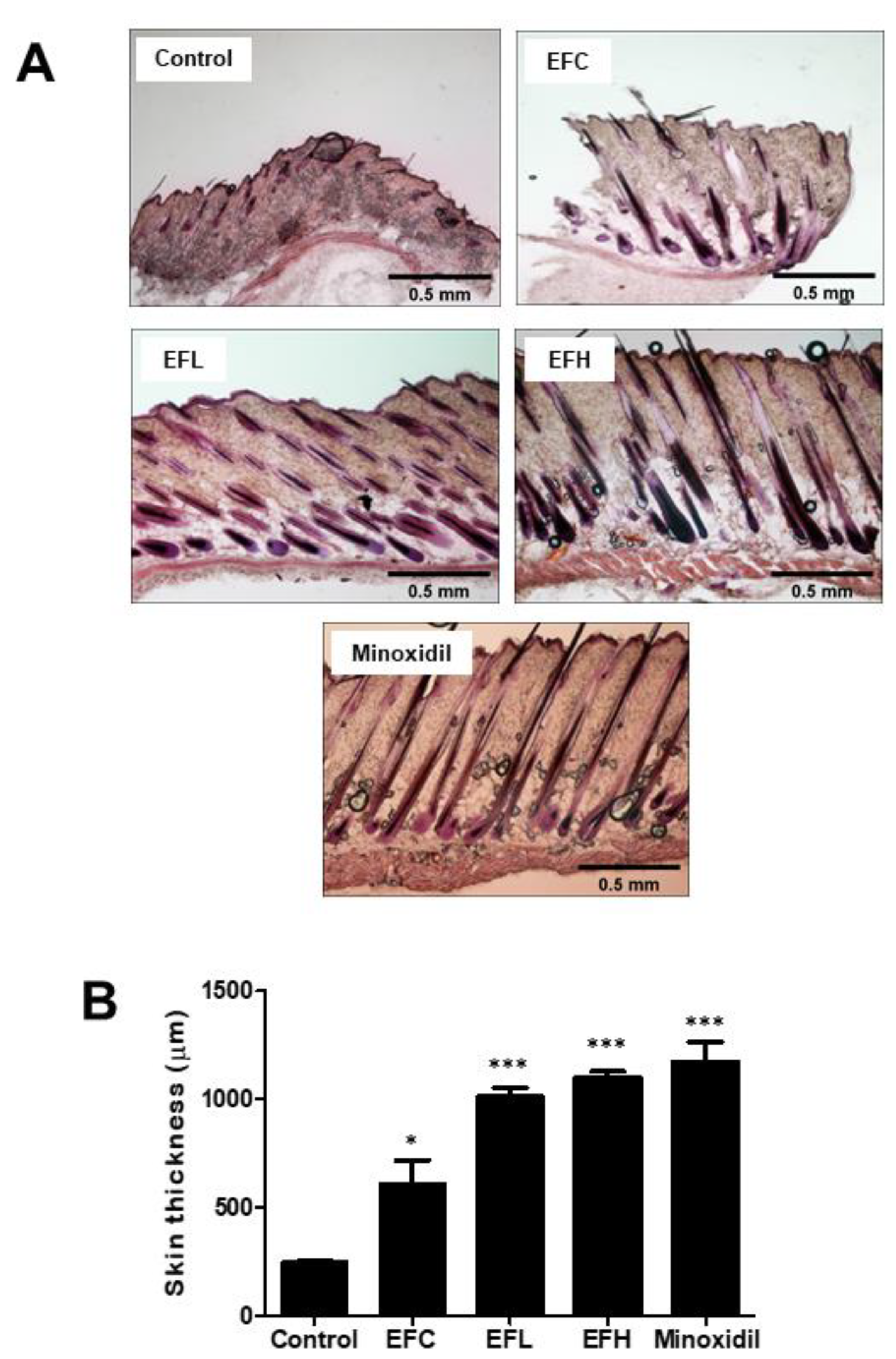
2.2. EF-2001 Induced the Anagen Stage in C57BL/6 Mice
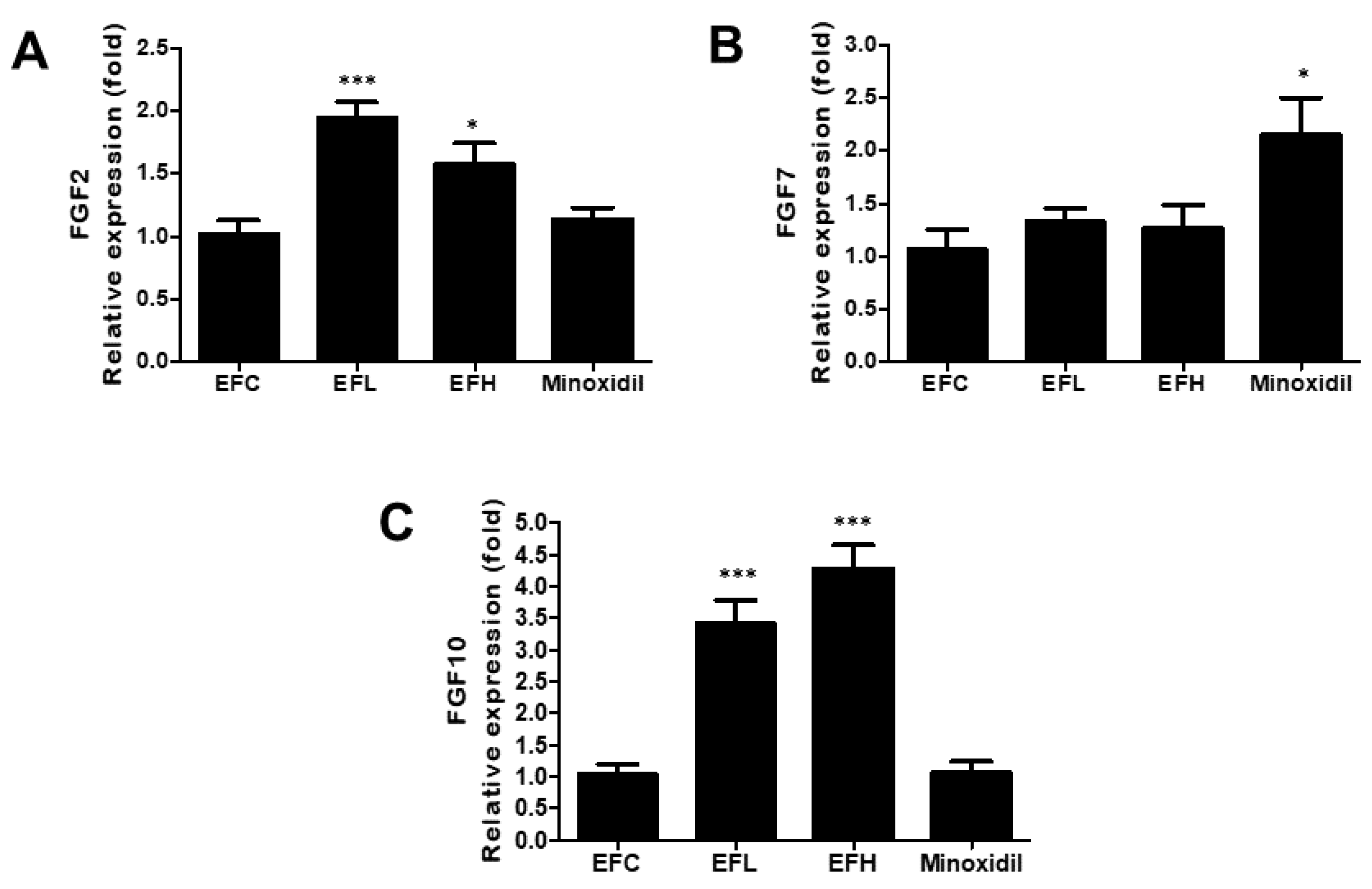
2.3. Effect of EF-2001 on the Expression of FGF2, FGF7, and FGF10 mRNA in C57BL/6 Male Mice
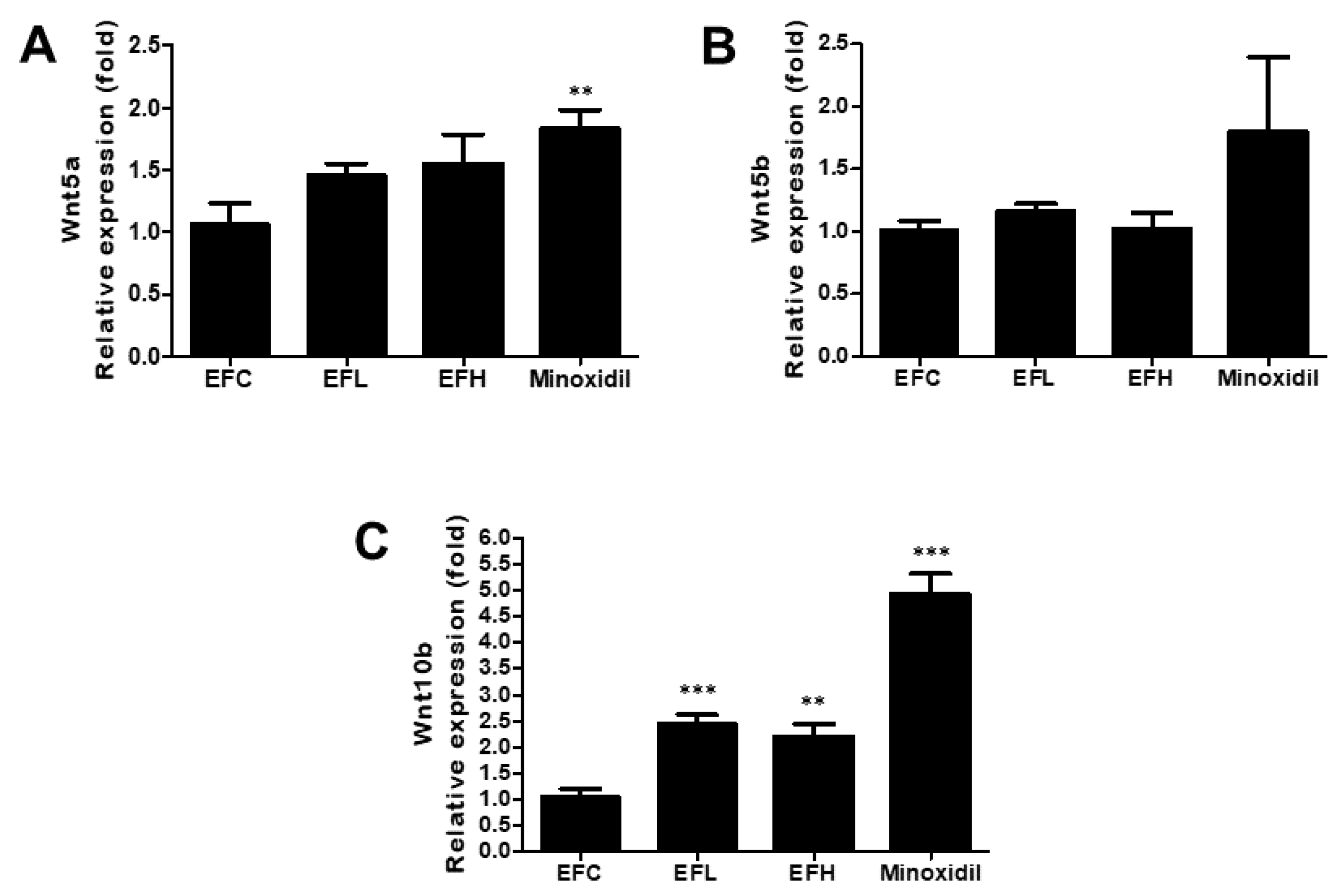
2.4. Effect of EF-2001 on the Expression of Wnt5a, Wnt5b, and Wnt10b mRNA in C57BL/6 Male Mice
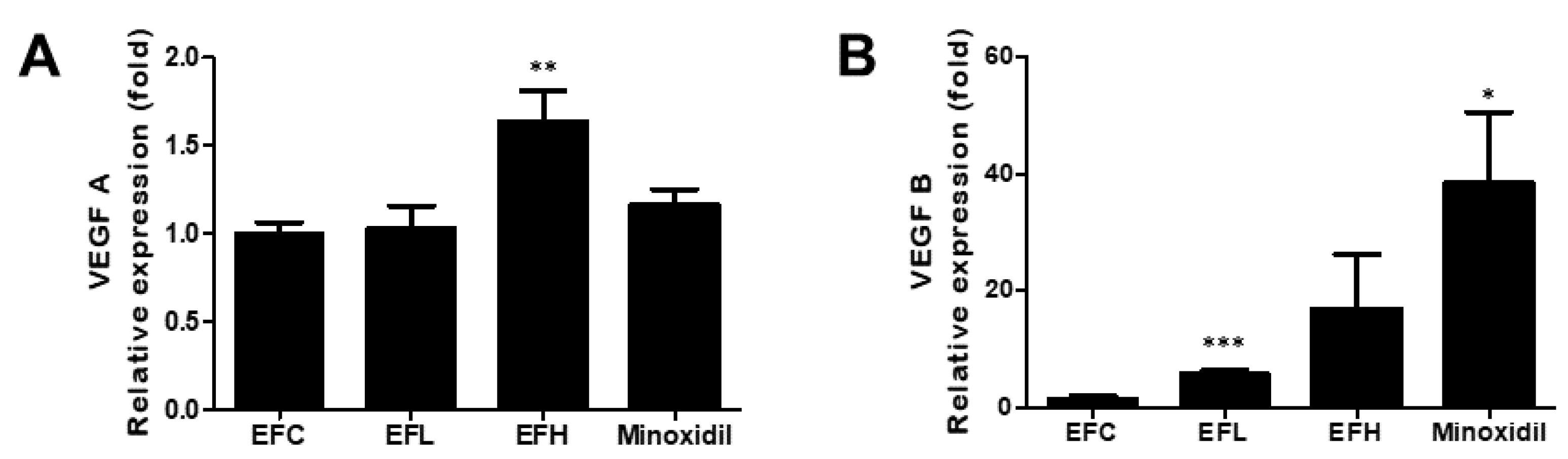
2.5. Effect of EF-2001 on the Expression of VEGF-A and VEGF-B mRNA in C57BL/6 Male Mice
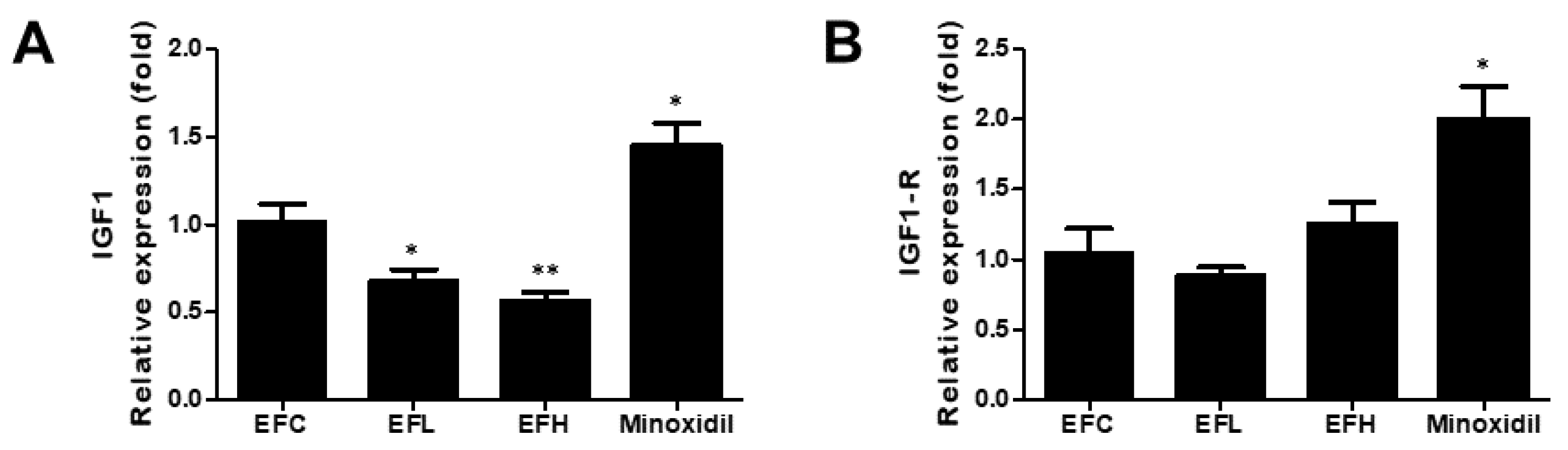
2.6. Effect of EF-2001 on the Expression of IGF1 and IGF1-R mRNA in C57BL/6 Male Mice
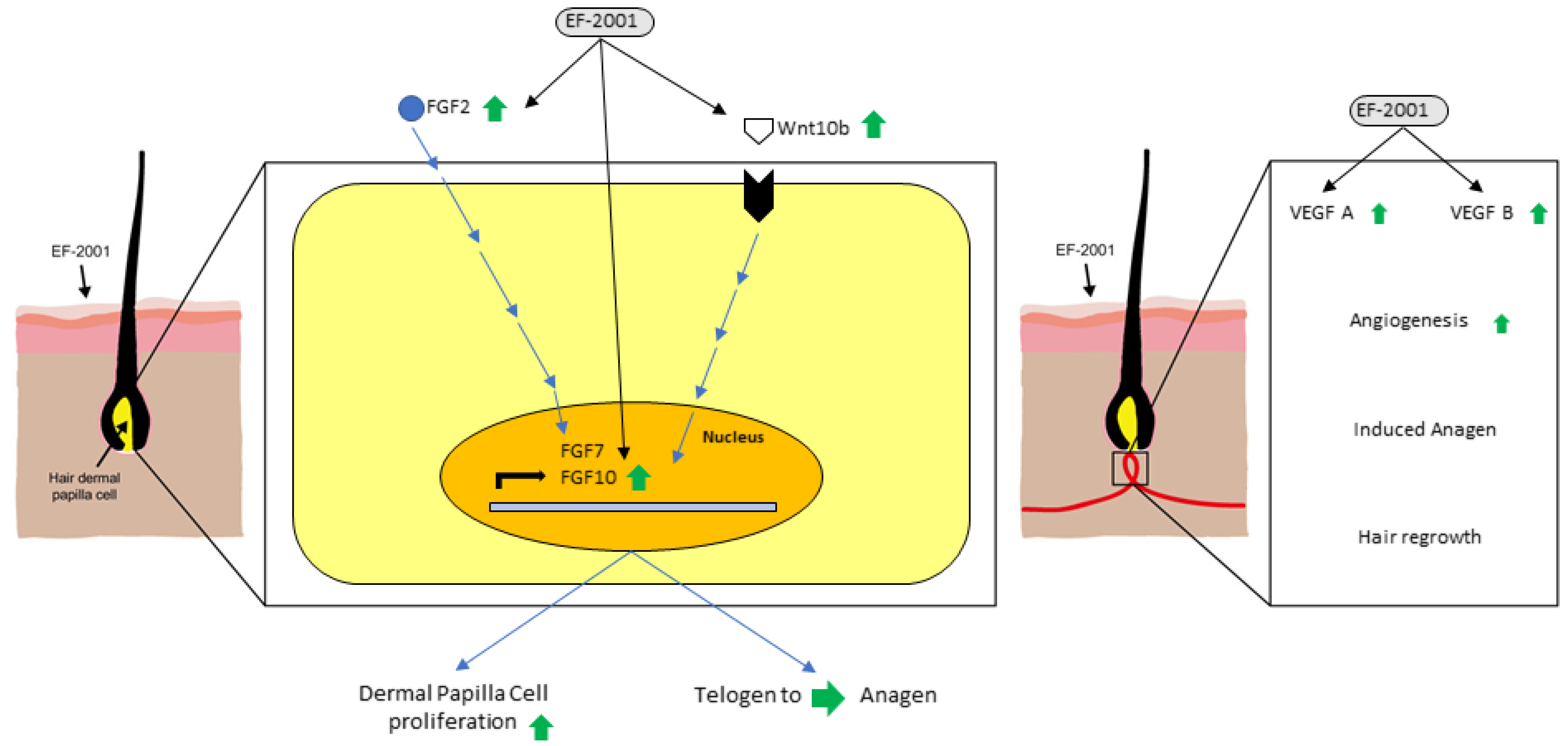
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. EF-2001 (Enterococcus faecalis-2001)
4.3. Cell Culture
4.4. Cell-Proliferation Assay
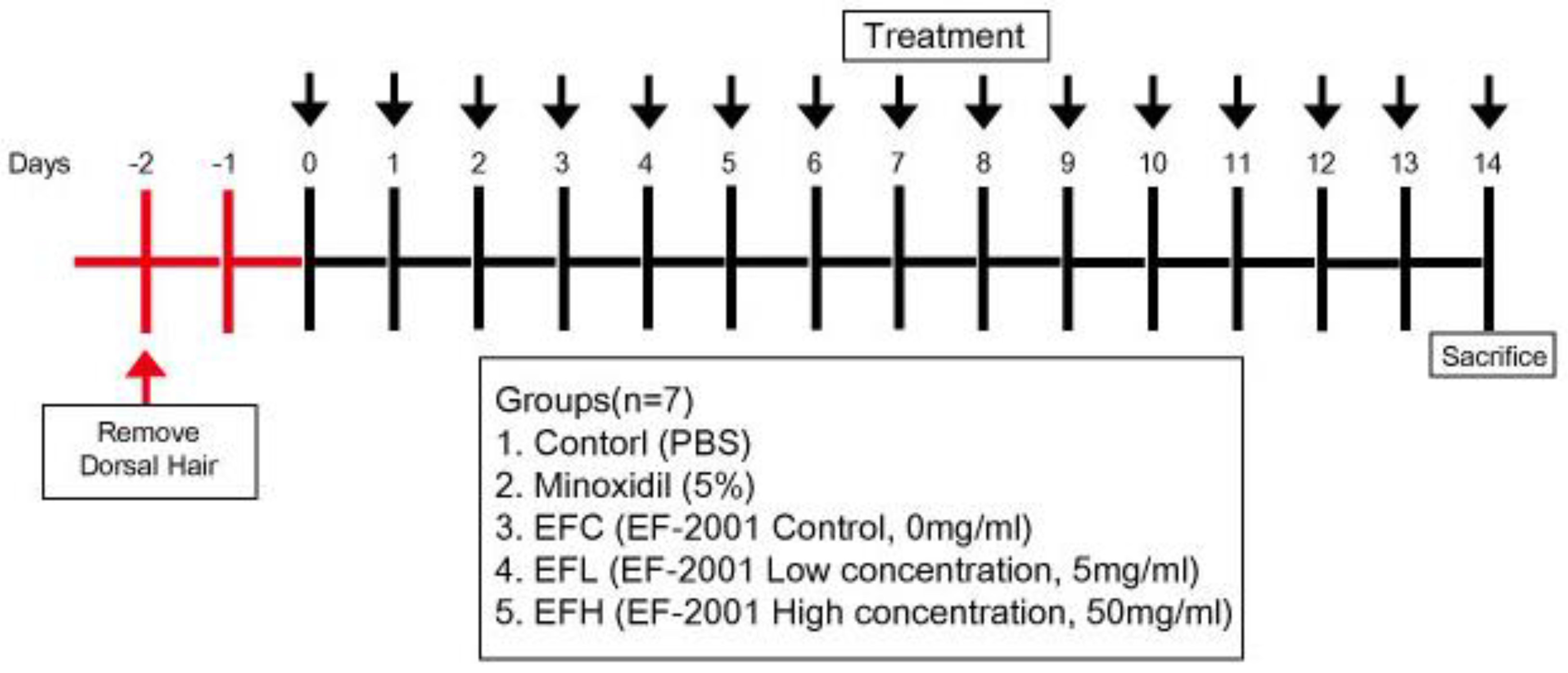
4.5. Animal Experiments
4.6. H&E Staining
4.7. RNA Preparation and qPCR
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stenn, K.S. Molecular insights into the hair follicle and its pathology: A review of recent developments. Int. J. Dermatol. 2003, 42, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.; Arck, P.C.; Paus, R. Hair growth inhibition by psychoemotional stress: A mouse model for neural mechanisms in hair growth control. Exp. Dermatol. 2006, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. Molecular mechanisms of androgenetic alopecia. Exp. Gerontol. 2002, 37, 981–990. [Google Scholar] [CrossRef]
- Lee, E.; Lee, K. A study on the behavioral of scalp, and care of losing hair of middle aged men’s hair-losers. J. Kor. Soc. Cosmetol. 2010, 16, 674–686. [Google Scholar]
- Park, J.; Lee, S.; Gwon, G. A study on the analysis of factor related to alopecia of women in 20~30 years old. J. Kor. Soc. B&A 2014, 15, 53–65. [Google Scholar]
- Olsen, E.A. Current and novel methods for assessing efficacy of hair growth promoters in pattern hair loss. J. Am. Acad. 2003, 48, 253–262. [Google Scholar] [CrossRef]
- Skalnaya, M.G.; Tkachev, V.P. Trace elements content and hormonal profiles in women with androgenetic alopecia. J. Trace Elem. Med. Biol. 2011, 25, S50–S53. [Google Scholar] [CrossRef]
- Alonso, L.; Fuchs, E. The hair cycle. J. Cell Sci. 2006, 119, 391–393. [Google Scholar] [CrossRef]
- Buhl, A.; Waldon, D.; Miller, B.; Brunden, M. Differences in activity of minoxidil and cyclosporin A on hair growth in nude and normal mice. Comparisons of in vivo and in vitro studies. Lab. Invest. 1990, 62, 104–107. [Google Scholar]
- Ahn, S.-Y.; Pi, L.-Q.; Hwang, S.T.; Lee, W.-S. Effect of IGF-I on hair growth is related to the anti-apoptotic effect of IGF-I and up-regulation of PDGF-A and PDGF-B. Ann. Dermatol. 2012, 24, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Cotsarelis, G.; Millar, S.E. Towards a molecular understanding of hair loss and its treatment. Trends Mol. Med. 2001, 7, 293–301. [Google Scholar] [CrossRef]
- Kiso, M.; Hamazaki, T.S.; Itoh, M.; Kikuchi, S.; Nakagawa, H.; Okochi, H. Synergistic effect of PDGF and FGF2 for cell proliferation and hair inductive activity in murine vibrissal dermal papilla in vitro. J. Dermatol. Sci. 2015, 79, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Won, C.H.; Yoo, H.G.; Kwon, O.S.; Sung, M.Y.; Kang, Y.J.; Chung, J.H.; Park, B.S.; Sung, J.-H.; Kim, W.S.; Kim, K.H. Hair growth promoting effects of adipose tissue-derived stem cells. J. Dermatol. Sci. 2010, 57, 134–137. [Google Scholar] [CrossRef] [PubMed]
- McElwee, K.J.; Shapiro, J. Promising therapies for treating and/or preventing androgenic alopecia. Skin Ther. Lett. 2012, 17, 1–4. [Google Scholar]
- Dargie, H.; Dollery, C.T.; Daniel, J. Minoxidil in resistant hypertension. Lancet 1977, 310, 515–518. [Google Scholar] [CrossRef]
- Messenger, A.G.; Rundergren, J. Minoxidil: Mechanisms of action on hair growth. Br. J. Dermatol. 2004, 150, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.L.; Marshall, A. Hypertrichosis due to minoxidil. Br. J. Dermatol. 1979, 101, 593–595. [Google Scholar] [CrossRef]
- Meisheri, K.D.; Cipkus, L.A.; Taylor, C.J. Mechanism of action of minoxidil sulfate-induced vasodilation: A role for increased K+ permeability. J. Pharmacol. Exp. Ther. 1988, 245, 751–760. [Google Scholar]
- Han, J.H.; Kwon, O.S.; Chung, J.H.; Cho, K.H.; Eun, H.C.; Kim, K.H. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J. Dermatol. Sci. 2004, 34, 91–98. [Google Scholar] [CrossRef]
- Glaser, R.L.; Dimitrakakis, C.; Messenger, A.G. Improvement in scalp hair growth in androgen-deficient women treated with testosterone: A questionnaire study. Br. J. Dermatol. 2012, 166, 274–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagemann, T.; Schlütter-Böhmer, B.; Allam, J.P.; Bieber, T.; Novak, N. Positive lymphocyte transformation test in a patient with allergic contact dermatitis of the scalp after short-term use of topical minoxidil solution. Contact Derm. 2005, 53, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Lee, M.; Kim, H.J.; Kwon, J.I.; Lee, Y. Effects of black soybean and fermented black soybean extracts on proliferation of human follicle dermal papilla cells. J. Korean Soc. Food Sci. 2017, 46, 671–680. [Google Scholar]
- Joo, S.-S. In vitro and in vivo hair growth promotion effects of Lactobacillus plantarum-fermented plant extracts (MBN). Korean J. Food Sci. Technol. 2011, 43, 381–386. [Google Scholar] [CrossRef]
- Kang, J.-I.; Moon, J.; Kim, E.-J.; Lee, Y.-K.; Koh, Y.-S.; Yoo, E.-S.; Kang, H.-K.; Yim, D. The hair growth effects of wheat bran. Saengyak Hakhoe Chi 2013, 44, 384–390. [Google Scholar]
- Park, H.; Kim, G.; Kim, S.; Kim, K. A screening of effective natural cosmetical materials for scalp care. J. Invest Cosmetol. 2011, 7, 115–122. [Google Scholar]
- Kanasugi, H.; Hasegawa, T.; Yamamoto, T.; Abe, S.; Yamaguchi, H. Optimal dose of enterococcal preparation (FK-23) supplemented perorally for stimulation of leukocyte reconstitution in dogs treated with cyclophosphamide. J. Vet. Med. Sci. 1996, 58, 563–565. [Google Scholar] [CrossRef] [Green Version]
- Satonaka, K.; Ohashi, K.; Nohmi, T.; Yamamoto, T.; Abe, S.; Uchida, K.; Yamaguchi, H. Prophylactic effect of Enterococcus faecalis FK-23 preparation on experimental candidiasis in Mice. Microbiol. Immunol. 1996, 40, 217–222. [Google Scholar] [CrossRef]
- Chang, S.-J.; Lee, M.-H.; Kim, W.-J.; Chae, Y.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Kim, T.-J. Effect of heat-killed Enterococcus faecalis, EF-2001 on C2C12 myoblast damage induced by oxidative stress and muscle volume decreased by sciatic denervation in C57BL/6 mice. J. Life Sci. 2019, 29, 215–222. [Google Scholar]
- Choi, M.-S.; Chang, S.-J.; Chae, Y.; Lee, M.-H.; Kim, W.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Kim, T.-J. Anti-inflammatory effect of Enterococcus faecalis, EF-2001. J. Life Sci. 2018, 28, 1361–1368. [Google Scholar]
- Choi, E.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Tang, Y.; Han, W.C.; Kim, E.-K.; Park, Z.-Y. Effect of Enterococcus faecalis EF-2001 on experimentally induced atopic eczema in mice. Food Sci. Biotechnol. 2016, 25, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-J.; Iwasa, M.; Hang, K.-I.; Kim, W.-J.; Tang, Y.; Hwang, Y.J.; Chae, J.R.; Han, W.C.; Shin, Y.-S.; Kim, E.-K. Heat-killed Enterococcus faecalis EF-2001 ameliorates atopic dermatitis in a murine model. Nutrients 2016, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, M.; Choi, Y.J.; Wedamulla, N.E.; Tang, Y.; Han, K.I.; Hwang, J.Y.; Kim, E.K. Heat-killed Enterococcus faecalis EF-2001 attenuate lipid accumulation in diet-induced obese (DIO) mice by activating AMPK signaling in liver. Foods 2022, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Woo, K.-J.; Kim, M.-A.; Hong, J.; Kim, J.; Kim, S.-H.; Han, K.-I.; Iwasa, M.; Kim, T.-J. Heat-killed Enterococcus faecalis prevents adipogenesis and high fat diet-induced obesity by inhibition of lipid accumulation through inhibiting C/EBP-α and PPAR-γ in the insulin signaling pathway. Nutrients 2022, 14, 1308. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Tong, C.; Hou, Q.; Han, Q.; Zhao, Y.; Liu, H. Treatment of MSCs with Wnt1a-conditioned medium activates DP cells and promotes hair follicle regrowth. Sci. Rep. 2014, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ramos, R.; Guerrero-Juarez, C.F.; Plikus, M.V. Hair follicle signaling networks: A dermal papilla-centric approach. J. Investig. Dermatol. 2013, 133, 2306–2308. [Google Scholar] [CrossRef] [Green Version]
- Müller-Röver, S.; Foitzik, K.; Paus, R.; Handjiski, B.; van der Veen, C.; Eichmüller, S.; McKay, I.A.; Stenn, K.S. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Balañá, M.E.; Charreau, H.E.; Leirós, G.J. Epidermal stem cells and skin tissue engineering in hair follicle regeneration. World J. Stem Cells 2015, 7, 711–727. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, M.; Tabata, Y. In vivo promoted growth of mice hair follicles by the controlled release of growth factors. Biomaterials 2003, 24, 2387–2394. [Google Scholar] [CrossRef]
- Hwang, D.; Lee, H.; Lee, J.; Lee, M.; Cho, S.; Kim, T.; Kim, H. Micro-Current Stimulation Has Potential Effects of Hair Growth-Promotion on Human Hair Follicle-Derived Papilla Cells and Animal Model. Int. J. Mol. Sci. 2021, 22, 4361. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Primer | Forward | Reverse |
|---|---|---|---|
| Mouse | Igf1 | 5′-GTCGTCTTCACACCTCTTCTACCT-3′ | 5′-GCACAGTACATCTCCAGTCTCCT-3′ |
| Igf1r | 5′-CTCAGGCTTCATCCGCAACAG-3′ | 5′-GTTCTCCAACTCCGAGGCAATG-3′ | |
| Wnt5a | 5′-CTGGCAGGACTTTCTCAAGG-3′ | 5′-CTCTAGCGTCCACGAACTCC-3′ | |
| Wnt5b | 5′-TCGGAGGAGCAGGGCCGAGC-3′ | 5′-CAGCTTGCCCTGGCGGGTGA-3′ | |
| Wnt10b | 5′-CCTGTCCGGACTGAGTAAGC-3′ | 5′-TTGCTCACCACTACCCTTCC-3′ | |
| Fgf2 | 5′-CAAGAACGGCGGCTTCTTC-3′ | 5′-GAAAGAAACAGTATGGCCT-3′ | |
| Fgf7 | 5′-AGACTGTTCTGTCGCACC-3′ | 5′-CCGCTGTGTGTCCATTTAG-3′ | |
| Fgf10 | 5′-TGTCCGCTGGAGAAGGCTGTTC-3′ | 5′-CTATGTTTGGATCGTCATGG-3′ | |
| Vegfa | 5′-CGAGATAGAGTACATCTTCAAGCC-3′ | 5′-TCATCGTTACAGCAGCCTGC-3′ | |
| Vegfb | 5′-AAAAAAAAAGGAGAGTGCTGTGAAG-3′ | 5′-TCCCAGCCCGGAACAGA-3′ | |
| Gapdh | 5′-GCCAAGGTCATCCATGACAACT-3′ | 5′-GAGGGGCCATCCACAGTCTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, Y.-H.; Lee, J.-H.; Chang, S.-J.; Chae, Y.; Lee, M.-H.; Kim, S.-H.; Han, K.-I.; Kim, T.-J. Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice. Int. J. Mol. Sci. 2022, 23, 5413. https://doi.org/10.3390/ijms23105413
Baek Y-H, Lee J-H, Chang S-J, Chae Y, Lee M-H, Kim S-H, Han K-I, Kim T-J. Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice. International Journal of Molecular Sciences. 2022; 23(10):5413. https://doi.org/10.3390/ijms23105413
Chicago/Turabian StyleBaek, Young-Hyun, Jin-Ho Lee, Sang-Jin Chang, Yuri Chae, Myung-Hun Lee, Sun-Hong Kim, Kwon-Il Han, and Tack-Joong Kim. 2022. "Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice" International Journal of Molecular Sciences 23, no. 10: 5413. https://doi.org/10.3390/ijms23105413