
Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
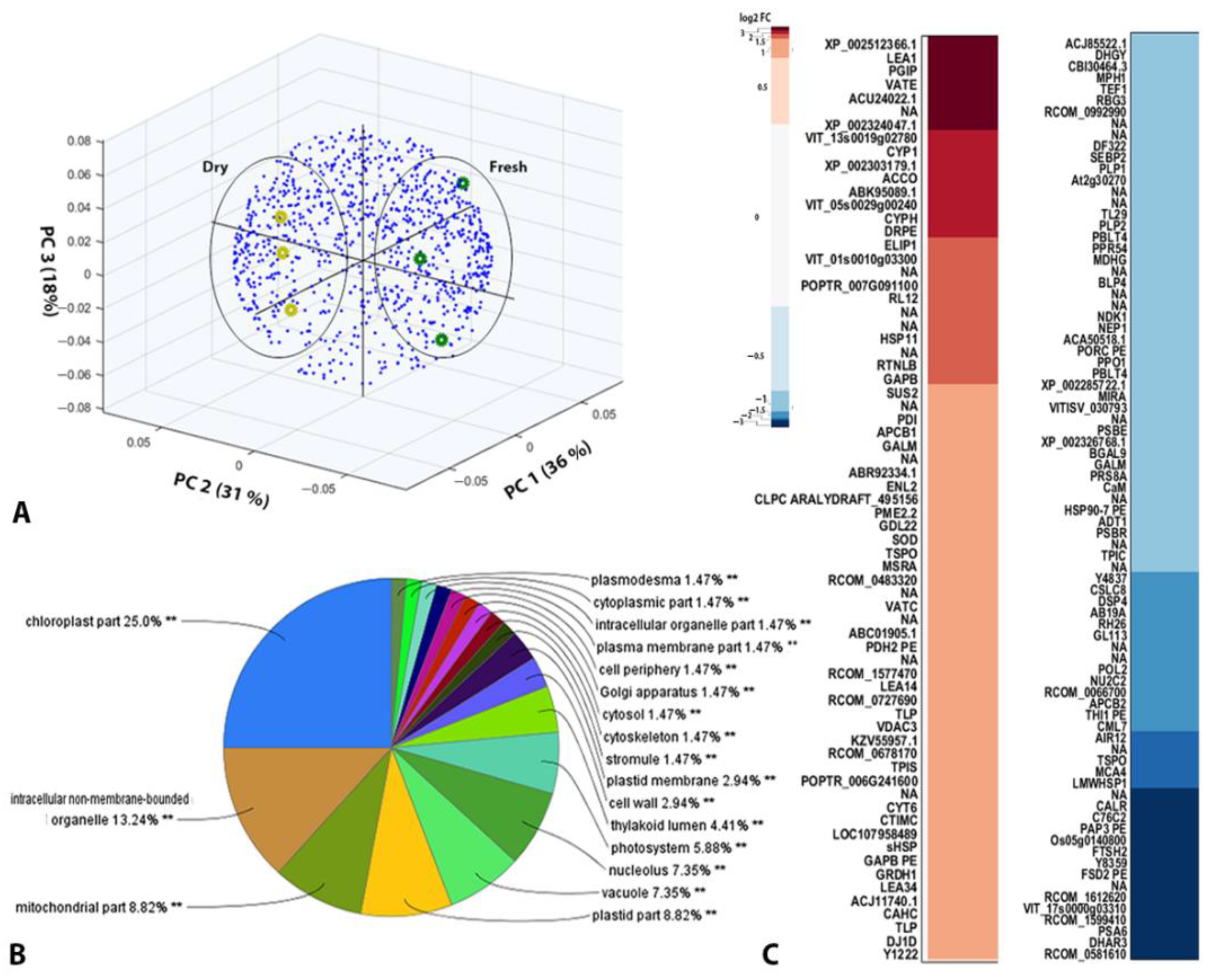
2. Results
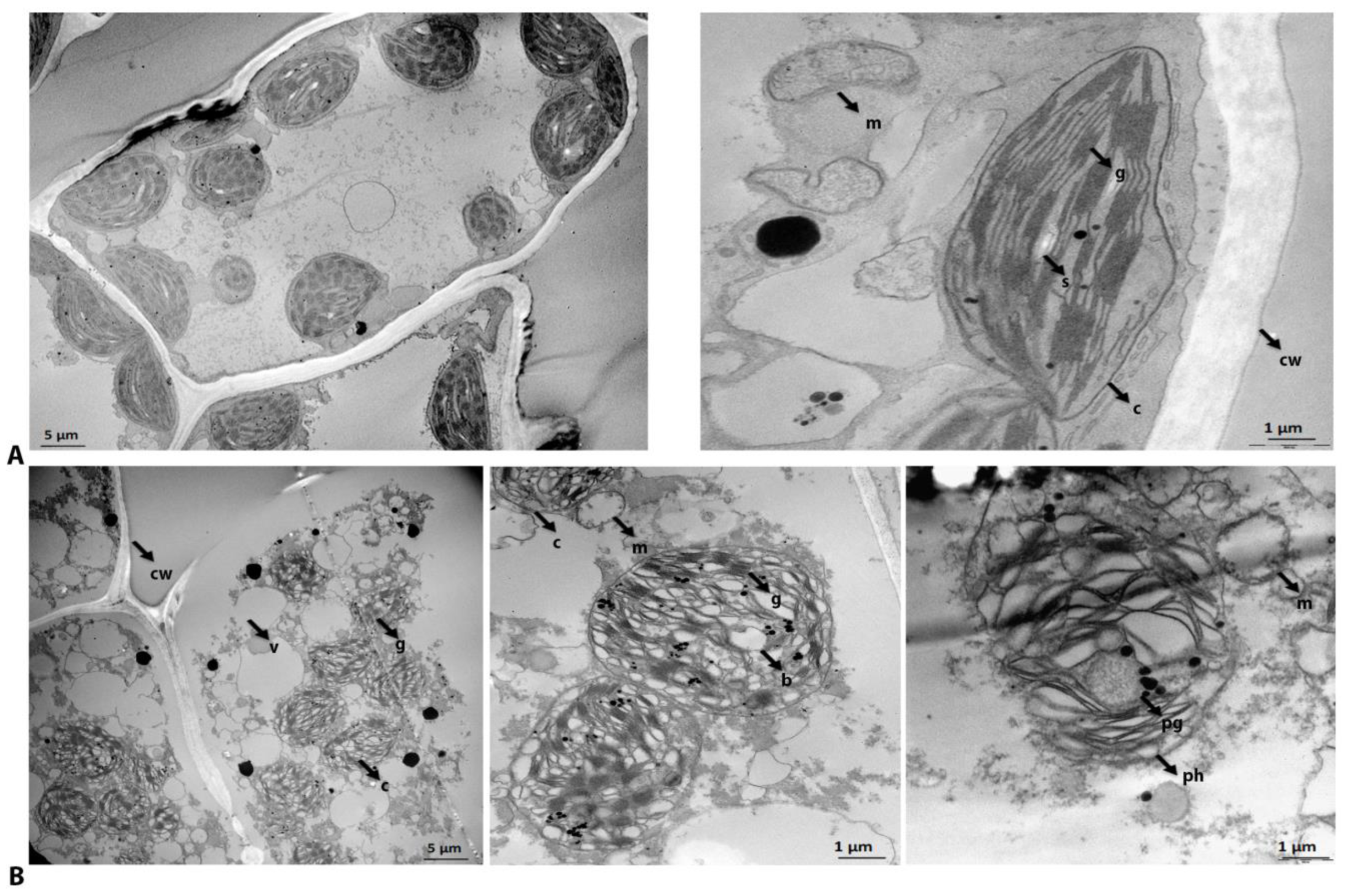
2.1. Cellular and Subcellular Ultrastructure of Fully Hydrated and Dry H. rhodopensis
2.2. Quantitative Proteomic Analysis of Fresh and Dry H. rhodopensis Crude Cell Samples
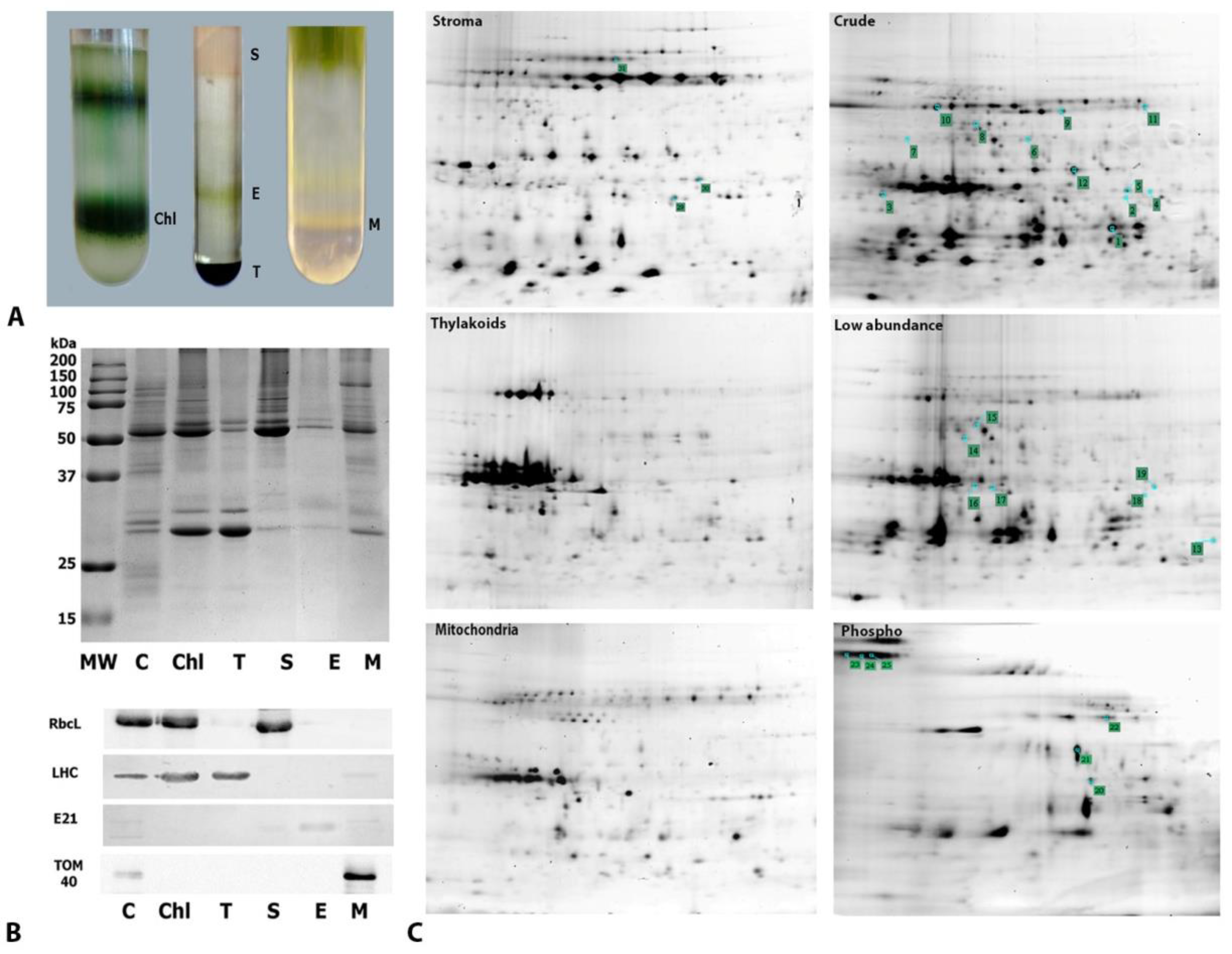
2.3. Subcellular Fractionation and Gel-Based Proteomics of Crude, Organelle, Phosphoprotein, and Low-Abundance Protein Fractions
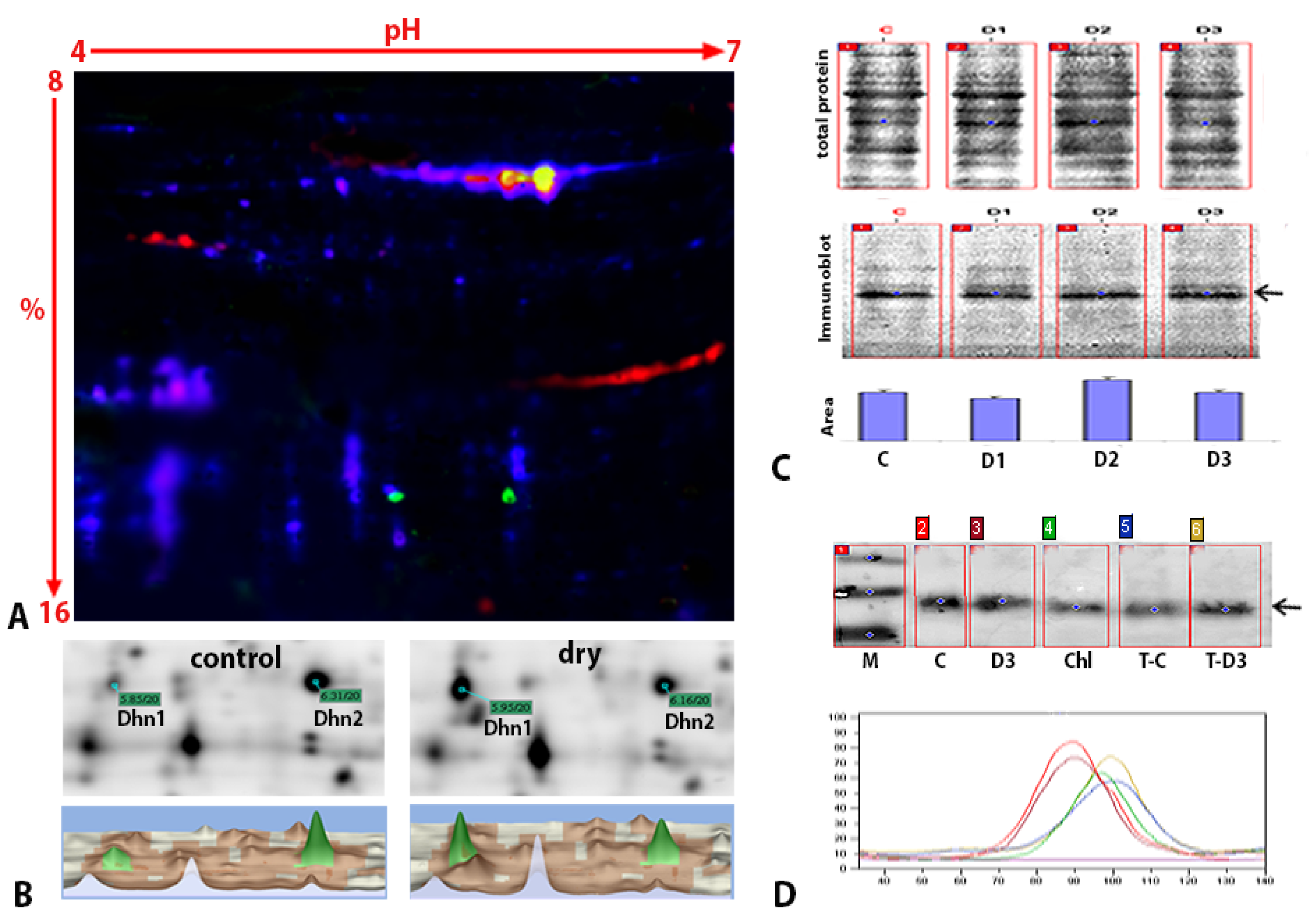
2.4. Expression, Posttranslational Modifications, and Subcellular Localization of Dehydrins
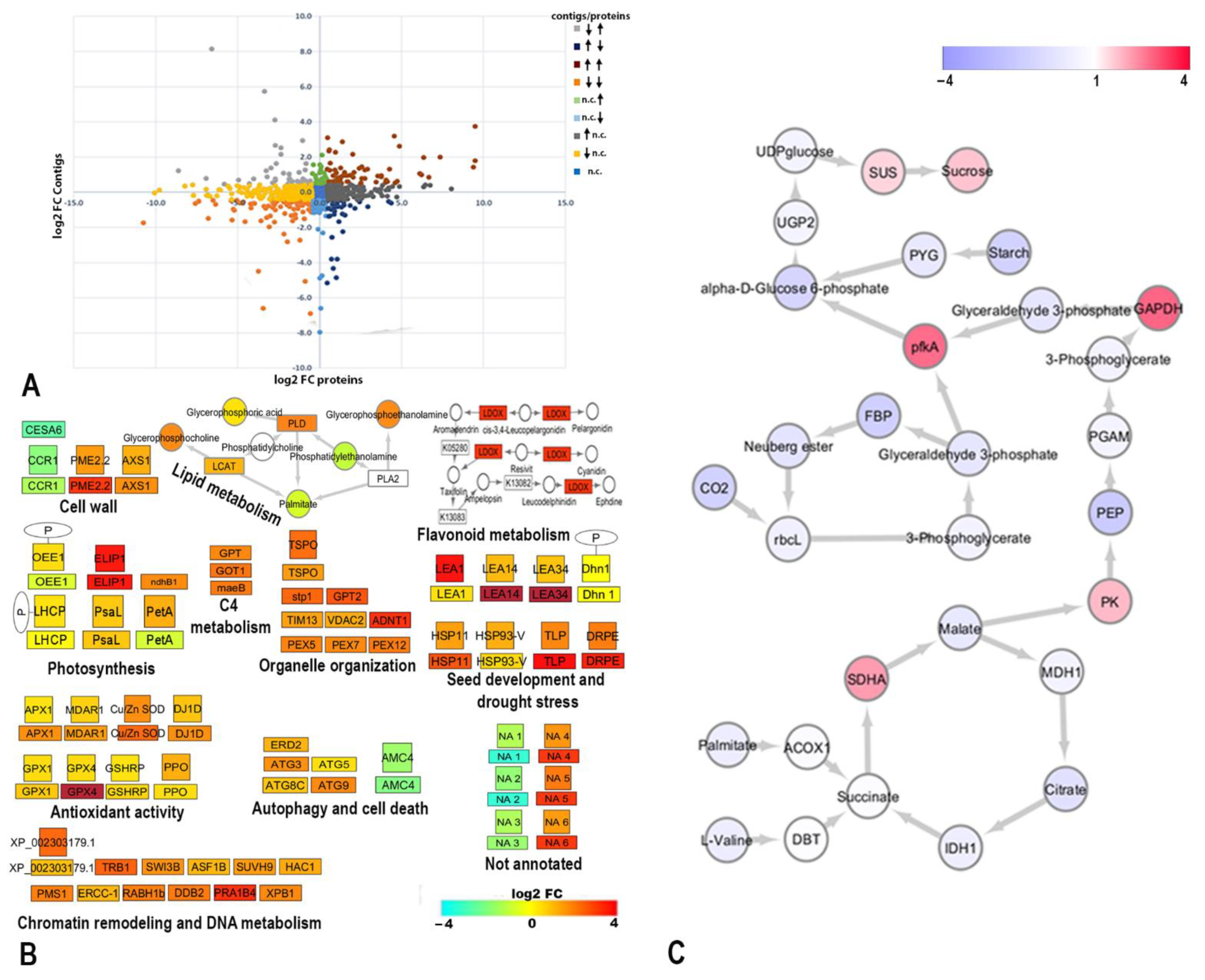
2.5. Pathway Analysis and Co-Expression Changes of the Proteome, Transcriptome, and Metabolomics of H. rhodopensis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Desiccation Stress
4.2. Transmission Electron Microscopy (TEM)
4.3. Shotgun Proteomics
4.4. Purification of Chloroplasts, Mitochondria, and Chloroplast Subcompartments
4.5. Purification of Phosphoproteins and Low-Abundance Enriched Proteins and Non-Targeted Gel-Based Proteomics
4.6. Targeted Gel-Based Proteomics
4.7. Statistics and Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, J.; Ji, M.; Xie, Y.; Wang, S.; He, Y.; Ran, J. Global Semi-Arid Climate Change over Last 60 Years. Clim. Dyn. 2016, 46, 1131–1150. [Google Scholar] [CrossRef] [Green Version]
- Spinoni, J.; Barbosa, P.; Cherlet, M.; Forzieri, G.; McCormick, N.; Naumann, G.; Vogt, J.V.; Dosio, A. How Will the Progressive Global Increase of Arid Areas Affect Population and Land-Use in the 21st Century? Glob. Planet. Chang. 2021, 205, 103597. [Google Scholar] [CrossRef]
- Oliver, M.J.; Tuba, Z.; Mishler, B.D. The evolution of vegetative desiccation tolerance in land plants. Plant Ecol. 2000, 151, 85–100. [Google Scholar] [CrossRef]
- Schwab, K.B.; Schreiber, U.; Heber, U. Response of Photosynthesis and Respiration of Resurrection Plants to Desiccation and Rehydration. Planta 1989, 177, 217–227. [Google Scholar] [CrossRef]
- Georgieva, K.; Szigeti, Z.; Sarvari, E.; Gaspar, L.; Maslenkova, L.; Peeva, V.; Peli, E.; Tuba, Z. Photosynthetic Activity of Homoiochlorophyllous Desiccation Tolerant Plant Haberlea Rhodopensis during Dehydration and Rehydration. Planta 2007, 225, 955–964. [Google Scholar] [CrossRef]
- Degl’Innocenti, E.; Guidi, L.; Stevanovic, B.; Navari, F. CO2 Fixation and Chlorophyll a Fluorescence in Leaves of Ramonda Serbica during a Dehydration-Rehydration Cycle. J. Plant Physiol. 2008, 165, 723–733. [Google Scholar] [CrossRef]
- Dinakar, C.; Djilianov, D.; Bartels, D. Photosynthesis in Desiccation Tolerant Plants: Energy Metabolism and Antioxidative Stress Defense. Plant Sci. 2012, 182, 29–41. [Google Scholar] [CrossRef]
- Nadal, M.; Perera-Castro, A.V.; Gulías, J.; Farrant, J.M.; Flexas, J. Resurrection Plants Optimize Photosynthesis despite Very Thick Cell Walls by Means of Chloroplast Distribution. J. Exp. Bot. 2021, 72, 2600–2610. [Google Scholar] [CrossRef]
- Xu, X.; Legay, S.; Sergeant, K.; Zorzan, S.; Leclercq, C.C.; Charton, S.; Giarola, V.; Liu, X.; Challabathula, D.; Renaut, J.; et al. Molecular Insights into Plant Desiccation Tolerance: Transcriptomics, Proteomics and Targeted Metabolite Profiling in Craterostigma Plantagineum. Plant J. 2021, 107, 377–398. [Google Scholar] [CrossRef]
- Georgieva, K.; Sárvári, É.; Keresztes, Á. Protection of Thylakoids against Combined Light and Drought by a Lumenal Substance in the Resurrection Plant Haberlea Rhodopensis. Ann. Bot. 2010, 105, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, P.; Finazzi, G.; Bligny, R.; Moyankova, D.; Zasheva, D.; Boisson, A.-M.; Brugière, S.; Krasteva, V.; Alipieva, K.; Simova, S.; et al. In Vivo Spectroscopy and NMR Metabolite Fingerprinting Approaches to Connect the Dynamics of Photosynthetic and Metabolic Phenotypes in Resurrection Plant Haberlea Rhodopensis during Desiccation and Recovery. Front. Plant Sci. 2015, 6, 564. [Google Scholar] [CrossRef] [Green Version]
- Charuvi, D.; Nevo, R.; Shimoni, E.; Naveh, L.; Zia, A.; Adam, Z.; Farrant, J.M.; Kirchhoff, H.; Reich, Z. Photoprotection Conferred by Changes in Photosynthetic Protein Levels and Organization during Dehydration of a Homoiochlorophyllous Resurrection Plant. Plant Physiol. 2015, 167, 1554–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, T.; Sun, Y.; Luo, S.; Zhang, C.; Zhou, H.; Lin, H. Efficient Modulation of Photosynthetic Apparatus Confers Desiccation Tolerance in the Resurrection Plant Boea Hygrometrica. Plant Cell Physiol. 2017, 58, 1976–1990. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Moyankova, D.; Lin, C.-T.; Mladenov, P.; Sun, R.-Z.; Djilianov, D.; Deng, X. Transcriptome Reprogramming during Severe Dehydration Contributes to Physiological and Metabolic Changes in the Resurrection Plant Haberlea Rhodopensis. BMC Plant Biol. 2018, 18, 351. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, S.-J.; Zhang, S.-B.; Zhang, J.-L.; Cao, K.-F. Cyclic Electron Flow Plays an Important Role in Photoprotection for the Resurrection Plant Paraboea Rufescens under Drought Stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef]
- Zia, A.; Walker, B.J.; Oung, H.M.O.; Charuvi, D.; Jahns, P.; Cousins, A.B.; Farrant, J.M.; Reich, Z.; Kirchhoff, H. Protection of the Photosynthetic Apparatus against Dehydration Stress in the Resurrection Plant Craterostigma Pumilum. Plant J. 2016, 87, 664–680. [Google Scholar] [CrossRef] [Green Version]
- Apostolova, E.; Rashkova, M.; Anachkov, N.; Denev, I.; Toneva, V.; Minkov, I.; Yahubyan, G. Molecular Cloning and Characterization of CDNAs of the Superoxide Dismutase Gene Family in the Resurrection Plant Haberlea Rhodopensis. Plant. Physiol. Biochem. 2012, 55, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Vasileva, I.; Gigova, L.; Gesheva, E.; Simova-Stoilova, L.; Georgieva, K. Antioxidant Defense during Recovery of Resurrection Plant Haberlea Rhodopensis from Drought- and Freezing-Induced Desiccation. Plants 2022, 11, 175. [Google Scholar] [CrossRef]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.-R.; Marriott, A.S.; Bergström, E.; et al. Molecular Mechanisms of Desiccation Tolerance in the Resurrection Glacial Relic Haberlea Rhodopensis. Cell Mol. Life Sci. 2013, 70, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic Profiling of the Resurrection Plant Haberlea Rhodopensis during Desiccation and Recovery. Physiol. Plant. 2014, 152, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Sprenger, N.; Bortlik, K.; Boller, T.; Wiemken, A. Desiccation Increases Sucrose Levels in Ramonda and Haberlea, Two Genera of Resurrection Plants in the Gesneriaceae. Physiol. Plant. 1997, 100, 153–158. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Rienstra, J.; Dennis, T.J.; Farrant, J.M.; Ligterink, W.; Hilhorst, H. Structural Plasticity of Intrinsically Disordered LEA Proteins from Xerophyta Schlechteri Provides Protection In Vitro and In Vivo. Front. Plant Sci. 2019, 10, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Man Wai, C.; Pardo, J.; Giarola, V.; Ambrosini, S.; Song, X.; Bartels, D. Desiccation Tolerance Evolved through Gene Duplication and Network Rewiring in Lindernia. Plant Cell 2018, 30, 2943–2958. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.-C.D.; Cooper, K.; Hilhorst, H.W.M.; Farrant, J.M. Orthodox Seeds and Resurrection Plants: Two of a Kind? Plant Physiol. 2017, 175, 589–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinakar, C.; Bartels, D. Desiccation Tolerance in Resurrection Plants: New Insights from Transcriptome, Proteome and Metabolome Analysis. Front. Plant Sci. 2013, 4, 482. [Google Scholar] [CrossRef] [Green Version]
- Gechev, T.; Lyall, R.; Petrov, V.; Bartels, D. Systems Biology of Resurrection Plants. Cell Mol. Life Sci. 2021, 78, 6365–6394. [Google Scholar] [CrossRef] [PubMed]
- Koonjul, P.K.; Brandt, W.F.; Lindsey, G.G.; Farrant, J.M. Isolation and Characterisation of Chloroplasts from Myrothamnus Flabellifolius Welw. J. Plant Physiol. 2000, 156, 584–594. [Google Scholar] [CrossRef]
- Mladenov, P.; Zasheva, D.; Peshev, D.; Christov, N.K.; Rolland, N.; Djilianov, D.; Tchorbadjieva, M. Sub-cellular fractionation and gel-based proteomics of Haberlea rhodopensis: A promising approach to open the black box of resurrection plants. Bulg. J. Agric. Sci. 2013, 19, 22–25. [Google Scholar]
- Salvi, D.; Rolland, N.; Joyard, J.; Ferro, M. Purification and Proteomic Analysis of Chloroplasts and Their Sub-Organellar Compartments. In Organelle Proteomics; Pflieger, D., Rossier, J., Eds.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2008; pp. 19–36. ISBN 978-1-59745-028-7. [Google Scholar]
- Bouchnak, I.; Moyet, L.; Salvi, D.; Kuntz, M.; Rolland, N. Preparation of Chloroplast Sub-Compartments from Arabidopsis for the Analysis of Protein Localization by Immunoblotting or Proteomics. J. Vis. Exp. 2018, 140, e58581. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.G.E.; Mueller, S.J.; Hoernstein, S.N.W.; Porankiewicz-Asplund, J.; Vervliet-Scheebaum, M.; Reski, R. Simultaneous Isolation of Pure and Intact Chloroplasts and Mitochondria from Moss as the Basis for Sub-Cellular Proteomics. Plant Cell Rep. 2011, 30, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, H. Differences in Rehydration of Three Desiccation-Tolerant Angiosperm Species. Ann. Bot. 1996, 78, 703–710. [Google Scholar] [CrossRef]
- Vecchia, F.D.; El Asmar, T.; Calamassi, R.; Rascio, N.; Vazzana, C. Morphological and Ultrastructural Aspects of Dehydration and Rehydration in Leaves of Sporobolus Stapfianus. Plant Growth Regul. 1998, 24, 219–228. [Google Scholar] [CrossRef]
- Georgieva, K.; Solti, Á.; Mészáros, I.; Keresztes, Á.; Sárvári, É. Light Sensitivity of Haberlea Rhodopensis Shade Adapted Phenotype under Drought Stress. Acta Physiol. Plant. 2017, 39, 164. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Koita, H.; Nakatsubo, T.; Hasegawa, K.; Wakabayashi, K.; Takahashi, H.; Umemura, K.; Umezawa, T.; Shimamoto, K. Cinnamoyl-CoA Reductase, a Key Enzyme in Lignin Biosynthesis, Is an Effector of Small GTPase Rac in Defense Signaling in Rice. Proc. Natl. Acad. Sci. USA 2006, 103, 230–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Jung, N.U.; Giarola, V.; Bartels, D. The Dynamic Responses of Cell Walls in Resurrection Plants During Dehydration and Rehydration. Front. Plant Sci. 2020, 10, 1698. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.-W.; Verma, R.; Kim, M.; Lee, J.-Y.; Kim, Y.-K.; Bang, J.-W.; Reiter, W.-D.; Pai, H.-S. Depletion of UDP-D-Apiose/UDP-D-Xylose Synthases Results in Rhamnogalacturonan-II Deficiency, Cell Wall Thickening, and Cell Death in Higher Plants. J. Biol. Chem. 2006, 281, 13708–13716. [Google Scholar] [CrossRef] [Green Version]
- Vicré, M.; Sherwin, H.W.; Driouich, A.; Jaffer, M.A.; Farrant, J.M. Cell Wall Characteristics and Structure of Hydrated and Dry Leaves of the Resurrection Plant Craterostigma Wilmsii, a Microscopical Study. J. Plant Physiol. 1999, 155, 719–726. [Google Scholar] [CrossRef]
- Jung, N.U.; Giarola, V.; Chen, P.; Knox, J.P.; Bartels, D. Craterostigma Plantagineum Cell Wall Composition Is Remodelled during Desiccation and the Glycine-Rich Protein CpGRP1 Interacts with Pectins through Clustered Arginines. Plant J. 2019, 100, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Kohorn, B.D.; Yakir, D. Movement of Newly Imported Light-Harvesting Chlorophyll-Binding Protein from Unstacked to Stacked Thylakoid Membranes Is Not Affected by Light Treatment or Absence of Amino-Terminal Threonines. J. Biol. Chem. 1990, 265, 2118–2123. [Google Scholar] [CrossRef]
- Minagawa, J. State Transitions—The Molecular Remodeling of Photosynthetic Supercomplexes That Controls Energy Flow in the Chloroplast. Biochim. Biophys. Acta 2011, 1807, 897–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesaresi, P.; Pribil, M.; Wunder, T.; Leister, D. Dynamics of Reversible Protein Phosphorylation in Thylakoids of Flowering Plants: The Roles of STN7, STN8 and TAP38. Biochim. Biophys. Acta 2011, 1807, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasi, S.; Venkatesh, J.; Daneshi, R.F.; Gururani, M.A. Photosystem II Extrinsic Proteins and Their Putative Role in Abiotic Stress Tolerance in Higher Plants. Plants 2018, 7, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, R.; Ifuku, K.; Takabayashi, A.; Shikanai, T.; Endo, T.; Sato, F. Characterization of an Arabidopsis Thaliana Mutant with Impaired PsbO, One of Two Genes Encoding Extrinsic 33-KDa Proteins in Photosystem II. FEBS Lett. 2002, 523, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.J.; Oh, Y.A.; Lee, E.S.; Park, A.R.; Cho, S.K.; Yoo, Y.J.; Park, O.K. Oxygen-Evolving Enhancer Protein 2 Is Phosphorylated by Glycine-Rich Protein 3/Wall-Associated Kinase 1 in Arabidopsis. Biochem. Biophys. Res. Commun. 2003, 305, 862–868. [Google Scholar] [CrossRef]
- Chen, P.; Giarola, V.; Bartels, D. The Craterostigma Plantagineum Protein Kinase CpWAK1 Interacts with Pectin and Integrates Different Environmental Signals in the Cell Wall. Planta 2021, 253, 92. [Google Scholar] [CrossRef]
- Xiao, L.; Yang, G.; Zhang, L.; Yang, X.; Zhao, S.; Ji, Z.; Zhou, Q.; Hu, M.; Wang, Y.; Chen, M.; et al. The Resurrection Genome of Boea Hygrometrica: A Blueprint for Survival of Dehydration. Proc. Natl. Acad. Sci. USA 2015, 112, 5833–5837. [Google Scholar] [CrossRef] [Green Version]
- Dumas, L.; Chazaux, M.; Peltier, G.; Johnson, X.; Alric, J. Cytochrome B6f Function and Localization, Phosphorylation State of Thylakoid Membrane Proteins and Consequences on Cyclic Electron Flow. Photosynth. Res. 2016, 129, 307–320. [Google Scholar] [CrossRef]
- Jiang, G.; Wang, Z.; Shang, H.; Yang, W.; Hu, Z.; Phillips, J.; Deng, X. Proteome Analysis of Leaves from the Resurrection Plant Boea Hygrometrica in Response to Dehydration and Rehydration. Planta 2006, 225, 1405. [Google Scholar] [CrossRef]
- Sullivan, M.L. Beyond Brown: Polyphenol Oxidases as Enzymes of Plant Specialized Metabolism. Front. Plant Sci. 2015, 5, 783. [Google Scholar] [CrossRef] [Green Version]
- Young, M.J.; Bay, D.C.; Hausner, G.; Court, D.A. The Evolutionary History of Mitochondrial Porins. BMC Evol. Biol. 2007, 7, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Baker, A.; Bartel, B.; Linka, N.; Mullen, R.T.; Reumann, S.; Zolman, B.K. Plant Peroxisomes: Biogenesis and Function. Plant Cell 2012, 24, 2279–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charuvi, D.; Nevo, R.; Aviv-Sharon, E.; Gal, A.; Kiss, V.; Shimoni, E.; Farrant, J.M.; Kirchhoff, H.; Reich, Z. Chloroplast Breakdown during Dehydration of a Homoiochlorophyllous Resurrection Plant Proceeds via Senescence-like Processes. Environ. Exp. Bot. 2019, 157, 100–111. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, B.; Phillips, J.; Zhang, Z.-N.; Du, H.; Xu, T.; Huang, L.-C.; Zhang, X.-F.; Xu, G.-H.; Li, W.-L.; et al. Global Transcriptome Analysis Reveals Acclimation-Primed Processes Involved in the Acquisition of Desiccation Tolerance in Boea Hygrometrica. Plant Cell Physiol. 2015, 56, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Du, H.; Zhang, Z.; Xu, W.; Deng, X. BhbZIP60 from Resurrection Plant Boea Hygrometrica Is an MRNA Splicing-Activated Endoplasmic Reticulum Stress Regulator Involved in Drought Tolerance. Front. Plant Sci. 2017, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avin-Wittenberg, T.; Baluška, F.; Bozhkov, P.V.; Elander, P.H.; Fernie, A.R.; Galili, G.; Hassan, A.; Hofius, D.; Isono, E.; Le Bars, R.; et al. Autophagy-Related Approaches for Improving Nutrient Use Efficiency and Crop Yield Protection. J. Exp. Bot. 2018, 69, 1335–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, M.; Nakamura, S.; Li, N. Autophagic Turnover of Chloroplasts: Its Roles and Regulatory Mechanisms in Response to Sugar Starvation. Front. Plant Sci. 2019, 10, 280. [Google Scholar] [CrossRef]
- Zhu, P.; Yu, X.-H.; Wang, C.; Zhang, Q.; Liu, W.; McSweeney, S.; Shanklin, J.; Lam, E.; Liu, Q. Structural Basis for Ca2+-Dependent Activation of a Plant Metacaspase. Nat. Commun. 2020, 11, 2249. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.; Njaci, I.; Moghaddam, L.; Long, H.; Dickman, M.B.; Zhang, X.; Mundree, S. Trehalose Accumulation Triggers Autophagy during Plant Desiccation. PLoS Genet. 2015, 11, e1005705. [Google Scholar] [CrossRef] [Green Version]
- Guidi, L.; Tattini, M. Antioxidant Defenses in Plants: A Dated Topic of Current Interest. Antioxidants 2021, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Nazarenus, T.J.; Frey, J.L.; Liang, X.; Wilson, M.A.; Stone, J.M. A Plant DJ-1 Homolog Is Essential for Arabidopsis Thaliana Chloroplast Development. PLoS ONE 2011, 6, e23731. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Lin, H.; Maple, J.; Björkblom, B.; Alves, G.; Larsen, J.P.; Møller, S.G. The Arabidopsis DJ-1a Protein Confers Stress Protection through Cytosolic SOD Activation. J. Cell Sci. 2010, 123, 1644–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrant, J.M.; Cooper, K.; Hilgart, A.; Abdalla, K.O.; Bentley, J.; Thomson, J.A.; Dace, H.J.W.; Peton, N.; Mundree, S.G.; Rafudeen, M.S. A Molecular Physiological Review of Vegetative Desiccation Tolerance in the Resurrection Plant Xerophyta Viscosa (Baker). Planta 2015, 242, 407–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jesús-Pires, C.; Ferreira-Neto, J.R.C.; Pacifico Bezerra-Neto, J.; Kido, E.A.; de Oliveira Silva, R.L.; Pandolfi, V.; Wanderley-Nogueira, A.C.; Binneck, E.; da Costa, A.F.; Pio-Ribeiro, G.; et al. Plant Thaumatin-like Proteins: Function, Evolution and Biotechnological Applications. Curr. Protein Pept. Sci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Zhang, L. Structural and Functional Dynamics of Dehydrins: A Plant Protector Protein under Abiotic Stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, Q.; Li, D.; Yang, X.; Li, D. Multifunctional Roles of Plant Dehydrins in Response to Environmental Stresses. Front. Plant Sci. 2017, 8, 1018. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Tang, M.-J.; Hu, Y.-L.; Lin, Z.-P. Isolation and Characterization of a Dehydrin-like Gene from Drought-Tolerant Boea Crassifolia. Plant Sci. 2004, 166, 1167–1175. [Google Scholar] [CrossRef]
- Maszkowska, J.; Dębski, J.; Kulik, A.; Kistowski, M.; Bucholc, M.; Lichocka, M.; Klimecka, M.; Sztatelman, O.; Szymańska, K.P.; Dadlez, M.; et al. Phosphoproteomic Analysis Reveals That Dehydrins ERD10 and ERD14 Are Phosphorylated by SNF1-Related Protein Kinase 2.10 in Response to Osmotic Stress. Plant Cell Environ. 2019, 42, 931–946. [Google Scholar] [CrossRef]
- Flori, S.; Jouneau, P.-H.; Gallet, B.; Estrozi, L.F.; Moriscot, C.; Schoehn, G.; Finazzi, G.; Falconet, D. Imaging Plastids in 2D and 3D: Confocal and Electron Microscopy. Methods Mol. Biol. 2018, 1829, 113–122. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 Update of the PRIDE Database and Its Related Tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef]
- Seigneurin-Berny, D.; Salvi, D.; Joyard, J.; Rolland, N. Purification of Intact Chloroplasts from Arabidopsis and Spinach Leaves by Isopycnic Centrifugation. Curr. Protoc. Cell Biol. 2008, 40, 3.30.1–3.30.14. [Google Scholar] [CrossRef]
- Vallon, O.; Wollman, F.A.; Olive, J. Lateral Distribution of the Main Protein Complexes of the Photosynthetic Apparatus in Chlamydomonas Reinhardtii and in Spinach: An Immunocytochemical Study Using Intact Thylakoid Membranes and a PS II Enriched Membrane Preparation. Photobiochem. Photobiophys. 1986, 12, 203–220. [Google Scholar]
- Perryman, R.A.; Mooney, B.; Harmey, M.A. Identification of a 42-KDa Plant Mitochondrial Outer Membrane Protein, MOM42, Involved in the Import of Precursor Proteins into Plant Mitochondria. Arch. Biochem. Biophys. 1995, 316, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vignani, R.; Scali, M.; Cresti, M. A Universal and Rapid Protocol for Protein Extraction from Recalcitrant Plant Tissues for Proteomic Analysis. Electrophoresis 2006, 27, 2782–2786. [Google Scholar] [CrossRef] [PubMed]
- Ferro, M.; Brugière, S.; Salvi, D.; Seigneurin-Berny, D.; Court, M.; Moyet, L.; Ramus, C.; Miras, S.; Mellal, M.; Le Gall, S.; et al. AT_CHLORO, a Comprehensive Chloroplast Proteome Database with Subplastidial Localization and Curated Information on Envelope Proteins. Mol. Cell Proteom. 2010, 9, 1063–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape Plug-in to Decipher Functionally Grouped Gene Ontology and Pathway Annotation Networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Nersisyan, L.; Johnson, G.; Riel-Mehan, M.; Pico, A.R.; Arakelyan, A. PSFC: A Pathway Signal Flow Calculator App for Cytoscape. F1000Res 2017, 4, 480. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gel Annotation | Protein Score | Name | Gene | Crude | Phospho | Low Abundant | Stroma | Thylakoids | Mito |
|---|---|---|---|---|---|---|---|---|---|
| 2 | 177 | Chlorophyll a-b binding protein 40 | CAB40 |  |  |  |  |  |  |
| 3 | 122 | Stem-specific protein | TSJT1 |  |  |  |  |  |  |
| 11 | 179 | ribulose bisphosphate carboxylase/oxygenase activase | RCA |  | N/D |  |  |  |  |
| 18 | 222 | Ribulose bisphosphate carboxylase large chain | RuBisCO |  | N/D |  |  |  |  |
| 12 | 95 | Oxygen-evolving enhancer protein 1 | OEE1 |  |  |  |  |  |  |
| 13 | 136 | Kunitz-type trypsin inhibitor B3 | BPTI-3 | N/D | N/D |  |  |  |  |
| 14 | 240 | quinone oxidoreductase-like protein | At1g23740 |  | N/D |  |  |  |  |
| 15 | 177 | phosphoglycerate kinase | PGKA | N/D | N/D |  |  |  |  |
| 16 | 150 | proteasome subunit alpha type-2-A-like | L195_g004424 | N/D | N/D |  |  |  |  |
| 17 | 331 | N/A | N/A | N/D | N/D |  |  |  |  |
| 20 | 131 | dehydrin-like protein | Dh2 |  |  |  |  |  |  |
| 21 | 219 | hypothetical protein | N/A |  |  | N/D |  |  |  |
| 22 | 86 | hypothetical protein | N/A |  |  |  |  |  |  |
| 23 | 86 | hypothetical protein | N/A | N/D |  |  |  |  |  |
| 24 | 273 | hypothetical protein | N/A | N/D |  | N/D |  |  |  |
| 26 | 211 | polyphenol oxidase | PPO |  | N/D | N/D |  |  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mladenov, P.; Zasheva, D.; Planchon, S.; Leclercq, C.C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis. Int. J. Mol. Sci. 2022, 23, 8520. https://doi.org/10.3390/ijms23158520
Mladenov P, Zasheva D, Planchon S, Leclercq CC, Falconet D, Moyet L, Brugière S, Moyankova D, Tchorbadjieva M, Ferro M, et al. Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis. International Journal of Molecular Sciences. 2022; 23(15):8520. https://doi.org/10.3390/ijms23158520
Chicago/Turabian StyleMladenov, Petko, Diana Zasheva, Sébastien Planchon, Céline C. Leclercq, Denis Falconet, Lucas Moyet, Sabine Brugière, Daniela Moyankova, Magdalena Tchorbadjieva, Myriam Ferro, and et al. 2022. "Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis" International Journal of Molecular Sciences 23, no. 15: 8520. https://doi.org/10.3390/ijms23158520