
The MdAux/IAA2 Transcription Repressor Regulates Cell and Fruit Size in Apple Fruit

 ,
,
Abstract
:1. Introduction
2. Results
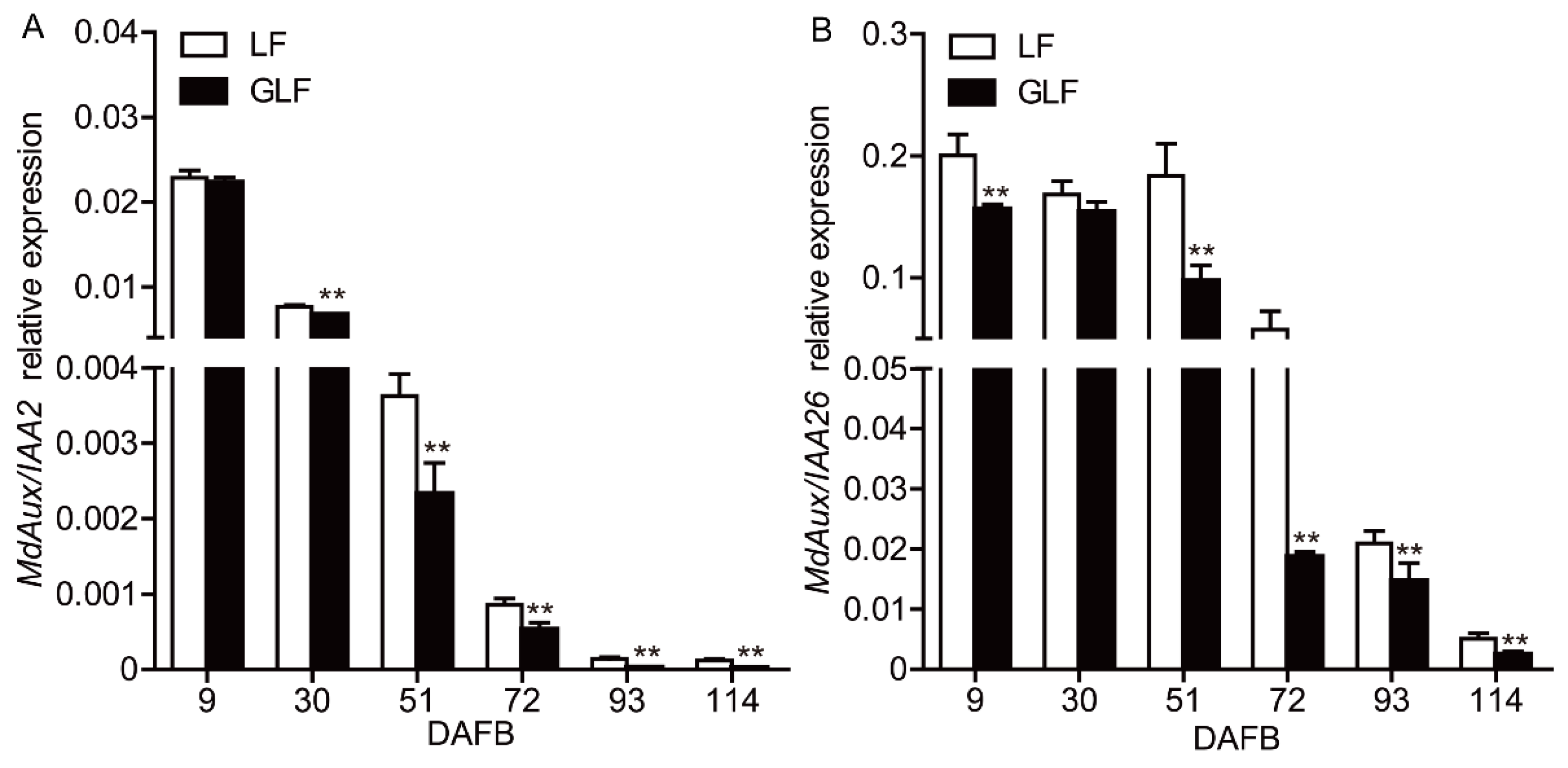
2.1. Expression of MdAux/IAA2 in GLF Was Lower Than That in LF Fruit
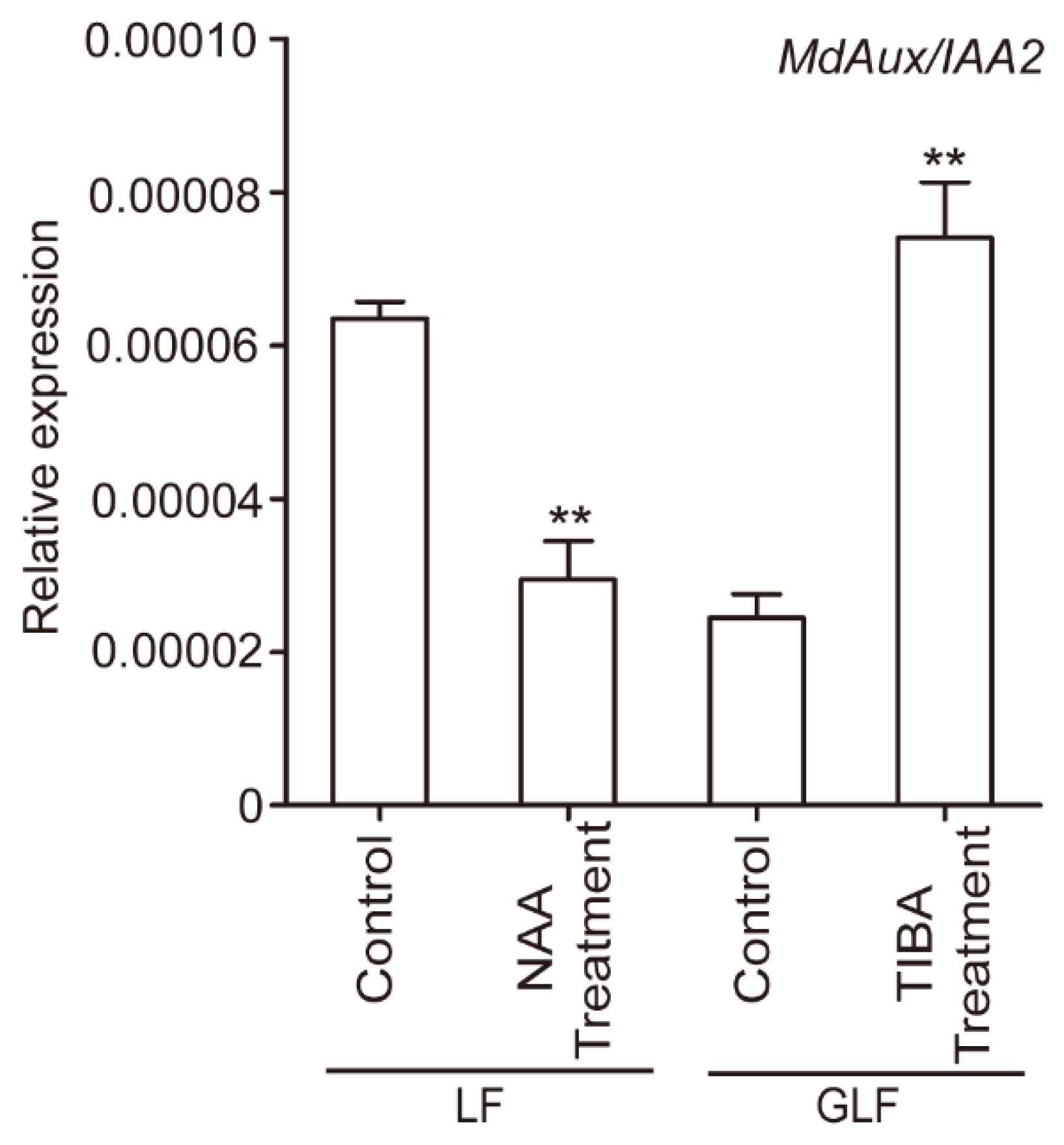
2.2. Exogenous Auxin Inhibited the Expression of MdAux/IAA2
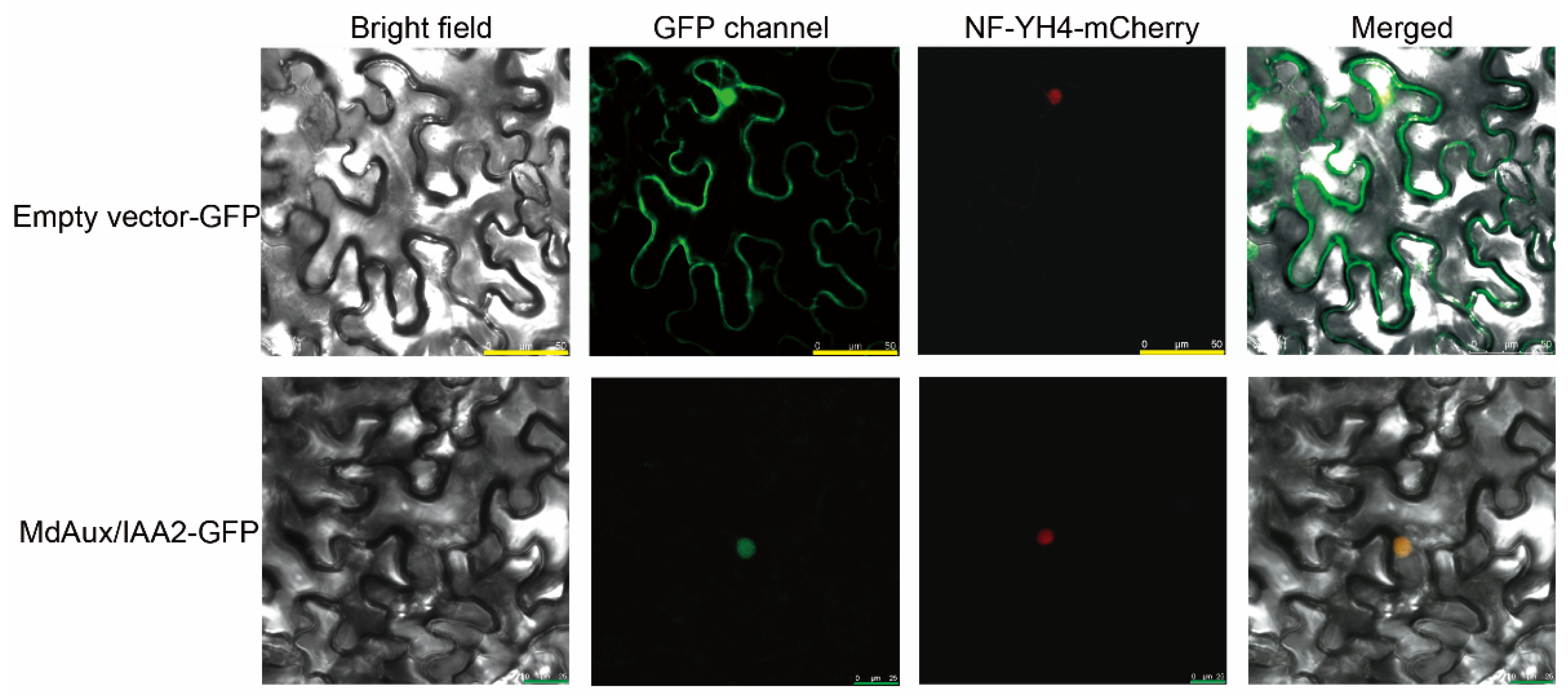
2.3. MdAux/IAA2 Is a Nuclear Location Protein
2.4. Overexpression of MdAux/IAA2 Decreases the Fruit Flesh Callus
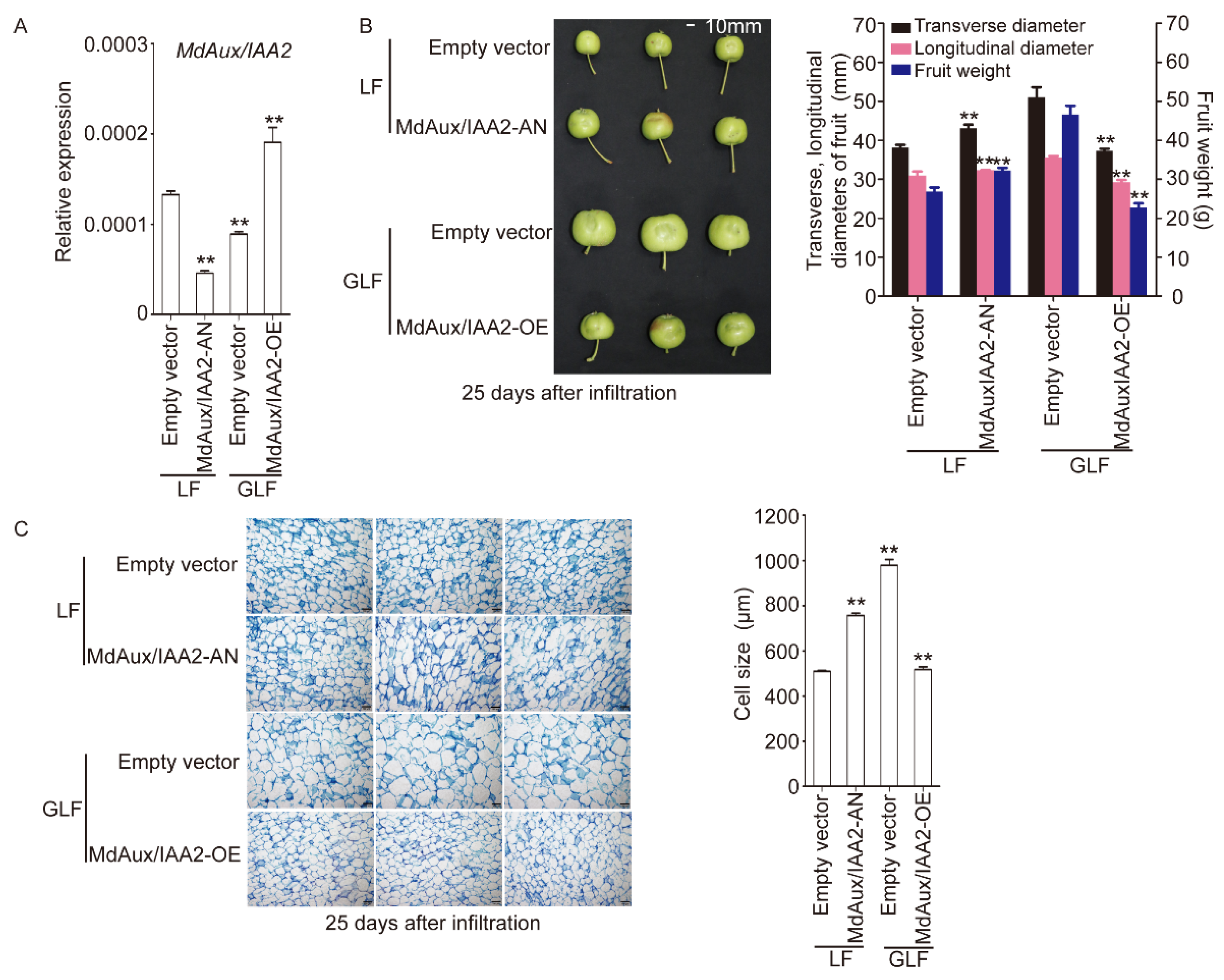
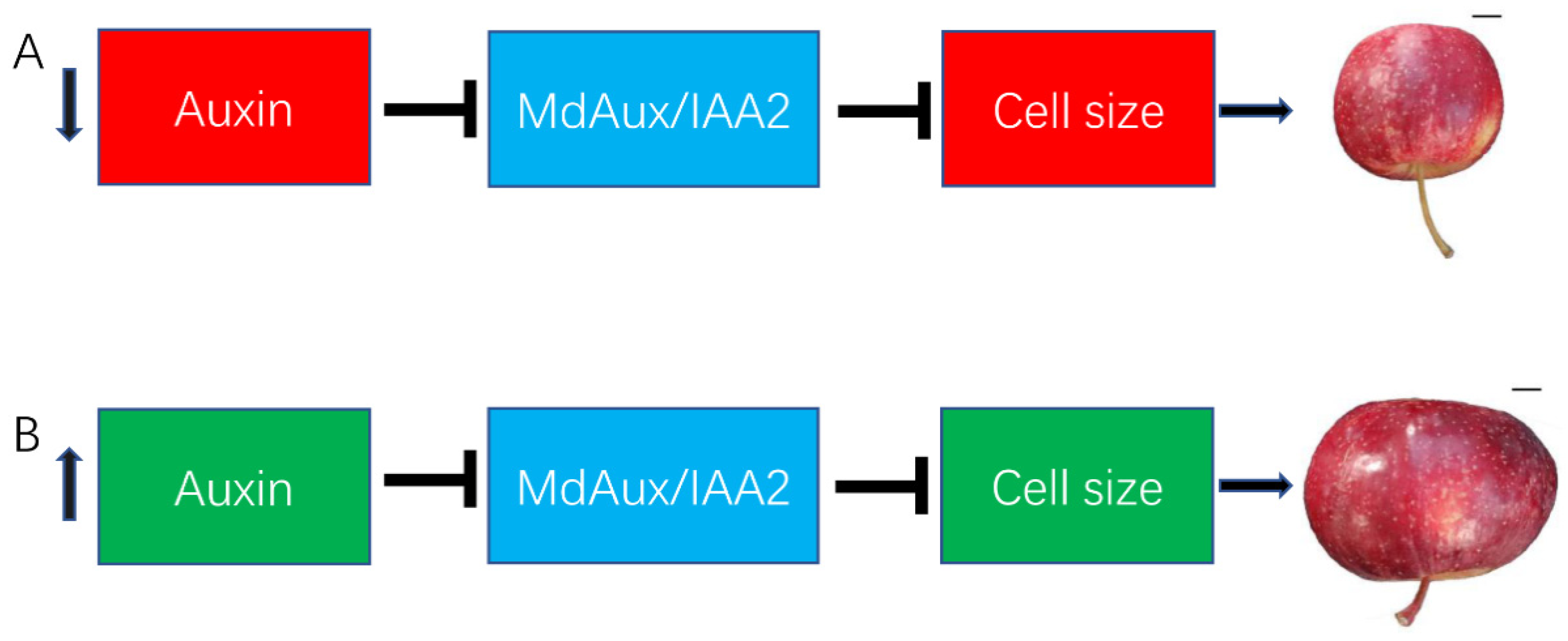
2.5. MdAux/IAA2 Negatively Regulated Apple Fruit Size
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatment
4.2. Gene Cloning and Bioinformatics Analysis
4.3. Vector Construction of MdAux/IAA2
4.4. Callus and Fruit Transformation
4.5. Bioinformatics Analysis
4.6. Statisticical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mészáros, M.; Hnátková, H.; Čonka, P.; Náměstek, J. Linking mineral nutrition and fruit quality to growth intensity and crop load in apple. Agronomy 2021, 11, 506. [Google Scholar] [CrossRef]
- Malladi, A.; Hirst, P.M. Increase in fruit size of a spontaneous mutant of ‘Gala’apple (Malus × domestica Borkh.) is facilitated by altered cell production and enhanced cell size. J. Exp. Bot. 2010, 61, 3003–3013. [Google Scholar] [CrossRef] [Green Version]
- Agusti, M.; Almela, V.; Andreu, I.; Juan, M.; Zacarias, L. Synthetic auxin 3, 5, 6-TPA promotes fruit development and climacteric in Prunus persica L. Batsch. J. Hortic. Sci. Biotechnol. 1999, 74, 556–560. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wismer, P.T.; Proctor, J.; Elfving, D. Benzyladenine affects cell division and cell size during apple fruit thinning. J. Am. Soc. Hortic. Sci. 1995, 120, 802–807. [Google Scholar] [CrossRef] [Green Version]
- IIto, A.; Sakamoto, D.; Itai, A.; Nishijima, T.; Oyama-Okubo, N.; Nakamura, Y.; Moriguchi, T.; Nakajima, I. Effects of GA3+4 and GA4+7 Application Either Alone or Combined with Prohexadione-Ca on Fruit Development of Japanese Pear ‘Kosui’. Hortic. J. 2016, 85, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.; Brown, D.; Nelson, M. Apple shape changing possible with cytokinin and gibbereilin sprays. Calif. Agric. 1970, 24, 14. [Google Scholar]
- Devoghalaere, F.; Doucen, T.; Guitton, B.; Keeling, J.; Payne, W.; Ling, T.J.; Ross, J.J.; Hallett, I.C.; Gunaseelan, K.; Dayatilake, G. A genomics approach to understanding the role of auxin in apple (Malus x domestica) fruit size control. BMC Plant Biol. 2012, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Di, D.; Wu, L.; Yang, J.; Lu, Y.; Shi, W. MicroRNAs Are Involved in Regulating Plant Development and Stress Response through Fine-Tuning of TIR1/AFB-Dependent Auxin Signaling. Int. J. Mol. Sci. 2022, 23, 510. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, F.; Friml, J.; Ding, Z. Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 2022, 64, 371–392. [Google Scholar] [CrossRef]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef]
- Pan, L.; Wang, M.; Yang, Y.; Chen, C.; Dai, H.; Zhang, Z.; Hua, B.; Miao, M. Whole-genome resequencing identified QTLs, candidate genes and Kompetitive Allele-Specific PCR markers associated with the large fruit of Atlantic Giant (Cucurbita maxima). Front. Plant Sci. 2022, 13, 942004. [Google Scholar] [CrossRef] [PubMed]
- Paponov, I.A.; Paponov, M.; Teale, W.; Menges, M.; Chakrabortee, S.; Murray, J.A.; Palme, K. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Dreher, K.A.; Brown, J.; Saw, R.E.; Callis, J. The Arabidopsis Aux/IAA protein family has diversified in degradation and auxin responsiveness. Plant Cell 2006, 18, 699–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, G.; Estelle, M. Auxin receptors: A new role for F-box proteins. Curr. Opin. Cell Biol. 2006, 18, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Cancé, C.; Martin-Arevalillo, R.; Boubekeur, K.; Dumas, R. ARFs are keys to the many auxin doors. New Phytol. 2022, 235, 402–419. [Google Scholar] [CrossRef]
- Lechner, E.; Achard, P.; Vansiri, A.; Potuschak, T.; Genschik, P. F-box proteins everywhere. Curr. Opin. Plant Biol. 2006, 9, 631–638. [Google Scholar] [CrossRef]
- Deng, W.; Yang, Y.; Ren, Z.; Audran-Delalande, C.; Mila, I.; Wang, X.; Song, H.; Hu, Y.; Bouzayen, M.; Li, Z. The tomato SlIAA15 is involved in trichome formation and axillary shoot development. New Phytol. 2012, 194, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jones, B.; Li, Z.; Frasse, P.; Delalande, C.; Regad, F.; Chaabouni, S.; Latche, A.; Pech, J.-C.; Bouzayen, M. The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis. Plant Cell 2005, 17, 2676–2692. [Google Scholar] [CrossRef] [Green Version]
- Bassa, C.; Mila, I.; Bouzayen, M.; Audran-Delalande, C. Phenotypes associated with down-regulation of Sl-IAA27 support functional diversity among Aux/IAA family members in tomato. Plant Cell Physiol. 2012, 53, 1583–1595. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Bassa, C.; Audran, C.; Mila, I.; Cheniclet, C.; Chevalier, C.; Bouzayen, M.; Roustan, J.-P.; Chervin, C. The auxin Sl-IAA17 transcriptional repressor controls fruit size via the regulation of endoreduplication-related cell expansion. Plant Cell Physiol. 2014, 55, 1969–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.-j.; Chen, J.-y.; Lu, W.-j. Expression and regulation of the early auxin-responsive Aux/IAA genes during strawberry fruit development. Mol. Biol. Rep. 2011, 38, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Yu, W.; Yuan, H.; Yue, P.; Wei, Y.; Wang, A. Endogenous Auxin Content Contributes to Larger Size of Apple Fruit. Front. Plant Sci. 2020, 11, 592540. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Harter, K.; Theologis, A. Protein–protein interactions among the Aux/IAA proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 11786–11791. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Liu, Y.; Wang, Y.; Li, H.; Liu, J.; Tan, J.; He, J.; Bai, J.; Ma, H. Evolution analysis of the Aux/IAA gene family in plants shows dual origins and variable nuclear localization signals. Int. J. Mol. Sci. 2017, 18, 2107. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Zhang, B.; Tian, J.-R.; Chen, M.-M.; Zhang, Y.-Y.; Zhang, Z.-H.; Ma, Y. Comparison of the morphology, growth and development of diploid and autotetraploid ‘Hanfu’apple trees. Sci. Hortic. 2017, 225, 277–285. [Google Scholar] [CrossRef]
- Szilágyi, S.; Horváth-Kupi, T.; Desiderio, F.; Bekefi, Z. Evaluation of sweet cherry (Prunus avium L.) cultivars for fruit size by FW_G2a QTL analysis and phenotypic characterization. Sci. Hortic. 2022, 292, 110656. [Google Scholar] [CrossRef]
- Perrot-Rechenmann, C.; Napier, R.M. Auxins. Vitam. Horm. 2005, 72, 203–233. [Google Scholar] [CrossRef]
- Chang, Y.; Sun, R.; Sun, H.; Zhao, Y.; Han, Y.; Chen, D.; Wang, Y.; Zhang, X.; Han, Z. Mapping of quantitative trait loci corroborates independent genetic control of apple size and shape. Sci. Hortic. 2014, 174, 126–132. [Google Scholar] [CrossRef]
- Brown, A. The inheritance of shape, size and season of ripening in progenies of the cultivated apple. Euphytica 1960, 9, 327–337. [Google Scholar] [CrossRef]
- Weiste, C.; Pedrotti, L.; Selvanayagam, J.; Muralidhara, P.; Fröschel, C.; Novák, O.; Ljung, K.; Hanson, J.; Dröge-Laser, W. The Arabidopsis bZIP11 transcription factor links low-energy signalling to auxin-mediated control of primary root growth. PLoS Genet. 2017, 13, e1006607. [Google Scholar] [CrossRef]
- Yue, P.; Lu, Q.; Liu, Z.; Lv, T.; Li, X.; Bu, H.; Liu, W.; Xu, Y.; Yuan, H.; Wang, A. Auxin-activated MdARF5 induces the expression of ethylene biosynthetic genes to initiate apple fruit ripening. New Phytol. 2020, 226, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alayón-Luaces, P.; Pagano, E.A.; Mroginski, L.A.; Sozzi, G.O. Four glycoside hydrolases are differentially modulated by auxins, cytokinins, abscisic acid and gibberellic acid in apple fruit callus cultures. Plant Cell Tissue Organ Cult. 2008, 95, 257–263. [Google Scholar] [CrossRef]
- An, J.-P.; Qu, F.-J.; Yao, J.-F.; Wang, X.-N.; You, C.-X.; Wang, X.-F.; Hao, Y.-J. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple. Hortic. Res. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | FDR (False Discovery Rate) | Log2FC | Relative Expression Mode |
|---|---|---|---|---|
| MD13G1083200 | PREDICTED: transcription factor bHLH79-like [Malus domestica] | 5.35948 × 10−22 | −2.471969519 | down |
| MD15G1377800 | BHLH domain class transcription factor [Malus domestica] | 1.05255 × 10−11 | −1.505775683 | down |
| MD02G1009200 | PREDICTED: transcription factor bHLH66-like [Malus domestica] | 8.88007 × 10−12 | −1.096337386 | down |
| MD10G1156100 | PREDICTED: transcription factor bHLH62 [Malus domestica] | 2.90976 × 10−14 | −2.156869865 | down |
| MD11G1173100 | PREDICTED: transcription factor LHW-like [Pyrus × bretschneideri] | 2.90818 × 10−5 | −1.014574381 | down |
| MD11G1230700 | PREDICTED: transcription factor bHLH123-like isoform X1 [Malus domestica] | 0.001677345 | −1.383493476 | down |
| MD07G1192000 | PREDICTED: transcription factor bHLH111-like isoform X2 [Malus domestica] | 3.31608 × 10−6 | −1.349332523 | down |
| MD12G1112400 | PREDICTED: transcription factor bHLH36-like isoform X3 [Malus domestica] | 0.001632799 | −1.674088612 | down |
| MD15G1023100 | PREDICTED: transcription factor bHLH110-like isoform X1 [Malus domestica] | 3.28979 × 10−7 | −1.074724796 | down |
| MD08G1026300 | PREDICTED: transcription factor bHLH110 isoform X1 [Malus domestica] | 3.17243 × 10−5 | −1.456370629 | down |
| Gene ID | Gene Name | FDR (False Discovery Rate) | Log2FC | Relative Expression Mode |
|---|---|---|---|---|
| MD06G1217200 | PREDICTED: transcription factor WER [Malus domestica] | 9.53635 × 10−18 | 1.005209081 | up |
| MD16G1228600 | PREDICTED: transcription factor MYB21-like [Pyrus × bretschneideri] | 2.60867 × 10−5 | 1.248571648 | up |
| MD08G1031200 | PREDICTED: protein RADIALIS-like 3 [Malus domestica] | 1.42606 × 10−5 | 2.575760236 | up |
| MD02G1308300 | PREDICTED: peptidyl-prolyl cis-trans isomerase FKBP42-like isoform X2 [Malus domestica] | 1.63394 × 10−7 | 2.073944629 | up |
| MD11G1104500 | PREDICTED: uncharacterized protein LOC108172356 [Malus domestica] | 7.86998 × 10−9 | 3.492959082 | up |
| MD14G1172900 | PREDICTED: transcription factor MYB86-like [Pyrus × bretschneideri] | 0.006193873 | −1.831702439 | down |
| MD16G1148300 | PREDICTED: transcription factor MYB108-like [Malus domestica] | 8.63823 × 10−7 | −3.028824041 | down |
| MD04G1184900 | MYB domain class transcription factor [Malus domestica] | 0.006696246 | −1.001992516 | down |
| MD02G1087900 | PREDICTED: transcription factor TT2 [Pyrus × bretschneideri] | 0.009579611 | −1.076780594 | down |
| MD14G1222200 | PREDICTED: transcription factor DIVARICATA-like [Pyrus × bretschneideri] | 0.000154883 | −1.155980792 | down |
| MD15G1025600 | PREDICTED: protein RADIALIS-like 6 [Malus domestica] | 5.1832 × 10−5 | −1.671371068 | down |
| MD16G1083300 | PREDICTED: transcription factor MYB1R1 [Malus domestica] | 3.09699 × 10−5 | −1.155418453 | down |
| MD05G1341500 | PREDICTED: MYB-like transcription factor ETC1 [Malus domestica] | 4.23716 × 10−5 | −1.340022381 | down |
| MD10G1216700 | PREDICTED: myb family transcription factor APL-like [Pyrus × bretschneideri] | 0.000537588 | −1.042637231 | down |
| MD11G1173100 | PREDICTED: transcription factor LHW-like [Pyrus × bretschneideri] | 2.90818 × 10−5 | −1.014574381 | down |
| Gene ID | Gene Name | FDR (False Discovery Rate) | Log2FC | Relative Expression Mode |
|---|---|---|---|---|
| MD08G1123300 | PREDICTED: basic leucine zipper 9-like [Malus domestica] | 6.47336 × 10−12 | 1.431449022 | up |
| MD03G1051900 | PREDICTED: transcription factor RF2b-like [Malus domestica] | 1.18702 × 10−9 | 1.264167539 | up |
| MD02G1189300 | PREDICTED: basic leucine zipper 8-like [Malus domestica] | 2.35816 × 10−5 | 1.127923266 | up |
| MD08G1123300 | PREDICTED: basic leucine zipper 9-like [Malus domestica] | 6.47336 × 10−12 | 1.431449022 | up |
| MD03G1051900 | PREDICTED: transcription factor RF2b-like [Malus domestica] | 1.18702 × 10−9 | 1.264167539 | up |
| MD02G1189300 | PREDICTED: basic leucine zipper 8-like [Malus domestica] | 2.35816 × 10−5 | 1.127923266 | up |
| Assay | Primer Name | Sequence (5′–3′) |
|---|---|---|
| Gene expression | MdAux/IAA2-ex-F | GAACTTGTTGTGGTGTGGAG |
| MdAux/IAA2-ex-R | CGCTGATTGCTACTAAGCTG | |
| MdActin-F | GGCTGGATTTGCTGGTGATG | |
| MdActin-R | TGCTCACTATGCCGTGCTCA | |
| Gene cloning | MdAux/IAA2-full-F | ATGGATAATTTGTATAGTATGAT |
| MdAux/IAA2-full-R | TCATCTTCCAATTCCAACTT | |
| MdAux/IAA2-pro-F | AGATAGCTAAGAATTTCTGGCGTA | |
| MdAux/IAA2-pro-R | GCCCTTACAACTCCATTATTCTCA | |
| Vector construction | pRI101-MdAux/IAA2-OE-F | TCTTCACTGTTGATACATATGATGGATAATTTGTATAGTATGAT |
| pRI101-MdAux/IAA2-OE-R | CGATCGGGGAAATTCGAGCTCTCATCTTCCAATTCCAACTT | |
| pRI101-MdAux/IAA2-AN-F | CGATCGGGGAAATTCGAGCTCATGGACGGGAAGAAACTGGA | |
| pRI101-MdAux/IAA2-AN-R | TCTTCACTGTTGATACATATGCTAAGGGATCCACTTAGGATCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, H.; Sun, X.; Yue, P.; Qiao, J.; Sun, J.; Wang, A.; Yuan, H.; Yu, W. The MdAux/IAA2 Transcription Repressor Regulates Cell and Fruit Size in Apple Fruit. Int. J. Mol. Sci. 2022, 23, 9454. https://doi.org/10.3390/ijms23169454
Bu H, Sun X, Yue P, Qiao J, Sun J, Wang A, Yuan H, Yu W. The MdAux/IAA2 Transcription Repressor Regulates Cell and Fruit Size in Apple Fruit. International Journal of Molecular Sciences. 2022; 23(16):9454. https://doi.org/10.3390/ijms23169454
Chicago/Turabian StyleBu, Haidong, Xiaohuan Sun, Pengtao Yue, Junling Qiao, Jiamao Sun, Aide Wang, Hui Yuan, and Wenquan Yu. 2022. "The MdAux/IAA2 Transcription Repressor Regulates Cell and Fruit Size in Apple Fruit" International Journal of Molecular Sciences 23, no. 16: 9454. https://doi.org/10.3390/ijms23169454