
The Promising Nanovectors for Gene Delivery in Plant Genome Engineering

 , and
, and
Abstract
:1. Introduction
2. The Merits and Limitations of Traditional Delivery Methods

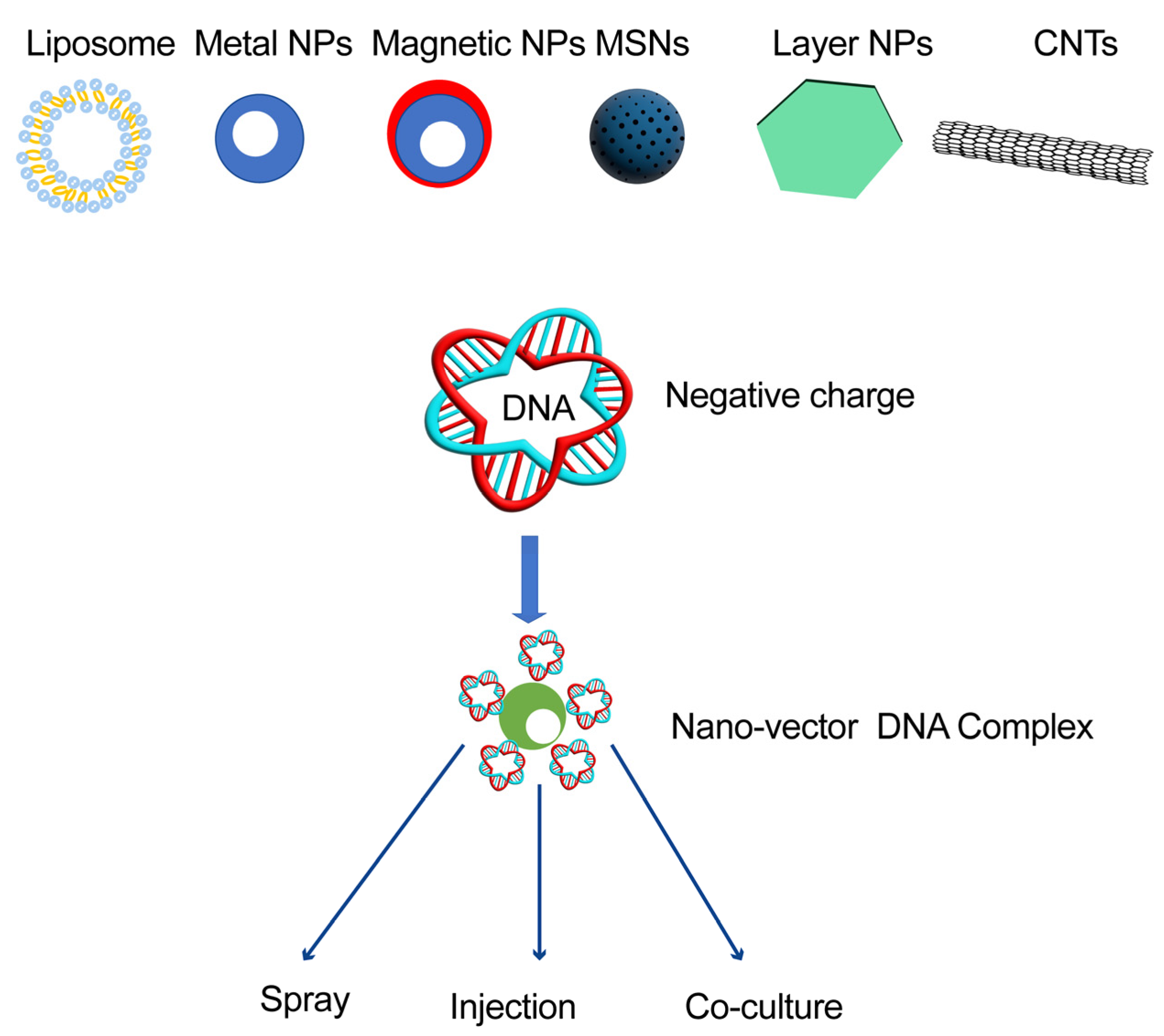{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traditional Delivery Methods | Merits | Limitations | Cargo Types | Ref. |
|---|---|---|---|---|
| Agrobacterium | Well-established protocols, low cost and widely used | Genotype-dependent; limited cargo type; Antagonism between Agrobacterium and plants, not appliable in several plant species; cargo type limitation | Plasmid DNA | [22,26] |
| Particle bombardment | Promising in the genome engineering of mitochondria and chloroplasts, suitable for all cargos | Random insertions, tissue type depended, host genome damages often happen, expensive equipment and materials | Plasmid DNA, RNA, RNPs, synthesized oligonucleotide | [37,38,41] |
| PEG | Highly efficient in protoplast, suitable for all cargos | Time-consuming, cell limitations, regeneration inefficient, polyploid formation | Plasmid DNA, RNA, RNPs, synthesized oligonucleotide | [13,42] |
| Plant virus | Genotype-independent, high level of transient expression | Cargo size limitations, plant species restricted, safety concern in crop yield | DNA, RNA | [46,48] |
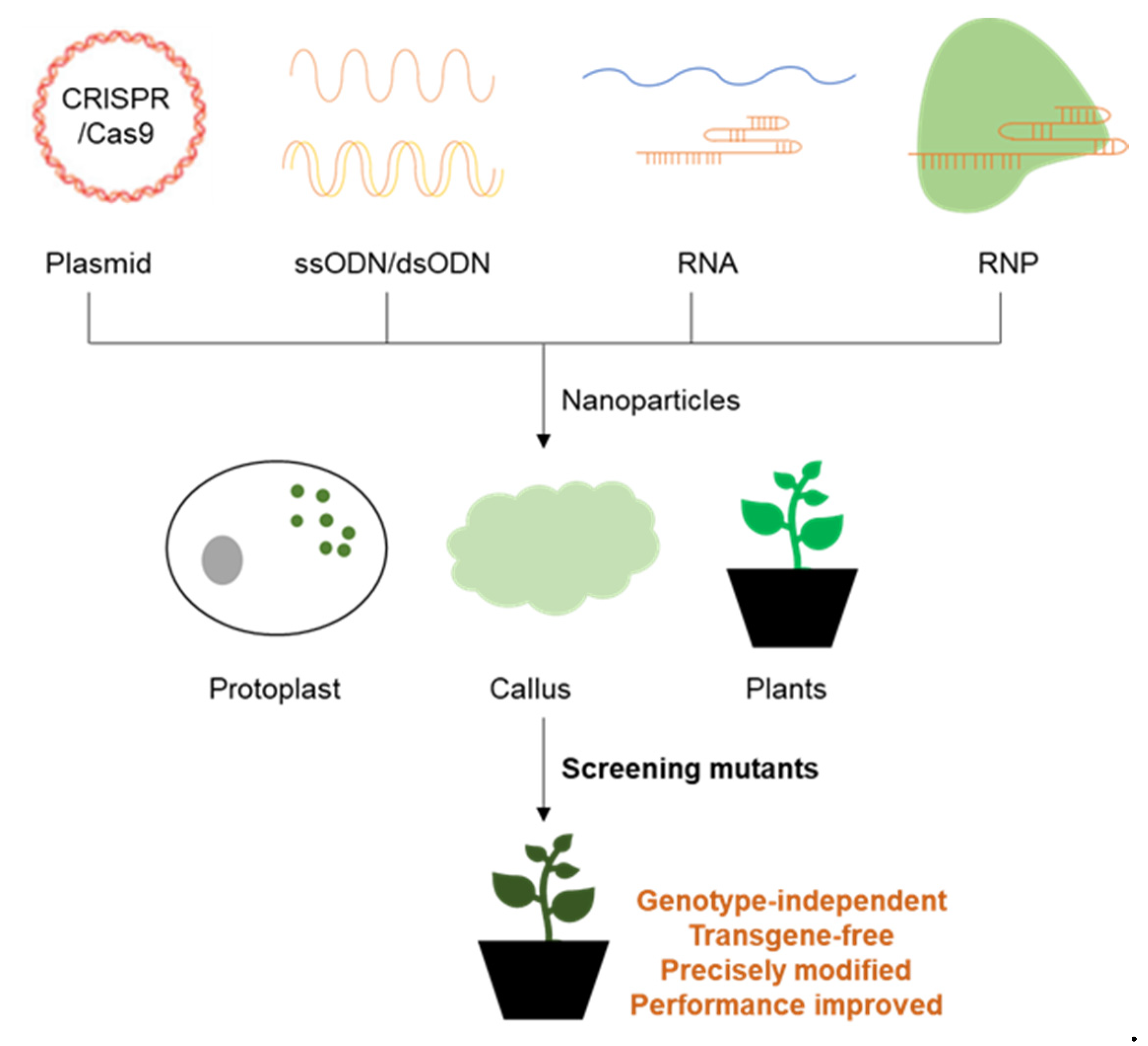
3. Nanomaterial-Induced Gene Delivery Systems
4. Future Prospects for Plant Genetic Engineering with Nanomaterials
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molla, K.A.; Sretenovic, S.; Bansal, K.C.; Qi, Y.J.N.P. Precise plant genome editing using base editors and prime editors. Nat. Plants 2021, 7, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, P.D.; Dively, G.P. Climate change, transgenic corn adoption and field-evolved resistance in corn earworm. R. Soc. Open Sci. 2017, 4, 170210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Altpeter, F.; Baisakh, N.; Beachy, R.; Bock, R.; Capell, T.; Christou, P.; Daniell, H.; Datta, K.; Datta, S.; Dix, P.J.; et al. Particle bombardment and the genetic enhancement of crops: Myths and realities. Mol. Breed. 2005, 15, 305–327. [Google Scholar] [CrossRef]
- Delporte, F.; Pretova, A.; du Jardin, P.; Watillon, B. Morpho-histology and genotype dependence of in vitro morphogenesis in mature embryo cultures of wheat. Protoplasma 2014, 251, 1455–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aesaert, S.; Impens, L.; Coussens, G.; Van Lerberge, E.; Vanderhaeghen, R.; Desmet, L.; Vanhevel, Y.; Bossuyt, S.; Wambua, A.N.; Van Lijsebettens, M.; et al. Optimized Transformation and Gene Editing of the B104 Public Maize Inbred by Improved Tissue Culture and Use of Morphogenic Regulators. Front. Plant Sci. 2022, 13, 883847. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Tricoli, D.M.; Ercoli, M.F.; Hayta, S.; Ronald, P.; Palatnik, J.F.; Dubcovsky, J. A GRF-GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 2020, 38, 1274–1279. [Google Scholar] [CrossRef]
- Kong, J.; Martin-Ortigosa, S.; Finer, J.; Orchard, N.; Gunadi, A.; Batts, L.A.; Thakare, D.; Rush, B.; Schmitz, O.; Stuiver, M.; et al. Overexpression of the Transcription Factor GROWTH-REGULATING FACTOR5 Improves Transformation of Dicot and Monocot Species. Front. Plant Sci. 2020, 11, 572319. [Google Scholar] [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Xing, S.; Xue, C.; Liu, J.; Chen, K.; Chai, T.; Gao, C. Transient expression of a TaGRF4-TaGIF1 complex stimulates wheat regeneration and improves genome editing. Sci. China Life Sci. 2022, 65, 731–738. [Google Scholar] [CrossRef]
- Pan, W.; Cheng, Z.; Han, Z.; Yang, H.; Zhang, W.; Zhang, H. Efficient genetic transformation and CRISPR/Cas9-mediated genome editing of watermelon assisted by genes encoding developmental regulators. J. Zhejiang Univ. Sci. B 2022, 23, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Sharma, A.; Toepfer, C.N.; Ward, T.; Wasson, L.; Agarwal, R.; Conner, D.A.; Hu, J.H.; Seidman, C.E. CRISPR/Cas9-Mediated Fluorescent Tagging of Endogenous Proteins in Human Pluripotent Stem Cells. Curr. Protoc. Hum. Genet. 2018, 96, 21.11.1–21.11.20. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tian, Y.; Shen, R.; Yao, Q.; Wang, M.; Chen, M.; Dong, J.; Zhang, T.; Li, F.; Lei, M.; et al. Targeted, efficient sequence insertion and replacement in rice. Nat. Biotechnol. 2020, 38, 1402–1407. [Google Scholar] [CrossRef]
- Duensing, N.; Sprink, T.; Parrott, W.A.; Fedorova, M.; Lema, M.A.; Wolt, J.D.; Bartsch, D. Novel Features and Considerations for ERA and Regulation of Crops Produced by Genome Editing. Front. Bioeng. Biotechnol. 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feynman, R.P. There’s Plenty of Room at the Bottom. J. Microelectromechanical Syst. 2011, 16, 890. [Google Scholar] [CrossRef] [Green Version]
- Thiruvengadam, M.; Rajakumar, G.; Chung, I.-M. Nanotechnology: Current uses and future applications in the food industry. 3 Biotech 2018, 8, 74. [Google Scholar] [CrossRef]
- Steinwand, M.A.; Ronald, P.C. Crop biotechnology and the future of food. Nat. Food 2020, 1, 273–283. [Google Scholar] [CrossRef]
- Jähne, A.; Becker, D.; Euphytica, H.L.J. Genetic engineering of cereal crop plants: A review. Euphytica 1995, 85, 35–44. [Google Scholar] [CrossRef]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular uptake of nanoparticles: Journey inside the cell. Chem. Soc. Rev. 2017, 46, 4218–4244. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, N.A.; Prakash, C.S.; McHughen, A.G. Genome editing for crop improvement: Challenges and opportunities. GM Crops Food 2015, 6, 183–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, J.E.; Christie, P.J. The Agrobacterium Ti Plasmids. Microbiol. Spectr. 2014, 2, 455–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelvin, S.B. Plant DNA Repair and Agrobacterium T-DNA Integration. Int. J. Mol. Sci. 2021, 22, 8458. [Google Scholar] [CrossRef]
- Wu, C.F.; Smith, D.A.; Lai, E.M.; Chang, J.H. The Agrobacterium Type VI Secretion System: A Contractile Nanomachine for Interbacterial Competition. Curr. Top. Microbiol. Immunol. 2018, 418, 215–231. [Google Scholar]
- Gelvin, S.B. Integration of Agrobacterium T-DNA into the Plant Genome. Annu. Rev. Genet. 2017, 51, 195–217. [Google Scholar] [CrossRef]
- Pu, X.-A.; Goodman, R.N. Induction of necrogenesis by Agrobacterium tumefaciens on grape explants. Physiol. Mol. Plant Pathol. 1992, 41, 241–254. [Google Scholar] [CrossRef]
- Deng, W.; Pu, X.A.; Goodman, R.N.; Gordon, M.P.; Nester, E.W. T-DNA genes responsible for inducing a necrotic response on grape vines. Mol. Plant-Microbe Interact. 1995, 8, 538–548. [Google Scholar] [CrossRef]
- Das, D.; Reddy, M.; Upadhyaya, K.; Sopory, S. An efficient leaf-disc culture method for the regeneration via somatic embryogenesis and transformation of grape (Vitis vinifera L.). Plant Cell Rep. 2002, 20, 999–1005. [Google Scholar] [CrossRef]
- Ciardi, J.A.; Tieman, D.M.; Lund, S.T.; Jeffrey, B.J.; Stall, R.E.; Klee, H.J. Response to Xanthomonas campestris pv. vesicatoria in Tomato Involves Regulation of Ethylene Receptor Gene Expression. Plant Physiol. 2000, 123, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, S.; Ezura, H. Plant–Agrobacterium interaction mediated by ethylene and super-Agrobacterium conferring efficient gene transfer. Front. Plant Sci. 2014, 5, 681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, S.; Someya, T.; Zhou, S.; Takayama, M.; Nakamura, K.; Ezura, H. An Agrobacterium tumefaciens Strain with Gamma-Aminobutyric Acid Transaminase Activity Shows an Enhanced Genetic Transformation Ability in Plants. Sci. Rep. 2017, 7, 42649. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.J.; Mock, N.M.; Whitaker, B.D.; Roberts, D.P.; Rice, C.P.; Deahl, K.L.; Aver’yanov, A.A. Involvement of acetosyringone in plant–pathogen recognition. Biochem. Biophys. Res. Commun. 2005, 328, 130–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Liu, L.; Zhang, H. Transgene-free Genome Editing in Plants. Front. Genome Ed. 2021, 3, 805317. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Choi, S.; Park, S.; Yoon, J.; Park, A.Y.; Choe, S. DNA-Free Genome Editing via Ribonucleoprotein (RNP) Delivery of CRISPR/Cas in Lettuce. Methods Mol. Biol. 2019, 1917, 337–354. [Google Scholar]
- Jinturkar, K.A.; Rathi, M.N.; Misra, A. 3—Gene Delivery Using Physical Methods. In Challenges in Delivery of Therapeutic Genomics and Proteomics; Misra, A., Ed.; Elsevier: London, UK, 2011; pp. 83–126. [Google Scholar]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [Green Version]
- Baltes, N.J.; Gil-Humanes, J.; Voytas, D.F. Chapter One—Genome Engineering and Agriculture: Opportunities and Challenges. In Progress in Molecular Biology and Translational Science; Weeks, D.P., Yang, B., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 149, pp. 1–26. [Google Scholar]
- Banakar, R.; Eggenberger, A.L.; Lee, K.; Wright, D.A.; Murugan, K.; Zarecor, S.; Lawrence-Dill, C.J.; Sashital, D.G.; Wang, K. High-frequency random DNA insertions upon co-delivery of CRISPR-Cas9 ribonucleoprotein and selectable marker plasmid in rice. Sci. Rep. 2019, 9, 19902. [Google Scholar] [CrossRef]
- Liu, J.; Nannas, N.J.; Fu, F.F.; Shi, J.; Aspinwall, B.; Parrott, W.A.; Dawe, R.K. Genome-Scale Sequence Disruption Following Biolistic Transformation in Rice and Maize. Plant Cell 2019, 31, 368–383. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.-D.; Cho, Y.-H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Hsu, C.T.; Yang, L.H.; Lee, L.Y.; Fu, J.Y.; Cheng, Q.W.; Wu, F.H.; Hsiao, H.C.; Zhang, Y.; Zhang, R.; et al. Application of protoplast technology to CRISPR/Cas9 mutagenesis: From single-cell mutation detection to mutant plant regeneration. Plant Biotechnol. J. 2018, 16, 1295–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maćkowska, K.; Jarosz, A.; Grzebelus, E. Plant regeneration from leaf-derived protoplasts within the Daucus genus: Effect of different conditions in alginate embedding and phytosulfokine application. Lant Cell Tiss Organ Cult. 2014, 117, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like immune system conferring DNA virus resistance in plants. Nat. Plants 2015, 1, 15144. [Google Scholar] [CrossRef] [PubMed]
- Dinesh-Kumar, S.P.; Anandalakshmi, R.; Marathe, R.; Schiff, M.; Liu, Y. Virus-induced gene silencing. Methods Mol. Biol. 2003, 236, 287–294. [Google Scholar]
- Torti, S.; Schlesier, R.; Thümmler, A.; Bartels, D.; Römer, P.; Koch, B.; Werner, S.; Panwar, V.; Kanyuka, K.; Wirén, N.v.; et al. Transient reprogramming of crop plants for agronomic performance. Nat. Plants 2021, 7, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhang, X.; Liu, H.; Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR–Cas9. Nat. Plants 2020, 6, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Hao, M.; Tian, B.; Cao, G.; Wei, F.; Xie, Z. A Methodological Advance of Tobacco Rattle Virus-Induced Gene Silencing for Functional Genomics in Plants. Front Plant Sci. 2021, 12, 671091. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Technical Advance. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef]
- Buriak, J.M.; Liz-Marzán, L.M.; Parak, W.J.; Chen, X. Nano and Plants. ACS Nano 2022, 16, 1681–1684. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol 2018, 13, 677–684. [Google Scholar] [CrossRef]
- Hamers, R.J. Nanomaterials and Global Sustainability. Acc. Chem. Res. 2017, 50, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Misra, R.P.; Giraldo, J.P.; Kwak, S.-Y.; Son, Y.; Landry, M.P.; Swan, J.W.; Blankschtein, D.; Strano, M.S. Lipid Exchange Envelope Penetration (LEEP) of Nanoparticles for Plant Engineering: A Universal Localization Mechanism. Nano Lett. 2016, 16, 1161–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, K.; Pramanik, P.; Maity, A.; Joshi, D.C.; Wani, S.H.; Krishnan, P. Chapter 4—Methods of Using Nanomaterials to Plant Systems and Their Delivery to Plants (Mode of Entry, Uptake, Translocation, Accumulation, Biotransformation and Barriers). In Advances in Phytonanotechnology; Ghorbanpour, M., Wani, S.H., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 123–152. [Google Scholar]
- Zhi, H.; Chen, H.; Yu, M.; Wang, C.; Cui, B.; Zhao, X.; Wang, Y.; Cui, H.; Zhang, B.; Zeng, Z. Layered Double Hydroxide Nanosheets Improve the Adhesion of Fungicides to Leaves and the Antifungal Performance. ACS Appl. Nano Mater. 2022, 5, 5316–5325. [Google Scholar] [CrossRef]
- Yong, J.; Zhang, R.; Bi, S.; Li, P.; Sun, L.; Neena, M.; Carroll, B.J.; Xu, Z.P. Sheet-like clay nanoparticles deliver RNA into developing pollen to efficiently silence a target gene. Plant Physiol. 2021, 2, 886–899. [Google Scholar] [CrossRef]
- Demirer, G.S.; Zhang, H.; Matos, J.L.; Goh, N.S.; Cunningham, F.J.; Sung, Y.; Chang, R.; Aditham, A.J.; Chio, L.; Cho, M.-J.; et al. High aspect ratio nanomaterials enable delivery of functional genetic material without DNA integration in mature plants. Nat. Nanotechnol. 2019, 14, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Gumustas, M.; Sengel-Turk, C.T.; Gumustas, A.; Ozkan, S.A.; Uslu, B. Chapter 5—Effect of Polymer-Based Nanoparticles on the Assay of Antimicrobial Drug Delivery Systems. In Multifunctional Systems for Combined Delivery, Biosensing and Diagnostics; Grumezescu, A.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 67–108. [Google Scholar]
- Zhang, H.; Goh, N.S.; Wang, J.W.; Pinals, R.L.; González-Grandío, E.; Demirer, G.S.; Butrus, S.; Fakra, S.C.; Del Rio Flores, A.; Zhai, R.; et al. Nanoparticle cellular internalization is not required for RNA delivery to mature plant leaves. Nat. Nanotechnol. 2022, 17, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Sharma, A.R.; Bhattacharya, M.; Lee, S.S.; Chakraborty, C. CRISPR-Cas9: A Preclinical and Clinical Perspective for the Treatment of Human Diseases. Mol. Ther. 2021, 29, 571–586. [Google Scholar] [CrossRef]
- Jonsson, K.; Hamant, O.; Bhalerao, R.P. Plant cell walls as mechanical signaling hubs for morphogenesis. Curr. Biol. 2022, 32, 334–340. [Google Scholar] [CrossRef]
- Kaeppler, H.F.; Somers, D.A.; Rines, H.W.; Cockburn, A.F. Silicon carbide fiber-mediated stable transformation of plant cells. Theor. Appl. Genet. 1992, 84, 560–566. [Google Scholar] [CrossRef]
- Serik, O.; Ainur, I.; Murat, K.; Tetsno, M.; Masaki, I. Silicon carbide fiber-mediated DNA delivery into cells of wheat (Triticum acstivum L.) mature embryos. Plant Cell Rep. 1996, 16, 133–136. [Google Scholar] [CrossRef]
- Asad, S.; Mukhtar, Z.; Nazir, F.; Hashmi, J.A.; Mansoor, S.; Zafar, Y.; Arshad, M. Silicon carbide whisker-mediated embryogenic callus transformation of cotton (Gossypium hirsutum L.) and regeneration of salt tolerant plants. Mol. Biotechnol. 2008, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Abdul Razak, N.A.; Othman, N.H.; Mat Shayuti, M.S.; Jumahat, A.; Sapiai, N.; Lau, W.J. Agricultural and industrial waste-derived mesoporous silica nanoparticles: A review on chemical synthesis route. J. Environ. Chem. Eng. 2022, 10, 107322. [Google Scholar] [CrossRef]
- Kaziem, A.E.; Yang, L.; Lin, Y.; Song, Z.; Xu, H.; Zhang, Z. Efficiency of mesoporous silica/carboxymethyl β-glucan as a fungicide nano-delivery system for improving chlorothalonil bioactivity and reduce biotoxicity. Chemosphere 2022, 287, 131902. [Google Scholar] [CrossRef]
- Torney, F.; Trewyn, B.G.; Lin, V.S.Y.; Wang, K. Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nat. Nanotechnol 2007, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ortigosa, S.; Valenstein, J.S.; Lin, S.Y.; Trewyn, B.C.; Wang, K. Gold Functionalized Mesoporous Silica Nanoparticle Mediated Protein and DNA Codelivery to Plant Cells Via the Biolistic Method. Adv. Funct. Materials 2012, 22, 3576–3582. [Google Scholar] [CrossRef]
- Hajiahmadi, Z.; Shirzadian-Khorramabad, R.; Kazemzad, M.; Sohani, M.M. Enhancement of tomato resistance to Tuta absoluta using a new efficient mesoporous silica nanoparticle-mediated plant transient gene expression approach. Sci. Hortic. 2019, 243, 367–375. [Google Scholar] [CrossRef]
- Yuzhi, H.; Xiaoyan, Y.; Yongzhong, S.; Steven, S.; James, X.; Janet, M.; Jianmin, C.; Jie, C. Magnetic gold nanoparticles as a vehicle for fluorescein isothiocyanate and DNA delivery into plant cells. Botany 2013, 91, 457–466. [Google Scholar]
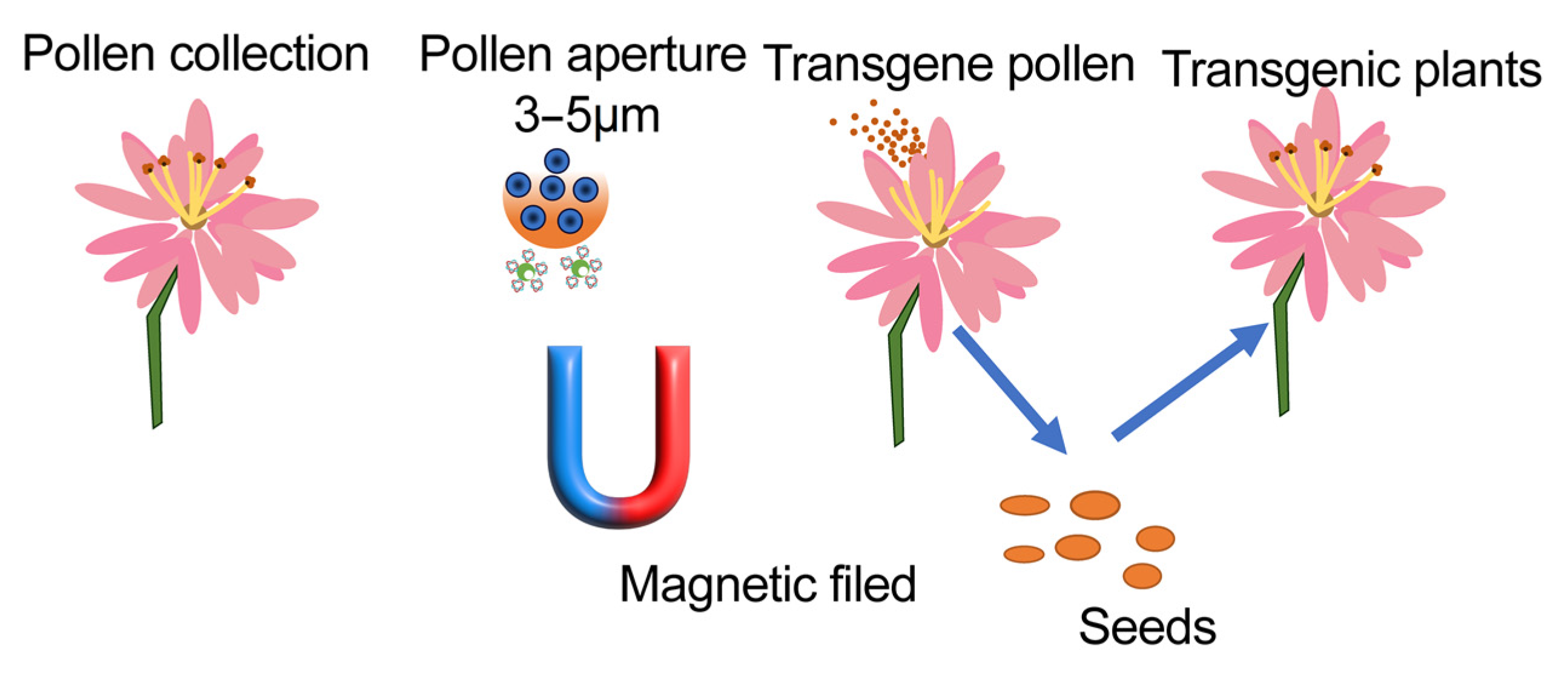
- Zhao, X.; Meng, Z.; Wang, Y.; Chen, W.; Sun, C.; Cui, B.; Cui, J.; Yu, M.; Zeng, Z.; Guo, S.; et al. Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nat. Plants 2017, 3, 956–964. [Google Scholar] [CrossRef]
- Wang, Z.-P.; Zhang, Z.-B.; Zheng, D.-Y.; Zhang, T.-T.; Li, X.-L.; Zhang, C.; Yu, R.; Wei, J.-H.; Wu, Z.-Y. Efficient and genotype independent maize transformation using pollen transfected by DNA-coated magnetic nanoparticles. J. Integr. Plant Biol. 2022, 64, 1145–1156. [Google Scholar] [CrossRef]
- Kwak, S.-Y.; Lew, T.T.S.; Sweeney, C.J.; Koman, V.B.; Wong, M.H.; Bohmert-Tatarev, K.; Snell, K.D.; Seo, J.S.; Chua, N.-H.; Strano, M.S. Chloroplast-selective gene delivery and expression in planta using chitosan-complexed single-walled carbon nanotube carriers. Nat. Nanotechnol. 2019, 14, 447–455. [Google Scholar] [CrossRef]
- Demirer, G.S.; Landry, M.P. Efficient Transient Gene Knock-down in Tobacco Plants Using Carbon Nanocarriers. Bio-Protocol 2021, 11, e3897. [Google Scholar] [CrossRef]
- Kelich, P.; Jeong, S.; Navarro, N.; Adams, J.; Sun, X.; Zhao, H.; Landry, M.P.; Vuković, L. Discovery of DNA–Carbon Nanotube Sensors for Serotonin with Machine Learning and Near-infrared Fluorescence Spectroscopy. ACS Nano 2022, 16, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gu, Z.; Kurniawan, N.; Chen, W.; Xu, Z.P. Manganese-Based Layered Double Hydroxide Nanoparticles as a T(1) -MRI Contrast Agent with Ultrasensitive pH Response and High Relaxivity. Adv. Mater. (Deerfield Beach Fla.) 2017, 29, 1700373. [Google Scholar] [CrossRef]
- Peng, L.; Mei, X.; He, J.; Xu, J.; Zhang, W.; Liang, R.; Wei, M.; Evans, D.G.; Duan, X. Monolayer Nanosheets with an Extremely High Drug Loading toward Controlled Delivery and Cancer Theranostics. Adv. Mater. 2018, 30, 1707389. [Google Scholar] [CrossRef]
- Bao, W.; Wang, J.; Wang, Q.; O’Hare, D.; Wan, Y. Layered Double Hydroxide Nanotransporter for Molecule Delivery to Intact Plant Cells. Sci. Rep. 2016, 6, 26738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect. Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taochy, C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Liu, Q.; Li, Y.; Xu, K.; Li, D.; Hu, H.; Zhou, F.; Song, P.; Yu, Y.; Wei, Q.; Liu, Q.; et al. Clay nanosheet-mediated delivery of recombinant plasmids expressing artificial miRNAs via leaf spray to prevent infection by plant DNA viruses. Hortic. Res. 2020, 7, 179. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, M.; Song, Y.; Li, H.; Huang, H.; Shao, M.; Liu, Y.; Kang, Z. Carbon dots promote the growth and photosynthesis of mung bean sprouts. Carbon 2018, 136, 94–102. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Hendrix, B.; Hoffer, P.; Sanders, R.A.; Zheng, W. Carbon Dots for Efficient Small Interfering RNA Delivery and Gene Silencing in Plants. Plant Physiol. 2020, 184, 647–657. [Google Scholar] [CrossRef]
- Wang, B.; Huang, J.; Zhang, M.; Wang, Y.; Wang, H.; Ma, Y.; Zhao, X.; Wang, X.; Liu, C.; Huang, H.; et al. Carbon Dots Enable Efficient Delivery of Functional DNA in Plants. ACS Appl. Bio Mater. 2020, 3, 8857–8864. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.-H.; Huang, Y.-F.; Zhang, C.-Z.; Niu, J.; Chen, Y.; Chu, Y.; Jiang, Z.-H.; Gao, J.-Q.; Mao, Z.-W. Integration of antimicrobial peptides with gold nanoparticles as unique non-viral vectors for gene delivery to mesenchymal stem cells with antibacterial activity. Biomaterials 2016, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Rudokas, M.; Najlah, M.; Alhnan, M.A.; Elhissi, A. Liposome Delivery Systems for Inhalation: A Critical Review Highlighting Formulation Issues and Anticancer Applications. Med. Princ. Pract. 2016, 2, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Li, X. Liposomal delivery of CRISPR/Cas9. Cancer Gene Ther. 2020, 27, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Vabbilisetty, P.; Sun, X.L. Liposome surface functionalization based on different anchoring lipids via Staudinger ligation. Org. Biomol. Chem. 2014, 12, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Phogat, N.; Kohl, M.; Uddin, I.; Jahan, A. Chapter 11—Interaction of Nanoparticles With Biomolecules, Protein, Enzymes, and Its Applications. In Precision Medicine; Deigner, H.-P., Kohl, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 253–276. [Google Scholar]
- Karny, A.; Zinger, A.; Kajal, A.; Shainsky-Roitman, J.; Schroeder, A. Therapeutic nanoparticles penetrate leaves and deliver nutrients to agricultural crops. Sci. Rep. 2018, 8, 7589. [Google Scholar] [CrossRef] [Green Version]
- Gregoriadis, G.; Buckland, R.A. Enzyme-containing Liposomes alleviate a Model for Storage Disease. Nature 1973, 244, 170–172. [Google Scholar] [CrossRef]
- Wang, C.Y.; Hughes, K.W.; Huang, L. Improved Cytoplasmic Delivery to Plant Protoplasts via pH-Sensitive Liposomes. Plant Physiol. 1986, 82, 179–184. [Google Scholar] [CrossRef]
- Fukunaga, Y.; Nagata, T.; Takebe, I. Liposome-mediated infection of plant protoplasts with tobacco mosaic virus RNA. Virology 1981, 113, 752–760. [Google Scholar] [CrossRef]
- Nagata, T.; Okada, K.; Takebe, I.; Matsui, C. Delivery of tobacco mosaic virus RNA into plant protoplasts mediated by reverse-phase evaporation vesicles (Liposomes). Mol. Genet. Genom. 1981, 184, 161–165. [Google Scholar] [CrossRef]
- Fraley, R.T.; Dellaporta, S.L.; Papahadjopoulos, D. Liposome-mediated delivery of tobacco mosaic virus RNA into tobacco protoplasts: A sensitive assay for monitoring liposome-protoplast interactions. Plant Mol. Biol. 1982, 79, 1859–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, T.; Okada, K.; Takebe, I. Mitotic protoplasts and their infection with tobacco mosaic virus RNA encapsulated in liposomes. Plant Cell Rep. 1982, 1, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Cutler, A.J.; Constabel, F.; Kurz, W.G.; Shargool, P.D. Quantitation of the delivery of liposome contents into plant protoplasts. Anal. Biochem. 1984, 139, 482–486. [Google Scholar] [CrossRef]
- Gad, A.E.; Zeewi, B.-Z.; Altman, A. Fusion of germinating watermelon pollen tubes with liposomes. Plant Sci. 1988, 55, 69–75. [Google Scholar] [CrossRef]
- Cassells, A.C. Uptake of charged lipid vesicles by isolated tomato protoplasts. Nature 1978, 275, 760. [Google Scholar] [CrossRef]
- Ahokas, H. Transfection by DNA-associated liposomes evidenced at pea pollination. Hereditas 1987, 106, 129–138. [Google Scholar] [CrossRef]
- Rosenberg, N.; Gad, A.E.; Altman, A.; Navot, N.; Czosnek, H. Liposome-mediated introduction of the chloramphenicol acetyl transferase (CAT) gene and its expression in tobacco protoplasts. Plant Mol. Biol. 1988, 10, 185–191. [Google Scholar] [CrossRef]
- Rosenberg, N.; Dekel-Reichenbach, M.; Navot, N.; Gad, A.E.; Altman, A.; Czosnek, H. Liposome-mediated introduction of DNA into plant protoplasts and calli. Physiol. Plant 1990, 509–516. [Google Scholar] [CrossRef]
- Liu, J.; Wang, F.-H.; Wang, L.-L.; Xiao, S.-Y.; Tong, C.-Y.; Tang, D.-Y.; Liu, X.-M. Preparation of fluorescence starch-nanoparticle and its application as plant transgenic vehicle. J. Cent. South Univ. 2008, 15, 768–773. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, H.; Demirer, G.S.; González-Grandío, E.; Fan, C.; Landry, M.P. Engineering DNA nanostructures for siRNA delivery in plants. Nat. Protoc. 2020, 15, 3064–3087. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Goyal, V. Agrobacterium-mediated gene transfer in plants and biosafety considerations. Appl. Biochem. Biotechnol. 2012, 168, 1953–1975. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.H. Fundamentals, Properties, and Applications of Polymer Nanocomposites; Cambridge University Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Ali, Z.; Serag, M.F.; Demirer, G.S.; Torre, B.; di Fabrizio, E.; Landry, M.P.; Habuchi, S.; Mahfouz, M. DNA–Carbon Nanotube Binding Mode Determines the Efficiency of Carbon Nanotube-Mediated DNA Delivery to Intact Plants. ACS Appl. Nano Mater. 2022, 5, 4663–4676. [Google Scholar] [CrossRef]
- Bao, A.; Burritt, D.J.; Chen, H.; Zhou, X.; Cao, D.; Tran, L.P. The CRISPR/Cas9 system and its applications in crop genome editing. Crit. Rev. Biotechnol. 2019, 39, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Kawasaki, S.; Abdellatif, I.M.Y.; Nishida, K.; Kondo, A.; Ariizumi, T.; Ezura, H.; Miura, K. Efficient base editing in tomato using a highly expressed transient system. Plant Cell Rep. 2021, 40, 667–676. [Google Scholar] [CrossRef]
- Ren, Q.; Zhong, Z.; Wang, Y.; You, Q.; Li, Q.; Yuan, M.; He, Y.; Qi, C.; Tang, X.; Zheng, X.; et al. Bidirectional Promoter-Based CRISPR-Cas9 Systems for Plant Genome Editing. Front. Plant Sci. 2019, 10, 1173. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Kim, S.G. RPS5A Promoter-Driven Cas9 Produces Heritable Virus-Induced Genome Editing in Nicotiana attenuata. Mol. Cells 2021, 44, 911–919. [Google Scholar] [CrossRef]
- Pan, W.; Liu, X.; Li, D.; Zhang, H. Establishment of an Efficient Genome Editing System in Lettuce Without Sacrificing Specificity. Front. Plant Sci. 2022, 13, 930592. [Google Scholar] [CrossRef]
- Liang, W.; Gong, H.; Yin, D.; Lu, S.; Fu, Q. High-molecular-weight polyethyleneimine conjuncted pluronic for gene transfer agents. Chem. Pharm. Bull. 2011, 59, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Hashemzadeh, I.; Hasanzadeh, A.; Radmanesh, F.; Khodadadi Chegeni, B.; Hosseini, E.S.; Kiani, J.; Shahbazi, A.; Naseri, M.; Fatahi, Y.; Nourizadeh, H.; et al. Polyethylenimine-Functionalized Carbon Dots for Delivery of CRISPR/Cas9 Complexes. ACS Appl. Bio Mater. 2021, 4, 7979–7992. [Google Scholar] [CrossRef]
- Demirer, G.S.; Silva, T.N.; Jackson, C.T.; Thomas, J.B.; Ehrhardt, D.W.; Rhee, S.Y.; Mortimer, J.C.; Landry, M.P. Nanotechnology to advance CRISPR–Cas genetic engineering of plants. Nat. Nanotechnol. 2021, 16, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Kim, J.-K. Transgenic Breeding Approaches for Improving Abiotic Stress Tolerance: Recent Progress and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, M.F.; Nasti, R.A.; Vollbrecht, M.; Starker, C.G.; Clark, M.D.; Voytas, D.F. Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 2020, 38, 84–89. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, X.B.; Lv, Y.; Gong, L.; Wang, R.; Zhu, X.; Yang, R.; Tan, W. Functional DNA-containing nanomaterials: Cellular applications in biosensing, imaging, and targeted therapy. Acc. Chem. Res. 2014, 47, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Meade, B.R.; Chang, Y.-C.; Fredrickson, C.T.; Willert, K.; Puri, N.; Dowdy, S.F. Efficient siRNA delivery into primary cells by a peptide transduction domain–dsRNA binding domain fusion protein. Nat. Biotechnol. 2009, 27, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Conboy, M.; Park, H.M.; Jiang, F.; Kim, H.J.; Dewitt, M.A.; Mackley, V.A.; Chang, K.; Rao, A.; Skinner, C.; et al. Nanoparticle delivery of Cas9 ribonucleoprotein and donor DNA in vivo induces homology-directed DNA repair. Nat. Biomed. Eng. 2017, 1, 889–901. [Google Scholar] [CrossRef] [Green Version]




| Materials-Vectors | Cargos | Plants | Cell Types | Delivery Methods | Ref. |
|---|---|---|---|---|---|
| Silicon carbide fibers | Plasmid (contains Bar and GUS) | N. tabacum, maize, rice, ryegrass, and cotton | Cells suspension, callus | Co-culture | [63,64,65] |
| Gold functional MSNs | Plasmid (GFP gene) | N. tabacum, maize, white onion | Mesophyll protoplasts, epidermis cells | Gene guns | [68,70] |
| Functional MSNs | Plasmid (GUS gene) | Tomato | Epidermis cells | Spraying or injection | [69] |
| Magnetic NPs | Plasmid (GFP, GUS and Bt gene) | Cotton, lily, maize | Pollen cells | Magnetic field | [72,73] |
| Layer double hydroxides | dsRNA for RNAi | Cowpea, A. thaliana, N. tabacum, N. benthamiana, S. lycopersicum, | Mature leaves | Spraying | [81,82] |
| tomato | Pollen cells | Co-culture | [54] | ||
| SWNT/MWNT | Plasmid (GFP, YFP) siRNA for RNAi | N. benthamiana, E. sativa, arugula, A. thaliana | Mature leaves, protoplast and chloroplast | injection without needle or co-culture | [58,74,75] |
| Carbon dots | Plasmids (GFP, GUS, hygromycin resistance gene); siRNA for RNAi | Wheat, rice, tomato | Mature leaves, callus | Spraying/ vacuum assisted | [84,85] |
| Different shapes of AuNPs or magnetic | Plasmid (GFP); siRNA for RNAi | N. benthamiana | Mature leaves | Injection without needle | [60,71] |
| DNA-nanostructure | siRNA for RNAi | N. benthamiana | Mature leaves | Injection without needle | [105] |
| Liposomes | Plasmid of DNA (acetyl transferase) TMV-RNA | Watermelon, Tobacco, Vinca, Petunia, Pea | Pollen cells Protoplast | Co-culture | [91,92,93,94,95,96,97,98,99,100,101,102,103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, H.; Zhou, S.; Pan, W.; Shang, Y.; Zeng, Z.; Zhang, H. The Promising Nanovectors for Gene Delivery in Plant Genome Engineering. Int. J. Mol. Sci. 2022, 23, 8501. https://doi.org/10.3390/ijms23158501
Zhi H, Zhou S, Pan W, Shang Y, Zeng Z, Zhang H. The Promising Nanovectors for Gene Delivery in Plant Genome Engineering. International Journal of Molecular Sciences. 2022; 23(15):8501. https://doi.org/10.3390/ijms23158501
Chicago/Turabian StyleZhi, Heng, Shengen Zhou, Wenbo Pan, Yun Shang, Zhanghua Zeng, and Huawei Zhang. 2022. "The Promising Nanovectors for Gene Delivery in Plant Genome Engineering" International Journal of Molecular Sciences 23, no. 15: 8501. https://doi.org/10.3390/ijms23158501