
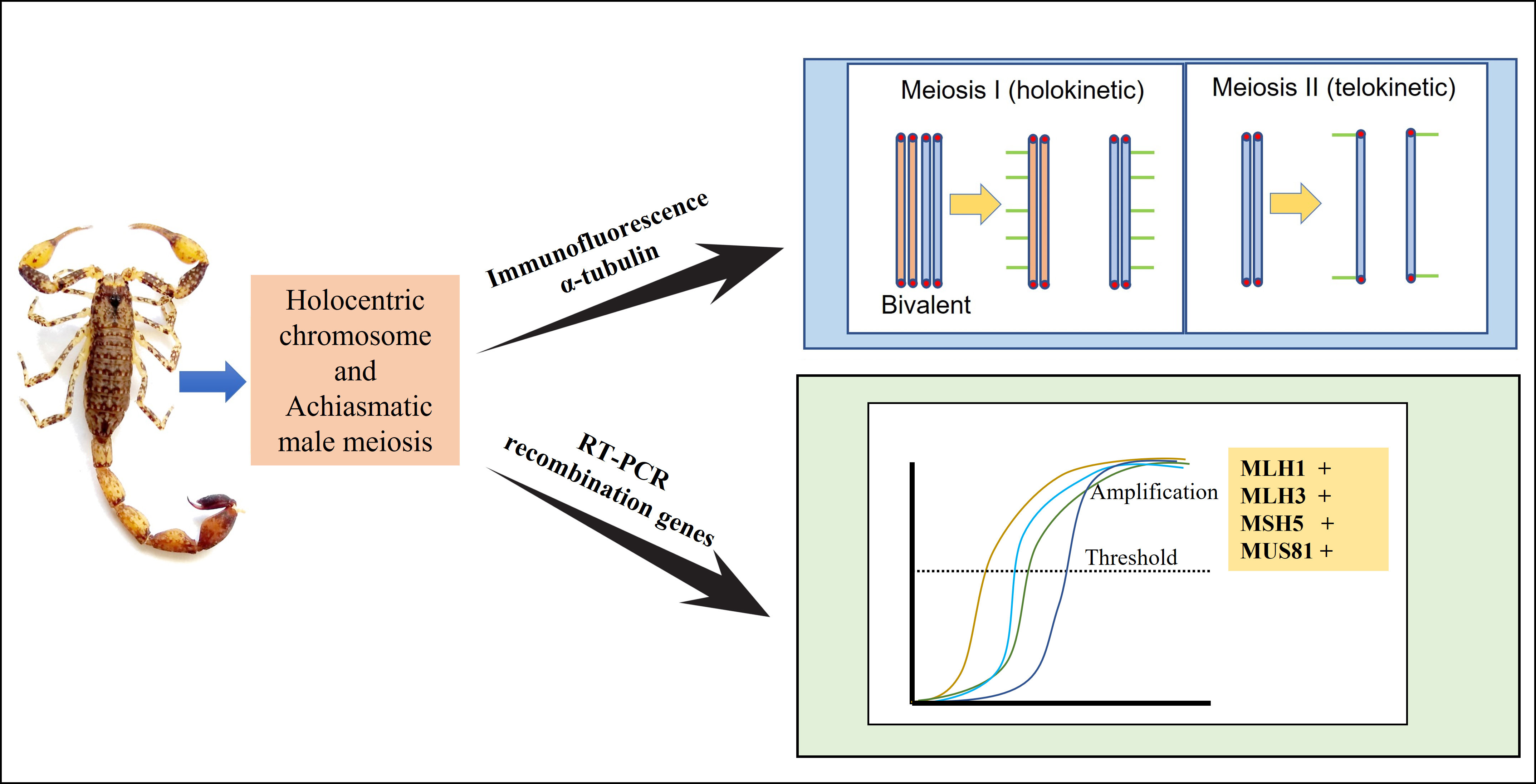
Kinetic Activity of Chromosomes and Expression of Recombination Genes in Achiasmatic Meiosis of Tityus (Archaeotityus) Scorpions

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Results
2.1. Karyotype and Synaptonemal Complex Formation in T. maranhensis
2.2. Distribution of γH2AX throughout Prophase I of T. maranhensis
2.3. Binding of Microtubules along the Chromosomes of T. maranhensis
2.4. Expression of Mismatch Repair Enzymes in T. silvestris
3. Discussion
4. Materials and Methods
4.1. Karyotype Analysis
4.2. Probes
4.3. Immunodetection of Meiotic Proteins
4.4. Immuno-FISH
4.5. RT-PCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloom, K.; Costanzo, V. Centromere Structure and Function. Prog. Mol. 2017, 56, 515–539. [Google Scholar]
- Melters, D.P.; Paliulis, L.V.; Korf, I.F.; Chan, S.W.F. Holocentric chromosome: Convergent evolution, meiotic adaptations, and genomic analysis. Chromosome Res. 2012, 20, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Mola, L.A.; Papeschi, A.G. Holokinetic chromosome an glance. BAG J. Basic Appl. Genet. 2006, 17, 17–33. [Google Scholar]
- Zedek, F.; Bureš, P. Holocentric chromosomes: From tolerance to fragmentation to colonization of the land. Ann. Bot. 2017, 121, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.C.; Zacaro, A.A.; Pinto-Da-Rocha, R.; Candido, D.M.; Cella, D.M. Complex meiotic configuration of the holocentric chromosomes: The intriguing case of the scorpion Tityus bahiensis. Chromosome Res. 2009, 17, 883–898. [Google Scholar] [CrossRef]
- Mattos, V.F.; Carvalho, L.S.; Carvalho, M.A.; Schneider, M.C. Insights into the origin of the high variability of multivalent-meiotic associations in holocentric chromosomes of Tityus (Archaeotityus) scorpions. PLoS ONE 2018, 13, e0192070. [Google Scholar] [CrossRef]
- Adilardi, R.S.; Ojanguren-Affilastro, A.A.; Martí, D.A.; Mola, L.M. Chromosome puzzle in the southernmost populations of the medically important scorpion Tityus bahiensis (Perty 1833) (Buthidae), a polymorphic species with striking structural rearrangements. Zoo. Anz. 2020, 288, 139–150. [Google Scholar] [CrossRef]
- Shanahan, C.M. Cytogenetics of Australian scorpions I. Interchange polymorphism in the family Buthidae. Genome 1989, 32, 882–889. [Google Scholar]
- Mattos, V.F.; Cella, D.M.; Carvalho, L.S.; Candido, D.M.; Schneider, M.C. High chromosome variability and the presence of multivalent associations in buthid scorpions. Chromosome Res. 2013, 21, 121–136. [Google Scholar] [CrossRef]
- Schneider, M.C.; Mattos, V.F.; Carvalho, L.S.; Cella, D.M. Organization and behavior of the synaptonemal complex during achiasmatic meiosis of four buthid scorpions. Cytogenet Genome Res. 2015, 144, 341–347. [Google Scholar] [CrossRef]
- Almeida, B.R.R.; Milhomem-Paixão, S.S.R.; Noronha, R.C.R.; Nagamachi, C.Y.; Costa, M.J.R.; Pardal, P.P.O.; Coelho, J.S.; Pieczarka, J.C. Karyotype diversity and chromosomal organization of repetitive DNA in Tityus obscurus (Scorpiones, Buhidae). BMC Genet. 2017, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Adilardi, R.S.; Ojanguren-Affilastro, A.A.; Mola, L.M. Sex-linked chromosome heterozygosity in males of Tityus confluens (Buthidae): A clue about the presence of sex chromosomes in scorpions. PLoS ONE 2016, 11, e0164427. [Google Scholar] [CrossRef] [PubMed]
- Almeida, B.R.R.; Noronha, R.C.R.; Costa, M.J.R.; Nagamachi, C.Y.; Pieczarka, J.C. Meiosis in the scorpion Tityus silvestris: New insights into achiasmatic chromosomes. Biol. Open 2019, 8, bio04352. [Google Scholar] [CrossRef] [PubMed]
- Manterola, M.; Page, J.; Vasco, C.; Berríos, S.; Parra, M.T.; Viera, A.; Rufas, J.S.; Zuccotti, M.; Garagna, S.; Fernández-Donoso, R. A high incidence of meiotic silencing of unsynapsed chromatin is not associated with substantial pachytene loss in heterozygous male mice carrying multiple simple robertsonian translocations. PLoS Genet. 2009, 5, e1000625. [Google Scholar] [CrossRef] [PubMed]
- Mary, N.; Calgaro, A.; Barasc, H.; Bonnet, N.; Ferchaud, S.; Raymond-Letron, I.; Ducos, A.; Pinton, A. Meiotic Silencing in Pigs: A Case Study in a Translocated Azoospermic Boar. Genes 2021, 12, 1137. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, S.; Wassenaar, E.; Laven, J.S.; Grootegoed, J.A.; Baarends, W.M. Meiotic silencing and fragmentation of the male germline restricted chromosome in zebra finch. Chromosoma 2010, 119, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Noronha, R.C.R.; Almeida, B.R.R.; Costa, M.J.R.; Nagamachi, C.Y.; Martins, C.; Pieczarka, J.C. Meiotic analyses show adaptations to maintenance of fertility in X1Y1 × 2Y2 × 3Y3 × 4Y4 × 5Y5 system of amazon frog Leptodactylus pentadactylus (Laurenti, 1768). Sci. Rep. 2020, 10, 16327. [Google Scholar] [CrossRef]
- Viera, A.; Parra, M.T.; Rufas, J.S.; Page, J. Transcription reactivation during the first meiotic prophase in bugs is not dependent on synapsis. Chromosoma 2017, 126, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.M.; Mahadevaiah, S.K.; Fernandez-Capetillo, O.; Nussenzweig, A.; Xu, X.; Deng, C.X.; Burgoyne, P.S. Silencing of unsynapsed meiotic chromosomes in the mouse. Nat. Genet. 2005, 37, 41–47. [Google Scholar] [CrossRef]
- Marques, A.; Pedrosa-Harand, A. Holocentromere identity: From the typical mitotic linear structure to the great plasticity of meiotic holocentromeres. Chromosoma 2016, 125, 669–681. [Google Scholar] [CrossRef]
- Heckmann, S.; Jankowska, M.; Schubert, V.; Kumke, K.; Ma, W.; Houben, A. Alternative meiotic chromatid segregation in the holocentric plant Luzula elegans. Nat. Commun 2014, 5, 4979. [Google Scholar] [CrossRef] [PubMed]
- Wignall, S.; Villeneuve, A. Lateral microtubule bundles promote chromosome alignment during acentrosomal oocyte meiosis. Nat. Cell Biol. 2009, 11, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Pérez, R.; Rufas, J.S.; Suja, J.A.; Page, J.; Panzera, F. Meiosis in holocentric chromosomes: Orientation and segregation of an autosome and sex chromosomes in Triatoma infestans (Heteroptera). Chromosome Res. 2000, 8, 17–25. [Google Scholar] [CrossRef]
- Kazuhiro, S.; Naoki, O.; Toshinori, E. Achiasmy and sex chromosome evolution. Ecol. Genet. Genom. 2019, 13, 100046. [Google Scholar]
- Adilardi, R.S.; Ojanguren-Affilastro, A.A.; Mattoni, C.I.; Mola, L.M. Male and female meiosis in the mountain scorpion Zabius fuscus (Scorpiones, Buthidae): Heterochromatin, rDNA and TTAGG telomeric repeats. Genetica 2015, 143, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Ubinski, C.V.; Carvalho, L.S.; Schneider, M.C. Mechanisms of karyotype evolution in the Brazilian scorpions of the subfamily Centruroidinae (Buthidae). Genetica 2018, 146, 475–486. [Google Scholar] [CrossRef]
- John, A.; Vinayan, K.; Varghese, J. Achiasmy: Male fruit flies are not ready to mix. Front. Cell Dev. Biol. 2016, 4, 75. [Google Scholar] [CrossRef]
- Moens, P.B.; Marcon, E.; Shore, J.S.; Kochakpour, N.; Spyropoulos, B. Initiation and resolution of interhomolog connections: crossover and non-crossover sites along mouse synaptonemal complexes. J. Cell Sci. 2007, 120, 1017–1027. [Google Scholar] [CrossRef]
- Arana, P.; Santos, J.L.; Henriques-Gil, N. Interference relationships in grasshopper reciprocal translocation heterozygotes. Heredity 1987, 59, 85–93. [Google Scholar] [CrossRef]
- Oliver-Bonet, M.; Benet, J.; Sun, F.; Navarro, J.; Abad, C.; Liehr, T.; Starkes, H.; Greene, C.; Ko, E.; Martin, R.H. Meiotic studies in two human reciprocal translocations and their association with spermatogenic failure. Hum. Reprod. 2005, 20, 683–688. [Google Scholar] [CrossRef]
- Ribagorda, M.; Berríos, S.; Solano, E.; Ayarza, E.; Martín-Ruiz, M.; Gil-Fernández, A.; Parra, M.T.; Viera, A.; Rufas, J.S.; Capanna, E.; et al. Meiotic behavior of a complex hexavalent in heterozygous mice for Robertsonian translocations: Insights for synapsis dynamics. Chromosoma 2019, 128, 149–163. [Google Scholar] [CrossRef]
- Berríos, S.; Fernández-Donoso, R.; Ayarza, E. Synaptic configuration of quadrivalents and their association with the XY bivalent in spermatocytes of Robertsonian heterozygotes of Mus domesticus. Biol. Res. 2017, 50, 38. [Google Scholar] [CrossRef] [PubMed]
- Checchi, P.M.; Engebrecht, J. Caenorhabditis elegans histone methyltransferase MET-2 shields the male X chromosome from checkpoint machinery and mediates meiotic sex chromosome inactivation. PLoS Genet. 2011, 7, e1002267. [Google Scholar] [CrossRef] [PubMed]
- Mahadevaiah, S.K.; Bourc’his, D.; DE Rooij, D.G.; Bestor, T.H.; Turner, J.M.; Burgoyne, P.S. Extensive meiotic asynapsis in mice antagonises meiotic silencing of unsynapsed chromatin and consequently disrupts meiotic sex chromosome inactivation. J. Cell Biol. 2008, 182, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cruz, R.; Roig, I.; Robles, P.; Scherthan, H.; Garcia Caldés, M. ATR, BRCA1 and gammaH2AX localize to unsynapsed chromosomes at the pachytene stage in human oocytes. Reprod Biomed. Online 2009, 18, 37–44. [Google Scholar] [CrossRef]
- Subramanian, V.V.; Hochwagen, A. The meiotic checkpoint network: Step-by-step through meiotic prophase. Cold Spring Harb Perspect Biol. 2014, 6, a016675. [Google Scholar] [CrossRef] [PubMed]
- Traut, W.; Schubert, V.; Daliková, M.; Marec, F.; Sahara, K. Activity and inactivity of moth sex chromosomes in somatic and meiotic cells. Chromosoma 2019, 128, 533–545. [Google Scholar] [CrossRef]
- Viera, A.; Parra, M.T.; Arévalo, S.; García De La Vega, C.; Santos, J.L.; Page, J. X Chromosome Inactivation during Grasshopper Spermatogenesis. Genes 2021, 12, 1844. [Google Scholar] [CrossRef]
- Lee, C.Y.; Horn, H.F.; Stewart, C.L.; Burke, B.; Bolcun-Filas, E.; Schimenti, J.C.; Dresser, M.E.; Pezza, R.J. Mechanism and regulation of rapid telomere prophase movements in mouse meiotic chromosomes. Cell Rep. 2015, 11, 551–563. [Google Scholar] [CrossRef]
- Claycomb, J.M.; Batista, P.J.; Pang, K.M.; Gu, W.; Vasale, J.J.; Van Wolfswinkel, J.C.; Chaves, D.A.; Shirayama, M.; Mitani, S.; Ketting, R.F.; et al. The Argonaute CSR-1 and its 22G-RNA cofactors are required for holocentric chromosome segregation. Cell 2009, 139, 123–134. [Google Scholar] [CrossRef]
- Bongiorni, S.; Fiorenzo, P.; Pippoletti, D.; Prantera, G. Inverted meiosis and meiotic drive in mealybugs. Chromosoma 2004, 112, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Mandrioli, M.; Manicardi, G.C. Holocentric chromosomes. PLoS Genet. 2020, 16, e1008918. [Google Scholar] [CrossRef]
- Goday, C.; Pimpinelli, S. Centromere organization in meiotic chromosomes of Parascaris univalens. Chromosoma 1989, 98, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Comings, D.E.; Okada, T.A. Holocentric chromosomes in Oncopeltus: Kinetochore plates are present in mitosis but absent in meiosis. Chromosoma 1972, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Drinnenberg, I.A.; Young, D.; Henikoff, S.; Malik, H.S. Recurrent loss of cenH3 is associated with independent transitions to holocentricity in insects. Elife 2014, 3, e03676. [Google Scholar] [CrossRef]
- Benavente, R. Holocentric chromosomes of arachnids: Presence of kinetochore plates during meiotic divisions. Genetica 1982, 59, 23–27. [Google Scholar] [CrossRef]
- Schvarzstein, M.; Wignall, S.M.; Villeneuve, A.M. Coordinating cohesion, co-orientation, and congression during meiosis: Lessons from holocentric chromosomes. Genes Dev. 2010, 24, 219–228. [Google Scholar] [CrossRef]
- Shanahan, C.M.; Hayman, D.L. Synaptonemal complex formation in male scorpions exhibiting achiasmate meiosis and structural heterozygosity. Genome 1990, 33, 914–926. [Google Scholar] [CrossRef]
- Marec, F.; Traut, W. Synaptonemal complexes in female and male meiotic prophase of Ephestia kuehniella (Lepidoptera). Heredity 1993, 71, 394–404. [Google Scholar] [CrossRef]
- Osman, F.; Dixon, J.; Doe, C.L.; Whitby, M.C. Generating Crossovers by Resolution of Nicked Holliday Junctions. Molecular Cell 2003, 12, 761–774. [Google Scholar] [CrossRef]
- Gunes, S.; Al-Sadaan, M.; Agarwal, A. Spermatogenesis, DNA damage and DNA repair mechanisms in male infertility. Reprod. Biomed. Online 2015, 31, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, W.R. Amazonian Arachnida and Miryapoda: Identification Keys to All Classes, Orders, Families, Some Genera an List of Know Terrestrial Species; Pensoft Publishes: Moscow, Russia, 2002; pp. 399–438. [Google Scholar]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef] [PubMed]
- Araya-Jaime, C.; Serrano, É.A.; Andrade Silva, D.M.; Yamashita, M.; Iwai, T.; Oliveira, C.; Foresti, F. Surface-spreading technique of meiotic cells and immunodetection of synaptonemal complex proteins in teleostean fishes. Mol. Cytogenet 2015, 8, 1–6. [Google Scholar] [CrossRef]
- Guo, J.; Wang, L.; Wu, H.; Cao, Y.; Xiao, R.; Lai, X.; Liu, J.; Yi, J.; Zhang, G. Molecular characterization and expression of vitellogenin genes from the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). Physiol. Entomol. 2018, 43, 295–305. [Google Scholar] [CrossRef]
- Simon, P. Q-Gene: Processing quantitative real-time RT-PCR data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Individuals | Collection Location |
|---|---|---|
| Tityus silvestris Pocock, 1897 | 1 male, 1 female, and 4 embryos | Manaus, Amazonas, Brazil (3°06′10″ S/59°58′42″ O) |
| 3 males and 3 females | Belém, Pará, Brazil (1°28′06″ S/48°26′24″ O) | |
| Tityus maranhensis Lourenço de Jesus Junior e Limeira-de-Oliveira, 2006 | 1 male | Marapanim, Pará, Brazil (0°56′12″ S/47°38′39″ O) |
| 1 male and 1 female | Curuçá, Pará, Brazil (0°46′40″ S/47°49′40″ O) |
| Gene | Primers | Tm (°C) | Amplicon Size |
|---|---|---|---|
| MLH1 | F:5′-GATAGCGAGGAGAGTGGATGG-3′ | 59.5 | 88 bp |
| R: 5′-CCGCCTTTGTCTGTAGGAAATC-3′ | 59.3 | ||
| MHL3 | F:5′-CCTCTGCCTTCCGAAGTTGTT-3′ | 60.5 | 118 bp |
| R: 5′-CGCAAACATTCGTAGCAAAAGC-3′ | 59.9 | ||
| MSH5 | F:5′-GCGGGACCTAACGAACATTC-3′ | 59 | 117 bp |
| R: 5′-GATGTCAACGGGCAACTCG-3′ | 59.2 | ||
| MUS81 | F:5′-CGTACCTGGATCGGGAAGC-3′ | 59.9 | 161 bp |
| R:5′-AAGCAGTGTAATGGCTACCAGG-3′ | 60.4 | ||
| β-Actin | F:5′-TGCGGTGGACAATGGAAGG-3′ | 62 °C | 109 bp |
| R:5′-GTCTGGATTGGTGGCTCTATCT-3′ | 60 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, B.R.R.d.; Noronha, R.C.R.; Cardoso, A.L.; Martins, C.; Martins, J.G.; Procópio, R.E.d.L.; Nagamachi, C.Y.; Pieczarka, J.C. Kinetic Activity of Chromosomes and Expression of Recombination Genes in Achiasmatic Meiosis of Tityus (Archaeotityus) Scorpions. Int. J. Mol. Sci. 2022, 23, 9179. https://doi.org/10.3390/ijms23169179
Almeida BRRd, Noronha RCR, Cardoso AL, Martins C, Martins JG, Procópio REdL, Nagamachi CY, Pieczarka JC. Kinetic Activity of Chromosomes and Expression of Recombination Genes in Achiasmatic Meiosis of Tityus (Archaeotityus) Scorpions. International Journal of Molecular Sciences. 2022; 23(16):9179. https://doi.org/10.3390/ijms23169179
Chicago/Turabian StyleAlmeida, Bruno Rafael Ribeiro de, Renata Coelho Rodrigues Noronha, Adauto Lima Cardoso, Cesar Martins, Jonas Gama Martins, Rudi Emerson de Lima Procópio, Cleusa Yoshiko Nagamachi, and Julio Cesar Pieczarka. 2022. "Kinetic Activity of Chromosomes and Expression of Recombination Genes in Achiasmatic Meiosis of Tityus (Archaeotityus) Scorpions" International Journal of Molecular Sciences 23, no. 16: 9179. https://doi.org/10.3390/ijms23169179