
In Vivo Bio-Activation of JWH-175 to JWH-018: Pharmacodynamic and Pharmacokinetic Studies in Mice

 ,
,  , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Results
2.1. Affinity and Potency of JWH-175 for CB1 and CB2 Receptors in Comparison to JWH-018
2.2. Effects of JWH-175 on Synaptic Transmission in CA1 Hippocampal Area
2.3. In Vivo Behavioral Studies
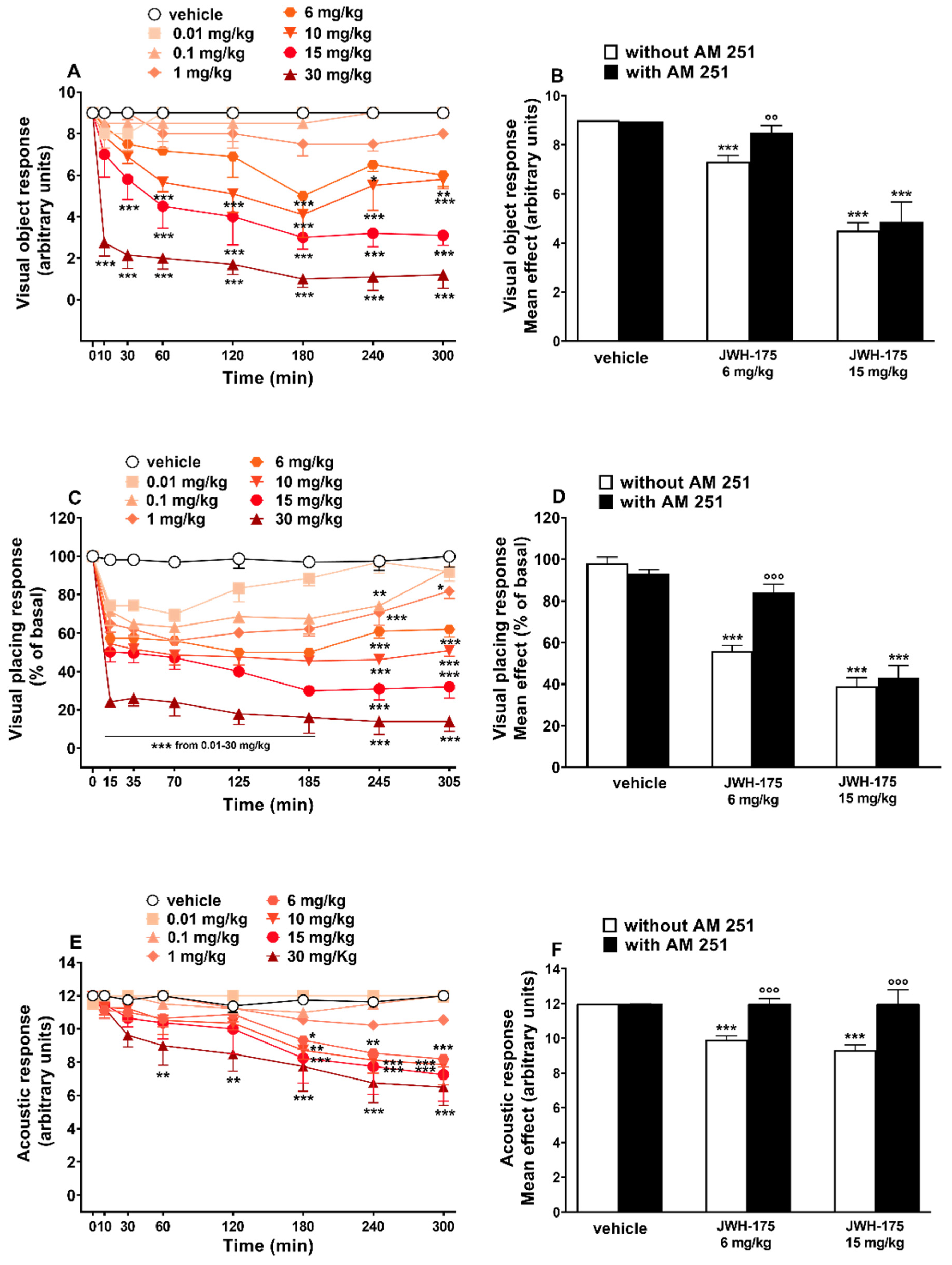
2.3.1. Evaluation of the Visual Object Response
2.3.2. Evaluation of the Visual Placing Response
2.3.3. Evaluation of the Acoustic Response
2.3.4. Evaluation of Breath Rate
2.3.5. Evaluation of Core Temperature
2.3.6. Evaluation of Pain Induced by a Mechanical Stimulus
2.3.7. Evaluation of Number of Steps
2.3.8. Results of Metabolic and Behavioral Studies
Plasmatic Profile and Correlations with Behavioral Responses
Urinary Excretion
3. Discussion
3.1. Behavioral Studies
3.2. JWH-175 and JWH-018 Profile in Plasma
4. Materials and Methods
4.1. Animals
4.2. Drug Preparation and Dose Selection
4.3. In Vitro Studies
4.3.1. Mouse Brain and Spleen Membrane Preparation
4.3.2. Cell Culture and Membrane Preparation
4.3.3. [3H] CP-55,940 Competition Binding Assays
4.3.4. Cyclic AMP Assays
4.3.5. Data Analysis
4.4. In Vitro Electrophysiological Studies in Hippocampal Slices
4.4.1. Tissue Preparation
4.4.2. Electrophysiological Recording
4.4.3. Data Analysis
4.5. In Vivo Behavioral Studies
4.5.1. Evaluation of the Visual Responses
4.5.2. Evaluation of the Acoustic Response
4.5.3. Evaluation of Breath Rate
4.5.4. Evaluation of Core Temperature
4.5.5. Evaluation of Pain Induced by Mechanical Stimulation of the Tail
4.5.6. Drag Test
4.6. Statistical Analysis
4.7. In Vitro and In Vivo Metabolic Studies
4.7.1. Chemicals
4.7.2. Protocol for the In Vitro Formation of the JWH-018 Phase I Metabolites and for the Characterization of the Enzymatic Isoforms Involved
4.7.3. Protocol for the In Vivo Pharmacokinetic Studies and Behavioral Correlation
Sample Collection
Sample Pre-Treatment
4.7.4. Instrumental Conditions
4.7.5. LC-MS/MS Method Validation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Statements
Abbreviations
| AM 251 | 1-(2,4-dichlorophenyl)-5-(4-iodophenyl)-4-methyl-N-(piperidin-1-yl)-1H-pyrazole-3-carboxamide |
| CB1R | Cannabinoid receptor 1 |
| JWH-018 | 1-naphthalenyl(1-pentyl-1H-indol-3-yl)-methanone |
| JWH-175 | 3-(1-naphthalenylmethyl)-1-pentyl-1H-indole |
| NPSs | Novel psychoactive substances |
| SCBs | Synthetic cannabinoids |
References
- United Nations Office on Drugs and Crime (UNODC). Synthetic Cannabinoids, UNODC Laboratory and Scientific Service Portals. 2020. Available online: https://www.unodc.org/LSS/SubstanceGroup/Details/ae45ce06-6d33-4f5f-916a-e873f07bde02 (accessed on 2 February 2022).
- Fattore, L.; Fratta, W. Beyond THC: The New Generation of Cannabinoid Designer Drugs. Front. Behav. Neurosci. 2011, 21, 5–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawilska, J.B.; Wojcieszak, J. Spice/K2 drugs—more than innocent substitutes for marijuana. Int. J. Neuropsychopharmacol. 2014, 17, 509–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsohly, M.A.; Gul, W.; Wanas, A.S.; Radwan, M.M. Synthetic cannabinoids: Analysis and metabolites. Life Sci. 2014, 97, 78–90. [Google Scholar] [CrossRef]
- Kemp, A.M.; Clark, M.S.; Dobbs, T.; Galli, R.; Sherman, J.; Cox, R. Top 10 Facts You Need to Know about Synthetic Cannabinoids: Not So Nice Spice. Am. J. Med. 2016, 129, 240–244.e1. [Google Scholar] [CrossRef] [PubMed]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). New Psychoactive Substances: Global Markets, Glocal Threats and the COVID-19 Pandemic; An update from the EU Early Warning System; Publications Office of the European Union: Luxembourg, 2020.
- Tait, R.J.; Caldicott, D.; Mountain, D.; Hill, S.L.; Lenton, S. A systematic review of adverse events arising from the use of synthetic cannabinoids and their associated treatment. Clin. Toxicol. 2016, 54, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Obafemi, A.I.; Kleinschmidt, K.; Goto, C.; Fout, D. Cluster of Acute Toxicity from Ingestion of Synthetic Cannabinoid-Laced Brownies. J. Med. Toxicol. 2015, 11, 426–429. [Google Scholar] [CrossRef] [Green Version]
- Le Boisselier, R.; Alexandre, J.; Lelong-Boulouard, V.; Debruyne, D. Focus on cannabinoids and synthetic cannabinoids. Clin. Pharmacol. Ther. 2017, 101, 220–229. [Google Scholar] [CrossRef]
- Monte, A.A.; Calello, D.P.; Gerona, R.R.; Hamad, E.; Campleman, S.L.; Brent, J.; Wax, P.; Carlson, R.G. ACMT Toxicology Investigators Consortium (ToxIC). Characteristics and Treatment of Patients with Clinical Illness Due to Synthetic Cannabinoid Inhalation Reported by Medical Toxicologists: A ToxIC Database Study. J. Med. Toxicol. 2017, 13, 146–152. [Google Scholar] [CrossRef]
- Deng, H.; Verrico, C.D.; Kosten, T.R.; Nielsen, D.A. Psychosis and synthetic cannabinoids. Psychiatry Res. 2018, 268, 400–412. [Google Scholar] [CrossRef]
- Mensen, V.T.; Vreeker, A.; Nordgren, J.; Atkinson, A.; de la Torre, R.; Farré, M.; Ramaekers, J.G.; Brunt, T.M. Psychopathological symptoms associated with synthetic cannabinoid use: A comparison with natural cannabis. Psychopharmacology 2019, 236, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Lenzi, M.; Cocchi, V.; Cavazza, L.; Bilel, S.; Hrelia, P.; Marti, M. Genotoxic Properties of Synthetic Cannabinoids on TK6 Human Cells by Flow Cytometry. Int. J. Mol. Sci. 2020, 21, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coccini, T.; De Simone, U.; Lonati, D.; Scaravaggi, G.; Marti, M.; Locatelli, C.A. MAM-2201, One of the Most Potent-Naphthoyl Indole Derivative-Synthetic Cannabinoids, Exerts Toxic Effects on Human Cell-Based Models of Neurons and Astrocytes. Neurotox. Res. 2021, 39, 1251–1273. [Google Scholar] [CrossRef] [PubMed]
- Canazza, I.; Ossato, A.; Trapella, C.; Fantinati, A.; De Luca, M.A.; Margiani, G.; Vincenzi, F.; Rimondo, C.; Di Rosa, F.; Gregori, A.; et al. Effect of the novel synthetic cannabinoids AKB48 and 5F-AKB48 on “tetrad”, sensorimotor, neurological and neurochemical responses in mice. In vitro and in vivo pharmacological studies. Psychopharmacology 2016, 233, 3685–3709. [Google Scholar] [CrossRef] [PubMed]
- Wouters, E.; Walraed, J.; Robertson, M.J.; Meyrath, M.; Szpakowska, M.; Chevigné, A.; Skiniotis, G.; Stove, C. Assessment of Biased Agonism among Distinct Synthetic Cannabinoid Receptor Agonist Scaffolds. ACS Pharmacol. Transl. Sci. 2019, 3, 285–295. [Google Scholar] [CrossRef]
- Tai, S.; Fantegrossi, W.E. Pharmacological and Toxicological Effects of Synthetic Cannabinoids and Their Metabolites. Curr. Top. Behav. Neurosci. 2017, 32, 249–262. [Google Scholar] [CrossRef]
- Brents, L.K.; Reichard, E.E.; Zimmerman, S.M.; Moran, J.H.; Fantegrossi, W.E.; Prather, P.L. Phase I hydroxylated metabolites of the K2 synthetic cannabinoid JWH-018 retain in vitro and in vivo cannabinoid 1 receptor affinity and activity. PLoS ONE. 2011, 6, e21917. [Google Scholar] [CrossRef]
- Longworth, M.; Connor, M.; Banister, S.D.; Kassiou, M. Synthesis and Pharmacological Profiling of the Metabolites of Synthetic Cannabinoid Drugs APICA, STS-135, ADB-PINACA, and 5F-ADB-PINACA. ACS Chem. Neurosci. 2017, 8, 1673–1680. [Google Scholar] [CrossRef]
- Hutchison, R.D.; Ford, B.M.; Franks, L.N.; Wilson, C.D.; Yarbrough, A.L.; Fujiwara, R.; Su, M.K.; Fernandez, D.; James, L.P.; Moran, J.H.; et al. Atypical Pharmacodynamic Properties and Metabolic Profile of the Abused Synthetic Cannabinoid AB-PINACA: Potential Contribution to Pronounced Adverse Effects Relative to Δ9-THC. Front. Pharmacol. 2018, 9, 1084. [Google Scholar] [CrossRef] [Green Version]
- Pinson, A.; Yarbrough, A.L.; Bush, J.M.; Cabanlong, C.V.; Shoeib, A.; Jackson, B.K.; Fukuda, S.; Gogoi, J.; Fantegrossi, W.E.; McCain, K.; et al. Metabolism, CB1 cannabinoid receptor binding and in vivo activity of synthetic cannabinoid 5F-AKB48: Implications for toxicity. Pharmacol. Biochem. Behav. 2020, 195, 172949. [Google Scholar] [CrossRef]
- Rajasekaran, M.; Brents, L.K.; Franks, L.N.; Moran, J.H.; Prather, P.L. Human metabolites of synthetic cannabinoids JWH-018 and JWH-073 bind with high affinity and act as potent agonists at cannabinoid type-2 receptors. Toxicol. Appl. Pharmacol. 2013, 269, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Couceiro, J.; Bandarra, S.; Sultan, H.; Bell, S.; Constantino, S.; Quintas, A. Toxicological impact of JWH-018 and its phase I metabolite N-(3-hydroxypentyl) on human cell lines. Forensic Sci. Int. 2016, 264, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Zendulka, O.; Dovrtělová, G.; Nosková, K.; Turjap, M.; Šulcová, A.; Hanuš, L.; Juřica, J. Cannabinoids and Cytochrome P450 Interactions. Curr. Drug Metab. 2016, 17, 206–226. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.Y.; Kim, J.H.; Kim, D.K.; Lee, H.S. Synthetic cannabinoids are substrates and inhibitors of multiple drug-metabolizing enzymes. Arch. Pharm. Res. 2018, 41, 691–710. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, D.K.; Shin, Y.; Jeon, J.H.; Song, I.S.; Lee, H.S. In Vitro Interaction of AB-FUBINACA with Human Cytochrome P450, UDP-Glucuronosyltransferase Enzymes and Drug Transporters. Molecules 2020, 25, 4589. [Google Scholar] [CrossRef]
- Huffman, J.W.; Mabon, R.; Wu, M.J.; Lu, J.; Hart, R.; Hurst, D.P.; Reggio, P.H.; Wiley, J.L.; Martin, B.R. 3-Indolyl-1-naphthylmethanes: New cannabimimetic indoles provide evidence for aromatic stacking interactions with the CB(1) cannabinoid receptor. Bioorg. Med. Chem. 2003, 11, 539–549. [Google Scholar] [CrossRef]
- Huffman, J.W.; Padgett, L.W. Recent developments in the medicinal chemistry of cannabimimetic indoles, pyrroles and indenes. Curr. Med. Chem. 2005, 12, 1395–1411. [Google Scholar] [CrossRef]
- Tampus, R.; Yoon, S.S.; de la Peña, J.B.; Botanas, C.J.; Kim, H.J.; Seo, J.W.; Jeong, E.J.; Jang, C.G.; Cheong, J.H. Assessment of the Abuse Liability of Synthetic Cannabinoid Agonists JWH-030, JWH-175, and JWH-176. Biomol. Ther. 2015, 23, 590–596. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, K.F.; Oliveira, K.D.; Huestis, M.A.; Costa, J.L. Screening of 104 New Psychoactive Substances (NPS) and Other Drugs of Abuse in Oral Fluid by LC-MS-MS. J. Anal. Toxicol. 2020, 44, 697–707. [Google Scholar] [CrossRef]
- Fietzke, M.; Thomas, A.; Beike, J.; Rothschild, M.A.; Thevis, M.; Mercer-Chalmers-Bender, K. In vitro elucidation of the metabolic profile of the synthetic cannabinoid receptor agonists JWH-175 and JWH-176. Forensic Toxicol. 2016, 34, 353–362. [Google Scholar] [CrossRef]
- Psychonaut, W. JWH-018. Available online: https://psychonautwiki.org/wiki/JWH-018 (accessed on 21 June 2022).
- Vigolo, A.; Ossato, A.; Trapella, C.; Vincenzi, F.; Rimondo, C.; Seri, C.; Varani, K.; Serpelloni, G.; Marti, M. Novel halogenated derivates of JWH-018: Behavioral and binding studies in mice. Neuropharmacology 2015, 95, 68–82. [Google Scholar] [CrossRef]
- Ossato, A.; Vigolo, A.; Trapella, C.; Seri, C.; Rimondo, C.; Serpelloni, G.; Marti, M. JWH-018 impairs sensorimotor functions in mice. Neuroscience 2015, 300, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, M.; Ossato, A.; Canazza, I.; Trapella, C.; Borelli, A.C.; Beggiato, S.; Rimondo, C.; Serpelloni, G.; Ferraro, L.; Marti, M. Synthetic cannabinoid JWH-018 and its halogenated derivatives JWH-018-Cl and JWH-018-Br impair Novel Object Recognition in mice: Behavioral, electrophysiological and neurochemical evidence. Neuropharmacology 2016, 109, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.F.; Lycas, M.D.; Kaczmarzyk, J.R.; Spivak, C.E.; Baumann, M.H.; Lupica, C.R. Disruption of hippocampal synaptic transmission and long-term potentiation by psychoactive synthetic cannabinoid ‘Spice’ compounds: Comparison with Δ9-tetrahydrocannabinol. Addict. Biol. 2017, 22, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Nowicky, A.V.; Teyler, T.J.; Vardaris, R.M. The modulation of long-term potentiation by delta-9-tetrahydrocannabinol in the rat hippocampus, in vitro. Brain Res. Bull. 1987, 19, 663–672. [Google Scholar] [CrossRef]
- Collins, D.R.; Pertwee, R.G.; Davies, S.N. The action of synthetic cannabinoids on the induction of long-term potentiation in the rat hippocampal slice. Eur. J. Pharmacol. 1994, 259, R7–R8. [Google Scholar] [CrossRef]
- Terranova, J.P.; Michaud, J.C.; Le Fur, G.; Soubrié, P. Inhibition of long-term potentiation in rat hippocampal slices by anandamide and WIN55212-2: Reversal by SR141716 A, a selective antagonist of CB1 cannabinoid receptors. Naunyn Schmiedebergs Arch. Pharmacol. 1995, 352, 576–579. [Google Scholar] [CrossRef]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 88, 773–778. [Google Scholar] [CrossRef] [Green Version]
- Hájos, N.; Ledent, C.; Freund, T.F. Novel cannabinoid-sensitive receptor mediates inhibition of glutamatergic synaptic transmission in the hippocampus. Neuroscience 2001, 106, 1–4. [Google Scholar] [CrossRef]
- Diana, G.; Malloni, M.; Pieri, M. Effects of the synthetic cannabinoid nabilone on spatial learning and hippocampal neurotransmission. Pharmacol. Biochem. Behav. 2003, 75, 585–591. [Google Scholar] [CrossRef]
- Domenici, M.R.; Azad, S.C.; Marsicano, G.; Schierloh, A.; Wotjak, C.T.; Dodt, H.U.; Zieglgänsberger, W.; Lutz, B.; Rammes, G. Cannabinoid receptor type 1 located on presynaptic terminals of principal neurons in the forebrain controls glutamatergic synaptic transmission. J. Neurosci. 2006, 26, 5794–5799. [Google Scholar] [CrossRef] [Green Version]
- Péterfi, Z.; Urbán, G.M.; Papp, O.I.; Németh, B.; Monyer, H.; Szabó, G.; Erdélyi, F.; Mackie, K.; Freund, T.F.; Hájos, N.; et al. Endocannabinoid-mediated long-term depression of afferent excitatory synapses in hippocampal pyramidal cells and GABAergic interneurons. J. Neurosci. 2012, 32, 14448–14463. [Google Scholar] [CrossRef] [PubMed]
- Ossato, A.; Canazza, I.; Trapella, C.; Vincenzi, F.; De Luca, M.A.; Rimondo, C.; Varani, K.; Borea, P.A.; Serpelloni, G.; Marti, M. Effect of JWH-250, JWH-073 and their interaction on “tetrad”, sensorimotor, neurological and neurochemical responses in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 67, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Bilel, S.; Tirri, M.; Arfè, R.; Ossato, A.; Trapella, C.; Serpelloni, G.; Neri, M.; Fattore, L.; Marti, M. Novel halogenated synthetic cannabinoids impair sensorimotor functions in mice. Neurotoxicology 2020, 76, 17–32. [Google Scholar] [CrossRef]
- Wiebelhaus, J.M.; Poklis, J.L.; Poklis, A.; Vann, R.E.; Lichtman, A.H.; Wise, L.E. Inhalation exposure to smoke from synthetic “marijuana” produces potent cannabimimetic effects in mice. Drug Alcohol Depend. 2012, 126, 316–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, J.L.; Marusich, J.A.; Huffman, J.W. Moving around the molecule: Relationship between chemical structure and in vivo activity of synthetic cannabinoids. Life Sci. 2014, 97, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macrì, S.; Lanuzza, L.; Merola, G.; Ceci, C.; Gentili, S.; Valli, A.; Macchia, T.; Laviola, G. Behavioral responses to acute and sub-chronic administration of the synthetic cannabinoid JWH-018 in adult mice prenatally exposed to corticosterone. Neurotox Res. 2013, 24, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Schifano, F.; Orsolini, L.; Duccio Papanti, G.; Corkery, J.M. Novel psychoactive substances of interest for psychiatry. World Psychiatry 2015, 14, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Bilel, S.; Tirri, M.; Arfè, R.; Stopponi, S.; Soverchia, L.; Ciccocioppo, R.; Frisoni, P.; Strano-Rossi, S.; Miliano, C.; De-Giorgio, F.; et al. Pharmacological and Behavioral Effects of the Synthetic Cannabinoid AKB48 in Rats. Front. Neurosci. 2019, 13, 1163. [Google Scholar] [CrossRef]
- Tsou, K.; Brown, S.; Sañudo-Peña, M.C.; Mackie, K.; Walker, J.M. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 1998, 83, 393–411. [Google Scholar] [CrossRef]
- Dasilva, M.A.; Grieve, K.L.; Cudeiro, J.; Rivadulla, C. Endocannabinoid CB1 receptors modulate visual output from the thalamus. Psychopharmacology 2012, 219, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, T.; Kameyama, K.; Esumi, K.; Daimyo, Y.; Watanabe, M.; Hata, Y. Developmental and visual input-dependent regulation of the CB1 cannabinoid receptor in the mouse visual cortex. PLoS ONE 2013, 8, e53082. [Google Scholar] [CrossRef] [PubMed]
- Reig, R.; Silberberg, G. Multisensory integration in the mouse striatum. Neuron 2014, 83, 1200–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järvinen, T.; Pate, D.W.; Laine, K. Cannabinoids in the treatment of glaucoma. Pharmacol. Ther. 2002, 95, 203–220. [Google Scholar] [CrossRef]
- Gómez-Nieto, R.; Horta-Júnior Jde, A.; Castellano, O.; Millian-Morell, L.; Rubio, M.E.; López, D.E. Origin and function of short-latency inputs to the neural substrates underlying the acoustic startle reflex. Front. Neurosci. 2014, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Tzounopoulos, T.; Rubio, M.E.; Keen, J.E.; Trussell, L.O. Coactivation of pre- and postsynaptic signaling mechanisms determines cell-specific spike-timing-dependent plasticity. Neuron 2007, 54, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.D.; Li, D.M. Cardiovascular and respiratory effects of cannabis in cat and rat. Br. J. Pharmacol. 1973, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rosenkrantz, H.; Heyman, I.A.; Braude, M.C. Inhalation, parenteral and oral LD50 values of delta 9-tetrahydrocannabinol in Fischer rats. Toxicol. Appl. Pharmacol. 1974, 28, 18–27. [Google Scholar] [CrossRef]
- Phillips, R.N.; Turk, R.F.; Forney, R.B. Acute toxicity of delta-9-tetrahydrocannabinol in rats and mice. Proc. Soc. Exp. Biol. Med. 1971, 136, 260–263. [Google Scholar] [CrossRef]
- Schmid, K.; Niederhoffer, N.; Szabo, B. Analysis of the respiratory effects of cannabinoids in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 368, 301–308. [Google Scholar] [CrossRef]
- Pfitzer, T.; Niederhoffer, N.; Szabo, B. Central effects of the cannabinoid receptor agonist WIN55212-2 on respiratory and cardiovascular regulation in anaesthetised rats. Br. J. Pharmacol. 2004, 142, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Calignano, A.; Kátona, I.; Désarnaud, F.; Giuffrida, A.; La Rana, G.; Mackie, K.; Freund, T.F.; Piomelli, D. Bidirectional control of airway responsiveness by endogenous cannabinoids. Nature 2000, 408, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.W.; Gramling, B.R.; Justinova, Z.; Thorndike, E.B.; Baumann, M.H. Synthetic cannabinoids found in “spice” products alter body temperature and cardiovascular parameters in conscious male rats. Drug Alcohol Depend. 2017, 179, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.S.; McMahon, L.R. JWH-018 in rhesus monkeys: Differential antagonism of discriminative stimulus, rate-decreasing, and hypothermic effects. Eur. J. Pharmacol. 2014, 740, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawls, S.M.; Cabassa, J.; Geller, E.B.; Adler, M.W. CB1 receptors in the preoptic anterior hypothalamus regulate WIN 55212-2 [(4,5-dihydro-2-methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo[3,2,1ij]quinolin-6-one]-inducedhypothermia. J. Pharmacol. Exp. Ther. 2002, 301, 963–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovadia, H.; Wohlman, A.; Mechoulam, R.; Weidenfeld, J. Characterization of the hypothermic effect of the synthetic cannabinoid HU-210 in the rat. Relation to the adrenergic system and endogenous pyrogens. Neuropharmacology 1995, 34, 175–180. [Google Scholar] [CrossRef]
- Schreiber, S.; Bader, M.; Lenchinski, T.; Meningher, I.; Rubovitch, V.; Katz, Y.; Cohen, E.; Gabet, Y.; Rotenberg, M.; Wolf, E.U.; et al. Functional effects of synthetic cannabinoids versus Δ9-THC in mice on body temperature, nociceptive threshold, anxiety, cognition, locomotor/exploratory parameters and depression. Addict. Biol. 2019, 24, 414–425. [Google Scholar] [CrossRef]
- Martin, W.J.; Hohmann, A.G.; Walker, J.M. Suppression of noxious stimulus-evoked activity in the ventral posterolateral nucleus of the thalamus by a cannabinoid agonist: Correlation between electrophysiological and antinociceptive effects. J. Neurosci. 1996, 16, 6601–6611. [Google Scholar] [CrossRef] [Green Version]
- Woodhams, S.G.; Chapman, V.; Finn, D.P.; Hohmann, A.G.; Neugebauer, V. The cannabinoid system and pain. Neuropharmacology 2017, 124, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Klinger-Gratz, P.P.; Ralvenius, W.T.; Neumann, E.; Kato, A.; Nyilas, R.; Lele, Z.; Katona, I.; Zeilhofer, H.U. Acetaminophen Relieves Inflammatory Pain through CB1 Cannabinoid Receptors in the Rostral Ventromedial Medulla. J. Neurosci. 2018, 38, 322–334. [Google Scholar] [CrossRef] [Green Version]
- Dogrul, A.; Seyrek, M.; Yalcin, B.; Ulugol, A. Involvement of descending serotonergic and noradrenergic pathways in CB1 receptor-mediated antinociception. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 97–105. [Google Scholar] [CrossRef]
- Yu, X.H.; Cao, C.Q.; Martino, G.; Puma, C.; Morinville, A.; St-Onge, S.; Lessard, É.; Perkins, M.N.; Laird, J.M.A. A peripherally restricted cannabinoid receptor agonist produces robust anti-nociceptive effects in rodent models of inflammatory and neuropathic pain. Pain 2010, 151, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Pacher, P.; Tegeder, I.; Amaya, F.; Constantin, C.E.; Brenner, G.J.; Rubino, T.; Michalski, C.W.; Marsicano, G.; Monory, K.; et al. Cannabinoids mediate analgesia largely via peripheral type 1 cannabinoid receptors in nociceptors. Nat. Neurosci. 2007, 10, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Lichtman, A.H.; Martin, B.R. Cannabinoid-induced antinociception is mediated by a spinal alpha 2-noradrenergic mechanism. Brain Res. 1991, 559, 309–314. [Google Scholar] [CrossRef]
- Sulcova, E.; Mechoulam, R.; Fride, E. Biphasic effects of anandamide. Pharmacol. Biochem. Behav. 1998, 59, 347–352. [Google Scholar] [CrossRef]
- Drews, E.; Schneider, M.; Koch, M. Effects of the cannabinoid receptor agonist WIN 55,212-2 on operant behavior and locomotor activity in rats. Pharmacol. Biochem. Behav. 2005, 80, 145–150. [Google Scholar] [CrossRef]
- Canazza, I.; Ossato, A.; Vincenzi, F.; Gregori, A.; Di Rosa, F.; Nigro, F.; Rimessi, A.; Pinton, P.; Varani, K.; Borea, P.A.; et al. Pharmaco-toxicological effects of the novel third-generation fluorinate synthetic cannabinoids, 5F-ADBINACA, AB-FUBINACA, and STS-135 in mice. In vitro and in vivo studies. Hum. Psychopharmacol. 2017, 32, e2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez de Fonseca, F.; Del Arco, I.; Martín-Calderón, J.L.; Gorriti, M.A.; Navarro, M. Role of the endogenous cannabinoid system in the regulation of motor activity. Neurobiol. Dis. 1998, 5, 483–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossato, A.; Uccelli, L.; Bilel, S.; Canazza, I.; Di Domenico, G.; Pasquali, M.; Pupillo, G.; De Luca, M.A.; Boschi, A.; Vincenzi, F.; et al. Psychostimulant Effect of the Synthetic Cannabinoid JWH-018 and AKB48: Behavioral, Neurochemical, and Dopamine Transporter Scan Imaging Studies in Mice. Front. Psychiatry 2017, 8, 130. [Google Scholar] [CrossRef] [Green Version]
- Funada, M.; Takebayashi-Ohsawa, M.; Tomiyama, K.I. Synthetic cannabinoids enhanced ethanol-induced motor impairments through reduction of central glutamate neurotransmission. Toxicol. Appl. Pharmacol. 2020, 408, 115283. [Google Scholar] [CrossRef]
- Morera-Herreras, T.; Miguelez, C.; Aristieta, A.; Ruiz-Ortega, J.Á.; Ugedo, L. Endocannabinoid modulation of dopaminergic motor circuits. Front. Pharmacol. 2012, 3, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, H.R.; Napier, I.; Connor, M. Inhibition of recombinant human T-type calcium channels by Delta9-tetrahydrocannabinol and cannabidiol. J. Biol. Chem. 2008, 283, 16124–16134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G. Receptors and channels targeted by synthetic cannabinoid receptor agonists and antagonists. Curr. Med. Chem. 2010, 17, 1360–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.F.; Galaj, E.; Bi, G.H.; Zhang, C.; He, Y.; Zhan, J.; Bauman, M.H.; Gardner, E.L.; Xi, Z.X. Different receptor mechanisms underlying phytocannabinoid- versus synthetic cannabinoid-induced tetrad effects: Opposite roles of CB1 /CB2 versus GPR55 receptors. Br. J. Pharmacol. 2020, 177, 1865–1880. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Pongratz, G.; Straub, R.H. The synthetic cannabinoid WIN55,212-2 mesylate decreases the production of inflammatory mediators in rheumatoid arthritis synovial fibroblasts by activating CB2, TRPV1, TRPA1 and yet unidentified receptor targets. J. Inflamm. 2016, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Tirumalai, P.S.; Bhamre, S.; Upadhya, S.C.; Boyd, M.R.; Ravindranath, V. Expression of multiple forms of cytochrome P450 and associated mono-oxygenase activities in rat brain regions. Biochem. Pharmacol. 1998, 56, 371–375. [Google Scholar] [CrossRef]
- Lavandera, J.; Ruspini, S.; Batlle, A.; Buzaleh, A.M. Cytochrome P450 expression in mouse brain: Specific isoenzymes involved in Phase I metabolizing system of porphyrinogenic agents in both microsomes and mitochondria. Biochem. Cell. Biol. 2015, 93, 102–107. [Google Scholar] [CrossRef]
- Stamou, M.; Wu, X.; Kania-Korwel, I.; Lehmler, H.J.; Lein, P.J. Cytochrome p450 mRNA expression in the rodent brain: Species-, sex-, and region-dependent differences. Drug Metab. Dispos. 2014, 42, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, V.; Strobel, H.W. Cytochrome P450-mediated metabolism in brain: Functional roles and their implications. Expert Opin. Drug Metab. Toxicol. 2013, 9, 551–558. [Google Scholar] [CrossRef]
- Vincenzi, F.; Targa, M.; Corciulo, C.; Tabrizi, M.A.; Merighi, S.; Gessi, S.; Saponaro, G.; Baraldi, P.G.; Borea, P.A.; Varani, K. Antinociceptive effects of the selective CB2 agonist MT178 in inflammatory and chronic rodent pain models. Pain 2013, 154, 864–873. [Google Scholar] [CrossRef]
- Anderson, W.W.; Collingridge, G.L. Capabilities of the WinLTP data acquisition program extending beyond basic LTP experimental functions. J. Neurosci. Methods. 2007, 162, 346–356. [Google Scholar] [CrossRef]
- Food and Drug Administration, HHS. International Conference on Harmonisation; Guidance on S7A Safety Pharmacology Studies for Human Pharmaceuticals; Availability. Notice 66 FR 36791. 2001; pp. 36791–36792. Available online: https://www.federalregister.gov/documents/2001/07/13/01-17498/international-conference-on-harmonisation-guidance-on-s7a-safety-pharmacology-studies-for-human (accessed on 1 February 2022).
- Ossato, A.; Bilel, S.; Gregori, A.; Talarico, A.; Trapella, C.; Gaudio, R.M.; De-Giorgio, F.; Tagliaro, F.; Neri, M.; Fattore, L.; et al. Neurological, sensorimotor and cardiorespiratory alterations induced by methoxetamine, ketamine and phencyclidine in mice. Neuropharmacology 2018, 141, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Arfè, R.; Bilel, S.; Tirri, M.; Frisoni, P.; Serpelloni, G.; Neri, M.; Boccuto, F.; Bernardi, T.; Foti, F.; De-Giorgio, F.; et al. Comparison of N-methyl-2-pyrrolidone (NMP) and the “date rape” drug GHB: Behavioral toxicology in the mouse model. Psychopharmacology 2021, 238, 2275–2295. [Google Scholar] [CrossRef] [PubMed]
- Camuto, C.; Pellegrini, S.; De-Giorgio, F.; de la Torre, X.; Marti, M.; Mazzarino, M.; Botrè, F. Urinary excretion profile of methiopropamine in mice following intraperitoneal administration: A liquid chromatography-tandem mass spectrometry investigation. Drug Test. Anal. 2021, 13, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Chieffi, C.; Camuto, C.; De-Giorgio, F.; de la Torre, X.; Diamanti, F.; Mazzarino, M.; Trapella, C.; Marti, M.; Botre, F. Metabolic profile of the synthetic drug 4,4′-dimethylaminorex in urine by LC–MS-based techniques: Selection of the most suitable markers of its intake. Forensic Toxicol. 2021, 39, 89–100. [Google Scholar] [CrossRef]
- Mazzarino, M.; Camuto, C.; Comunità, F.; de la Torre, X.; Stacchini, C.; Botrè, F. Application of liquid chromatography coupled to data-independent acquisition mass spectrometry for the metabolic profiling of N-ethyl heptedrone. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2021, 15, 1185–122989. [Google Scholar] [CrossRef] [PubMed]
- Camuto, C.; Arfè, R.; Tirri, M.; de la Torre, X.; Mazzarino, M.; Marti, M.; De Giorgio, F.; Botrè, F. Urinary excretion and effects on visual placing response in mice of gam-ma-valero-lactone, an alternative to gamma-hydroxy-butyrate for drug-facilitated sexual assault. Emerg. Trends Drugs Addict. Health 2022, 2, 100028. [Google Scholar] [CrossRef]
- Mestria, S.; Odoardi, S.; Federici, S.; Bilel, S.; Tirri, M.; Marti, M.; Strano Rossi, S. Metabolism Study of N-Methyl 2-Aminoindane (NM2AI) and Determination of Metabolites in Biological Samples by LC-HRMS. J. Anal. Toxicol. 2021, 45, 475–483. [Google Scholar] [CrossRef]
- Mazzarino, M.; de la Torre, X.; Botrè, F. A liquid chromatography-mass spectrometry method based on class characteristic fragmentation pathways to detect the class of indole-derivative synthetic cannabinoids in biological samples. Anal. Chim. Acta. 2014, 837, 70–82. [Google Scholar] [CrossRef]
- Bush, D.M.; Woodwell, D.A. Update: Drug-Related Emergency Department Visits Involving Synthetic Cannabinoids. In The CBHSQ Report; Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2013. [Google Scholar] [PubMed]
- Corli, G.; Tirri, M.; Arfè, R.; Bilel, S.; Marchetti, B.; Gregori, A.; Di Rosa, F.; Vincenzi, F.; De-Giorgio, F.; Borea, P.A.; et al. Behavioral and binding studies on the quinolinyl ester indoles 5F-PB22 (5F-QUPIC) and BB-22 (QUCHIC) in the mouse model. Emerg. Trends Drugs Addict. Health. 2022, 2, 100039. [Google Scholar] [CrossRef]
- Fattore, L.; Marti, M.; Mostallino, R.; Castelli, M.P. Sex and Gender Differences in the Effects of Novel Psychoactive Substances. Brain Sci. 2020, 10, 606. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | hCB1 CHO Membranes a Ki (nM) | hCB2 CHO Membranes a Ki (nM) | Mouse Cortex Membranes CB1 a Ki (nM) | Mouse Spleen Membranes CB2 a Ki (nM) | hCB1 CHO Cells b IC50 (nM) | hCB2 CHO Cells b IC50 (nM) |
|---|---|---|---|---|---|---|
| JWH-175 | 25.8 ± 1.9 | 363 ± 31 | 33.6 ± 2.4 | 487 ± 39 | 72 ± 5 | 864 ± 61 |
| JWH-018 * | 9.52 ± 0.73 | 8.63 ± 0.69 | 5.82 ± 0.44 | 7.13 ± 0.53 | 13.69 ± 1.04 | 11.62 ± 0.97 |
| Test | JWH-175 ED50 (mg/kg) | JWH-018 # ED50 (mg/kg) |
|---|---|---|
| Visual Object | 9.6 ± 0.02 *** | 0.32 ± 0.06 |
| Visual Placing | 5.2 ± 0.08 *** | 0.7 ± 0.06 |
| Startle Reflex | 4.0 ± 0.03 *** | 0.53 ± 0.05 |
| Breath Rate | 1.7 ± 0.15 | N/D |
| Core Temperature | 5.6 ± 0.04 *** | 1.7 ± 0.04 |
| Tail Pinch | 4.0 ± 0.12 * | 1.8 ± 0.11 |
| Drag Test | 10.9 ± 0.12 ** | 2.4 ± 0.12 |
| Compound | Precursor Ions (m/z) | Product Ions (m/z) | Collision Energy (eV) |
|---|---|---|---|
| JWH-210 (ISTD) | 370 | 183 | 30 |
| JWH-175 | 370 | 155; 144; 127; 200 | 35; 40; 40; 30 |
| JWH-018 | 342 | 155; 144; 127; 200; 216 | 35; 40; 40; 30; 30 |
| - Mono-OH | 358 | 155; 127; 144; 200; 237 | 35; 40; 40; 30; 30 |
| - Di-OH | 374 | 155; 144; 127; 200; 230 | 35; 40; 40; 30; 30 |
| - Carboxylate - Dehydrogenate-mono-OH | 372 | 155; 144; 127; 200; 230 | 35; 40; 40; 30; 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirri, M.; Arfè, R.; Bilel, S.; Corli, G.; Marchetti, B.; Fantinati, A.; Vincenzi, F.; De-Giorgio, F.; Camuto, C.; Mazzarino, M.; et al. In Vivo Bio-Activation of JWH-175 to JWH-018: Pharmacodynamic and Pharmacokinetic Studies in Mice. Int. J. Mol. Sci. 2022, 23, 8030. https://doi.org/10.3390/ijms23148030
Tirri M, Arfè R, Bilel S, Corli G, Marchetti B, Fantinati A, Vincenzi F, De-Giorgio F, Camuto C, Mazzarino M, et al. In Vivo Bio-Activation of JWH-175 to JWH-018: Pharmacodynamic and Pharmacokinetic Studies in Mice. International Journal of Molecular Sciences. 2022; 23(14):8030. https://doi.org/10.3390/ijms23148030
Chicago/Turabian StyleTirri, Micaela, Raffaella Arfè, Sabrine Bilel, Giorgia Corli, Beatrice Marchetti, Anna Fantinati, Fabrizio Vincenzi, Fabio De-Giorgio, Cristian Camuto, Monica Mazzarino, and et al. 2022. "In Vivo Bio-Activation of JWH-175 to JWH-018: Pharmacodynamic and Pharmacokinetic Studies in Mice" International Journal of Molecular Sciences 23, no. 14: 8030. https://doi.org/10.3390/ijms23148030