
Genetic and Biochemical Aspects of Floral Scents in Roses


Abstract
:1. Introduction
2. Scent Composition of Modern Roses
3. Molecular Research Progress on Rose Scent Biosynthesis
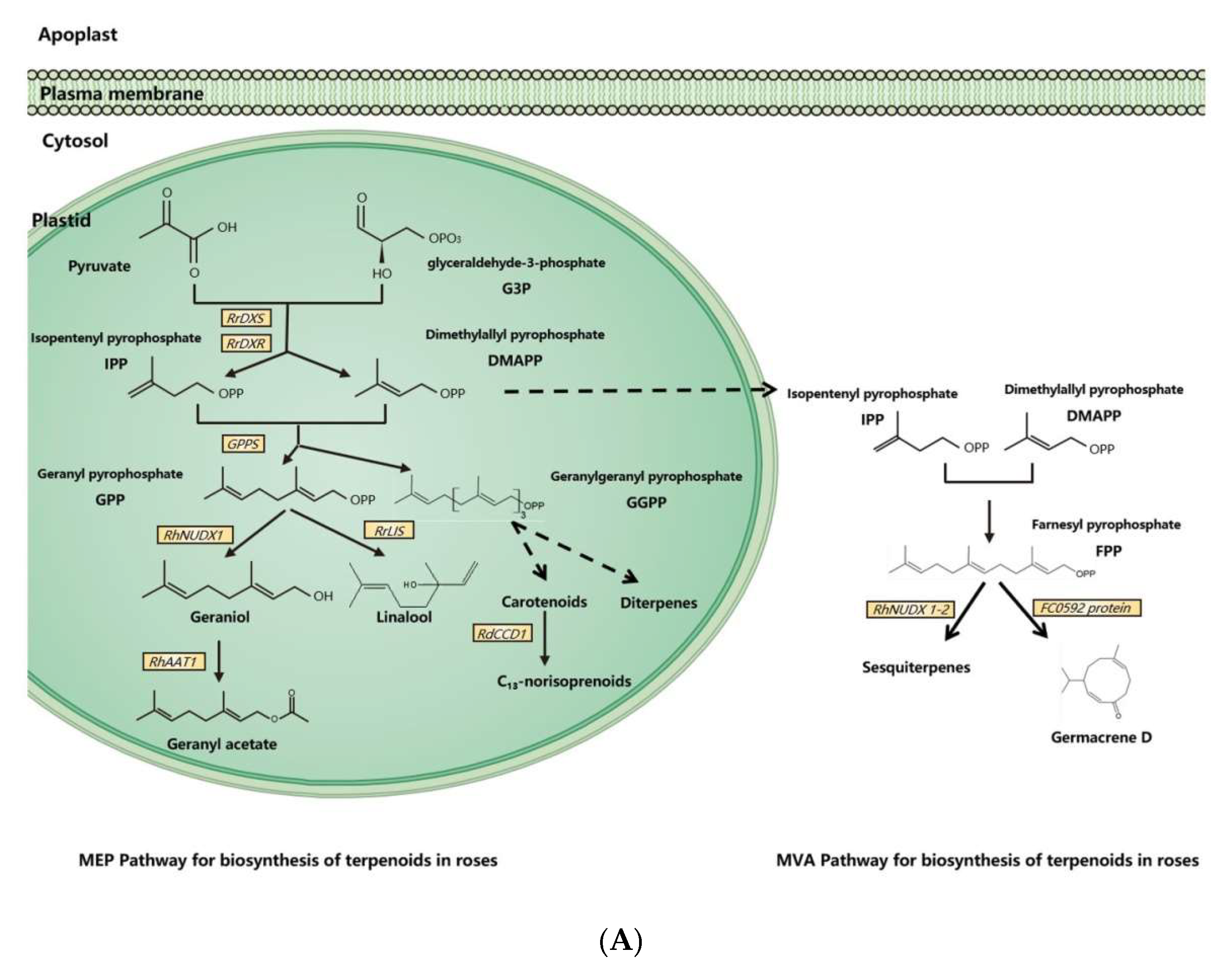
3.1. Biosynthesis of Terpenoids in Rose Floral Scents
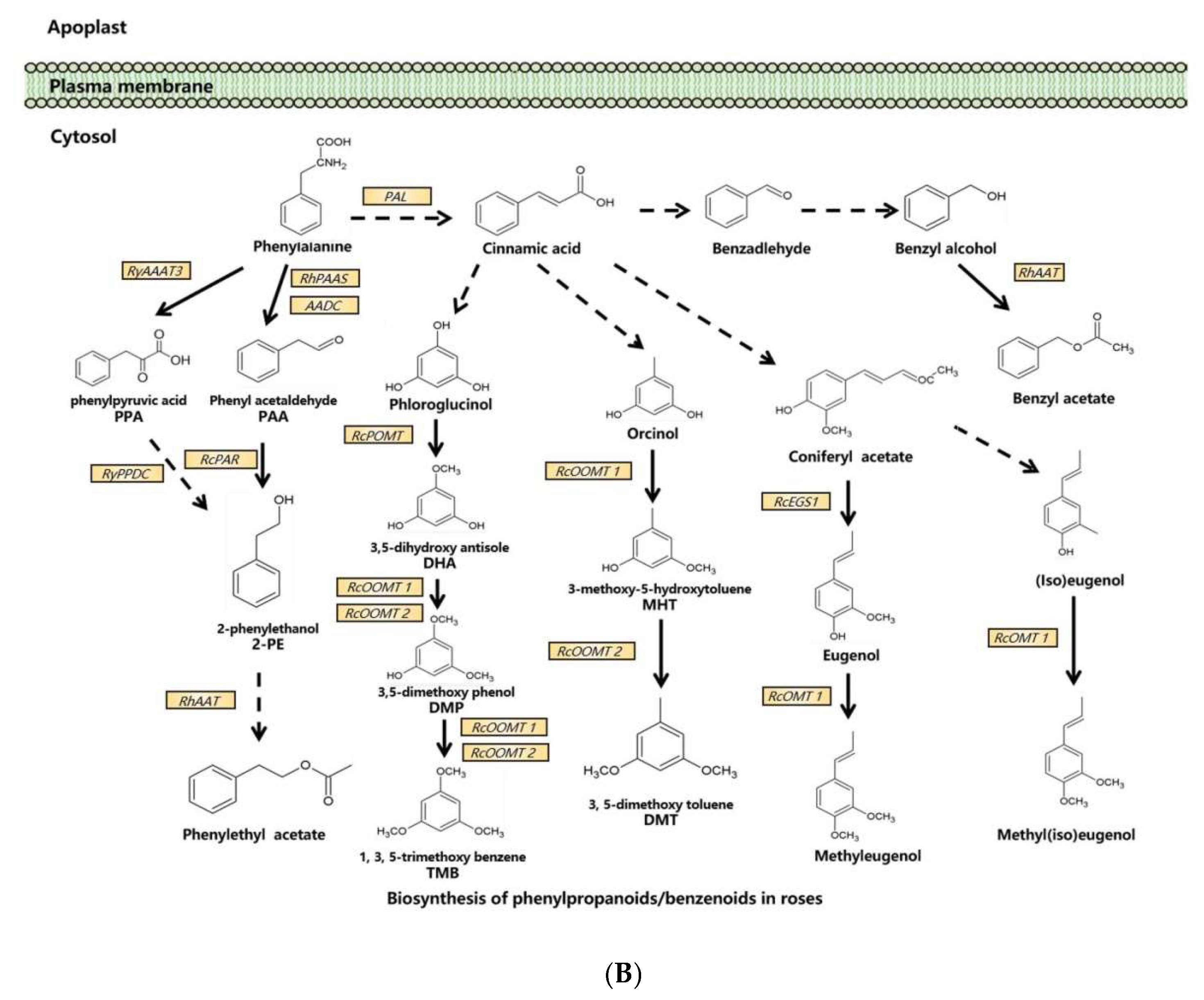
3.2. Biosynthesis of Phenylpropanoids/Benzenoids in Rose Floral Scents
3.3. Transcriptional Regulation of Rose Floral Scent Synthesis
4. Regulation of Floral Scent Production and Emission in Roses
4.1. Floral Scent Production Sites in Roses
4.2. Flower Developmental Stages for Scent Production in Roses
4.3. Diurnal Regulation of Scent Production in Rose Flowers
4.4. Environmental and Internal Factors Affecting Rose Scent Production
4.5. Internal Mechanisms of Rose Scent Emission after Production
5. Molecular Breeding and Metabolic Engineering of Rose Floral Scents
6. Obstacles in Further Study for Rose Floral Scents
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [Green Version]
- Caruso, C.M.; Parachnowitsch, A.L. Do Plsants Eavesdrop on Floral Scent Signals? Trends Plant Sci. 2016, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant Volatile Organic Compounds Evolution: Transcriptional Regulation, Epigenetics and Polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef] [PubMed]
- Grammer, K.; Fink, B.; Møller, A.P.; Thornhill, R. Darwinian aesthetics: Sexual selection and the biology of beauty. Biol. Control 2003, 78, 385–407. [Google Scholar] [CrossRef] [PubMed]
- Jianwu, R.; Lina, Y.; Yan, W.; Hongjun, Y. Chemical profile of floral scent at different flower developmental stages of rose de rescht (Rosa damascena Mill.) cultivated in Beijing. J. Essent. Oil Bear. Plants 2016, 19, 433–443. [Google Scholar]
- Croteau, R.; Kutchan, T.; Lewis, N. Natural Products (Secondary Metabolites). In Biochemistry & Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2000; pp. 1251–1318. [Google Scholar]
- Hsieh, M.-H.; Chang, C.-Y.; Hsu, S.-J.; Chen, J.-J. Chloroplast localization of methylerythritol 4-phosphate pathway enzymes and regulation of mitochondrial genes in ispD and ispE albino mutants in Arabidopsis. Plant Mol. Biol. 2008, 66, 663–673. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Gershenzon, J. The chemical diversity of floral scent. In Biology of PLANT volatiles; Jette, T., Knudsen, J.G., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 57–78. [Google Scholar]
- Pulido, P.; Perello, C.; Rodriguez-Concepcion, M. New insights into plant isoprenoid metabolism. Mol. Plant 2012, 5, 964–967. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; Guirimand, G.; Papon, N.; Courdavault, V.; Thabet, I.; Ginis, O.; Bouzid, S.; Giglioli-Guivarc’h, N.; Clastre, M. Peroxisomal localisation of the final steps of the mevalonic acid pathway in planta. Planta 2011, 234, 903. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Boatright, J.; Negre, F.; Chen, X.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N.; et al. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004, 135, 1993–2011. [Google Scholar] [CrossRef] [Green Version]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Schaller, F. Enzymes of the biosynthesis of octadecanoid-derived signalling molecules. J. Exp. Bot. 2001, 52, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Raymond, O.; Gouzy, J.; Just, J.; Badouin, H.; Verdenaud, M.; Lemainque, A.; Vergne, P.; Moja, S.; Choisne, N.; Pont, C. The Rosa genome provides new insights into the domestication of modern roses. Nat. Genet. 2018, 50, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Basim, E.; Basim, H. Antibacterial activity of Rosa damascena essential oil. Fitoterapia 2003, 74, 394–396. [Google Scholar] [CrossRef]
- Elliott, J. Delmar’s Integrated Herb Guide for Nurses. Northeast. Nat. 2002, 9, 360. [Google Scholar]
- Magnard, J.-L.; Roccia, A.; Caissard, J.-C.; Vergne, P.; Sun, P.; Hecquet, R.; Dubois, A.; Hibrand-Saint Oyant, L.; Jullien, F.; Nicolè, F. Biosynthesis of monoterpene scent compounds in roses. Science 2015, 349, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Borda, A.M.; Clark, D.G.; Huber, D.J.; Welt, B.A.; Nell, T.A. Effects of ethylene on volatile emission and fragrance in cut roses: The relationship between fragrance and vase life. Postharvest. Biol. Technol. 2011, 59, 245–252. [Google Scholar] [CrossRef]
- Bendahmane, M.; Dubois, A.; Raymond, O.; Bris, M.L. Genetics and genomics of flower initiation and development in roses. J. Exp. Bot. 2013, 64, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Raymond, O. Domestication et Sélection Dirigée Chez le Rosier: Analyse Historique via les Phénotypes Morphologique, Chimique et Biochimique. Ph.D. Thesis, Université Claude Bernard Lyon 1, Lyon, France, 1999. [Google Scholar]
- De Vries, D.; Dubois, L.A. Rose breeding: Past, present, prospects. In Proceedings of the II International Rose Symposium 424, Antibes, France, 20–24 February 1995. [Google Scholar]
- Reynders-Aloisi, S.; Bollereau, P. Characterisation of genetic diversity in genus Rosa by randomly amplified polymorphic DNA. In Proceedings of the II International Rose Symposium 424, Antibes, France, 20–24 February 1995; ISHS Acta: Antibes, France, 1995. [Google Scholar]
- Flament, I.; Debonneville, C.; Furrer, A. Volatile compounds of roses: Characterization of cultivars based on the headspace analysis of living flower emissions. In ACS Symposium Series (USA); AGRIS: San Francisco, CA, USA, 1993. [Google Scholar]
- Gang, D.R. Evolution of flavors and scents. Annu. Rev. Plant Biol. 2005, 56, 301–325. [Google Scholar] [CrossRef]
- Jirovetz, L.; Buchbauer, G.; Stoyanova, A.; Balinova, A.; Guangjiun, Z.; Xihan, M. Solid phase microextraction/gas chromatographic and olfactory analysis of the scent and fixative properties of the essential oil of Rosa damascena L. from China. Flavour Fragr. J. 2005, 20, 7–12. [Google Scholar] [CrossRef]
- Joichi, A.; Yomogida, K.; Awano, K.i.; Ueda, Y. Volatile components of tea-scented modern roses and ancient Chinese roses. Flavour Fragr. J. 2005, 20, 152–157. [Google Scholar] [CrossRef]
- Scalliet, G.; Journot, N.; Jullien, F.; Baudino, S.; Magnard, J.-L.; Channelière, S.; Vergne, P.; Dumas, C.; Bendahmane, M.; Cock, J.M. Biosynthesis of the major scent components 3,5-dimethoxytoluene and 1,3,5-trimethoxybenzene by novel rose O-methyltransferases. FEBS Lett. 2002, 523, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.S.; Padalia, R.C.; Chauhan, A.; Singh, A.; Yadav, A.K. Volatile constituents of essential oil and rose water of damask rose (Rosa damascena Mill.) cultivars from North Indian hills. Nat. Prod. Res. 2011, 25, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Washio, H.; Straubinger, M.; Knapp, H.; Winterhalter, P. Occurrence of a glucosidic progenitor of rose oxide in rose flowers, Rosa damascena Mill. Nat. Prod. Lett. 1998, 12, 5–10. [Google Scholar] [CrossRef]
- Guterman, I. Rose Scent: Genomics Approach to Discovering Novel Floral Fragrance-Related Genes. Plant Cell 2002, 14, 2325–2338. [Google Scholar] [CrossRef] [Green Version]
- Shalit, M.; Guterman, I.; Volpin, H.; Bar, E.; Tamari, T.; Menda, N.; Adam, Z.; Zamir, D.; Vainstein, A.; Weiss, D. Volatile ester formation in roses. Identification of an acetyl-coenzyme A. Geraniol/Citronellol acetyltransferase in developing rose petals. Plant Physiol. 2003, 131, 1868–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalliet, G.; Lionnet, C.; Le Bechec, M.; Dutron, L.; Magnard, J.-L.; Baudino, S.; Bergougnoux, V.; Jullien, F.; Chambrier, P.; Vergne, P. Role of petal-specific orcinol O-methyltransferases in the evolution of rose scent. Plant Physiol. 2006, 140, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Watanabe, N.; Mita, S.; Ueda, Y.; Shibuya, M.; Ebizuka, Y. Two O-Methyltransferases isolated from flower petals of Rosa chinensis var. spontanea involved in scent biosynthesis. J. Biosci. Bioeng. 2003, 96, 119–128. [Google Scholar] [CrossRef]
- Lavid, N.; Wang, J.; Shalit, M.; Guterman, I.; Bar, E.; Beuerle, T.; Menda, N.; Shafir, S.; Zamir, D.; Adam, Z. O-methyltransferases involved in the biosynthesis of volatile phenolic derivatives in rose petals. Plant Physiol. 2002, 129, 1899–1907. [Google Scholar] [CrossRef] [Green Version]
- Scalliet, G.; Piola, F.; Douady, C.J.; Réty, S.; Raymond, O.; Baudino, S.; Bordji, K.; Bendahmane, M.; Dumas, C.; Cock, J.M. Scent evolution in Chinese roses. Proc. Natl. Acad. Sci. USA 2008, 105, 5927–5932. [Google Scholar]
- Babaei, A.; Tabaei-Aghdaei, S.R.; Khosh-Khui, M.; Omidbaigi, R.; Naghavi, M.R.; Esselink, G.D.; Smulders, M.J. Microsatellite analysis of Damask rose (Rosa damascena Mill.) accessions from various regions in Iran reveals multiple genotypes. BMC Plant Biol. 2007, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baydar, N.G.; Baydar, H.; Debener, T. Analysis of genetic relationships among Rosa damascena plants grown in Turkey by using AFLP and microsatellite markers. J. Biotechnol. 2004, 111, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Kiani, M.; Zamani, Z.; Khalighi, A.; Fatahi, R.; Byrne, D.H. Wide genetic diversity of Rosa damascena Mill. germplasm in Iran as revealed by RAPD analysis. Sci. Hortic. 2008, 115, 386–392. [Google Scholar] [CrossRef]
- Rusanov, K.; Kovacheva, N.; Vosman, B.; Zhang, L.; Rajapakse, S.; Atanassov, A.; Atanassov, I. Microsatellite analysis of Rosa damascena Mill. accessions reveals genetic similarity between genotypes used for rose oil production and old Damask rose varieties. Theor. Appl. Genet. 2005, 111, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Hayashi, K.; Yagi, K.; Asai, T.; MacTavish, H.; Picone, J.; Turnbull, C.; Watanabe, N. Biogenesis of 2-phenylethanol in rose flowers: Incorporation of [2H8]L-phenylalanine into 2-phenylethanol and its beta-D-glucopyranoside during the flower opening of Rosa ’Hoh-Jun’ and Rosa damascena Mill. Biosci. Biotechnol. Biochem. 2002, 66, 943–947. [Google Scholar] [CrossRef]
- Dubois, A.; Carrere, S.; Raymond, O.; Pouvreau, B.; Cottret, L.; Roccia, A.; Onesto, J.-P.; Sakr, S.; Atanassova, R.; Baudino, S. Transcriptome database resource and gene expression atlas for the rose. BMC Genom. 2012, 13, 638. [Google Scholar] [CrossRef] [Green Version]
- Dubois, A.; Raymond, O.; Maene, M.; Baudino, S.; Langlade, N.B.; Boltz, V.; Vergne, P.; Bendahmane, M. Tinkering with the C-function: A molecular frame for the selection of double flowers in cultivated roses. PLoS ONE 2010, 5, e9288. [Google Scholar] [CrossRef]
- Ma, N.; Xue, J.; Li, Y.; Liu, X.; Dai, F.; Jia, W.; Luo, Y.; Gao, J. Rh-PIP2; 1, a rose aquaporin gene, is involved in ethylene-regulated petal expansion. Plant Physiol. 2008, 148, 894–907. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, N.; Fukuchi-Mizutani, M.; Katsumoto, Y.; Togami, J.; Senior, M.; Matsuda, Y.; Furuichi, K.; Yoshimoto, M.; Matsunaga, A.; Ishiguro, K.; et al. Environmental risk assessment and field performance of rose (Rosa × hybrida) genetically modified for delphinidin production. Plant Biotechnol. 2011, 28, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Spiller, M.; Berger, R.G.; Debener, T. Genetic dissection of scent metabolic profiles in diploid rose populations. Theor. Appl. Genet. 2010, 120, 1461–1471. [Google Scholar] [CrossRef]
- Hendel-Rahmanim, K.; Masci, T.; Vainstein, A.; Weiss, D. Diurnal regulation of scent emission in rose flowers. Planta 2007, 226, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C.; Horváth, G.; Molnár, P.; Turcsi, E.; Deli, J.; Schrader, J.; Sandmann, G.; Schmidt, H.; Schwab, W. Substrate promiscuity of RdCCD1, a carotenoid cleavage oxygenase from Rosa damascena. Phytochemistry 2009, 70, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C.; Molnár, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Chen, C.; Li, T.; Wang, M.; Tao, J.; Zhao, D.; Sheng, L. Flowery odor formation revealed by differential expression of monoterpene biosynthetic genes and monoterpene accumulation in rose (Rosa rugosa Thunb.). Plant Physiol. Biochem. 2014, 75, 80–88. [Google Scholar] [CrossRef]
- Guterman, I.; Masci, T.; Chen, X.; Negre, F.; Pichersky, E.; Dudareva, N.; Weiss, D.; Vainstein, A. Generation of phenylpropanoid pathway-derived volatiles in transgenic plants: Rose alcohol acetyltransferase produces phenylethyl acetate and benzyl acetate in petunia flowers. Plant Mol. Biol. 2006, 60, 555–563. [Google Scholar] [CrossRef]
- Farhi, M.; Lavie, O.; Masci, T.; Hendel-Rahmanim, K.; Weiss, D.; Abeliovich, H.; Vainstein, A. Identification of rose phenylacetaldehyde synthase by functional complementation in yeast. Plant Mol. Biol. 2010, 72, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Kobayashi, H.; Sakai, M.; Hirata, H.; Asai, T.; Ohnishi, T.; Baldermann, S.; Watanabe, N. Functional characterization of rose phenylacetaldehyde reductase (PAR), an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol. J. Plant Physiol. 2011, 168, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Ohnishi, T.; Ishida, H.; Tomida, K.; Sakai, M.; Hara, M.; Watanabe, N. Functional characterization of aromatic amino acid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts. J. Plant Physiol. 2012, 169, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Ohnishi, T.; Tomida, K.; Ishida, H.; Kanda, M.; Sakai, M.; Yoshimura, J.; Suzuki, H.; Ishikawa, T.; Dohra, H. Seasonal induction of alternative principal pathway for rose flower scent. Sci. Rep. 2016, 6, 20234. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Watanabe, N.; Mita, S.; Dohra, H.; Ueda, Y.; Shibuya, M.; Ebizuka, Y. The key role of phloroglucinol O-methyltransferase in the biosynthesis of Rosa chinensis volatile 1, 3, 5-trimethoxybenzene. Plant Physiol. 2004, 135, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yan, H.; Zhang, H. Cloning and expression analysis of eugenol synthase gene RcEGS1 in Rosa chinensis Pallida. Acta Hortic. Sin. 2012, 39, 1387–1394. [Google Scholar]
- Sakai, M.; Hirata, H.; Sayama, H.; Sekiguchi, K.; Itano, H.; Asai, T.; Dohra, H.; Hara, M.; Watanabe, N. Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes, a PLP-dependent decarboxylase and a phenylacetaldehyde reductase. Biosci. Biotechnol. Biochem. 2007, 71, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, H.; Wang, Q.; Jian, H.; Qiu, X.; Wang, J.; Tang, K. Isolation and identification of a putative scent-related gene RhMYB1 from rose. Mol. Biol. Rep. 2011, 38, 4475–4482. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Concepción, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids. A metabolic milestone achieved through genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Dégut, C.; Réty, S.; Caissard, J.C.; Hibrand-Saint Oyant, L.; Bony, A.; Paramita, S.N.; Conart, C.; Magnard, J.L.; Jeauffre, J. Functional diversification in the Nudix hydrolase gene family drives sesquiterpene biosynthesis in Rosa× wichurana. Plant J. 2020, 104, 185–199. [Google Scholar] [CrossRef]
- Aharoni, A.; Keizer, L.C.; Bouwmeester, H.J.; Sun, Z.; Alvarez-Huerta, M.; Verhoeven, H.A.; Blaas, J.; van Houwelingen, A.M.; De Vos, R.C.; van der Voet, H. Identification of the SAAT gene involved in strawberry flavor biogenesis by use of DNA microarrays. Plant Cell 2000, 12, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Shalit, M.; Katzir, N.; Tadmor, Y.; Larkov, O.; Burger, Y.; Shalekhet, F.; Lastochkin, E.; Ravid, U.; Amar, O.; Edelstein, M. Acetyl-CoA: Alcohol acetyltransferase activity and aroma formation in ripening melon fruits. J. Agric. Food Chem. 2001, 49, 794–799. [Google Scholar] [CrossRef]
- Colby, S.M.; Crock, J.; Dowdle-Rizzo, B.; Lemaux, P.G.; Croteau, R. Germacrene C synthase from Lycopersicon esculentum cv. VFNT Cherry tomato: cDNA isolation, characterization, and bacterial expression of the multiple product sesquiterpene cyclase. Proc. Natl. Acad. Sci. USA 1998, 95, 2216–2221. [Google Scholar] [CrossRef] [Green Version]
- Sakai, M.; Saori, T.; Hiroshi, H.; Tatsuo, A.; Hideo, D.; Masakazu, H.; Naoharu, W. Purification and Characterization of β-Glucosidase Involved in the Emission of 2-Phenylethanol from Rose Flowers. Biosci. Biotechnol. Biochem. 2008, 72, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Nagegowda, D.A.; Gupta, P. Advances in biosynthesis, regulation, and metabolic engineering of plant specialized terpenoids. Plant Sci. 2020, 294, 110457. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, J.C.; Haring, M.A.; van Tunen, A.J.; Schuurink, R.C. ODORANT1 regulates fragrance biosynthesis in petunia flowers. Plant Cell 2005, 17, 1612–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzer-Rimon, B.; Farhi, M.; Albo, B.; Cna’ani, A.; Zvi, M.M.B.; Masci, T.; Edelbaum, O.; Yu, Y.; Shklarman, E.; Ovadis, M. The R2R3-MYB–like regulatory factor EOBI, acting downstream of EOBII, regulates scent production by activating ODO1 and structural scent-related genes in petunia. Plant Cell 2012, 24, 5089–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Moerkercke, A.; Haring, M.A.; Schuurink, R.C. The transcription factor EMISSION OF BENZENOIDS II activates the MYB ODORANT1 promoter at a MYB binding site specific for fragrant petunias. Plant J. 2011, 67, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, T.A.; Schwieterman, M.L.; Wedde, A.E.; Schimmel, B.C.; Marciniak, D.M.; Verdonk, J.C.; Kim, J.Y.; Oh, Y.; Gális, I.; Baldwin, I.T. EOBII controls flower opening by functioning as a general transcriptomic switch. Plant Physiol. 2011, 156, 974–984. [Google Scholar] [CrossRef] [Green Version]
- Spitzer-Rimon, B.; Marhevka, E.; Barkai, O.; Marton, I.; Edelbaum, O.; Masci, T.; Prathapani, N.-K.; Shklarman, E.; Ovadis, M.; Vainstein, A. EOBII, a gene encoding a flower-specific regulator of phenylpropanoid volatiles’ biosynthesis in petunia. Plant Cell 2010, 22, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Cna’ani, A.; Spitzer-Rimon, B.; Ravid, J.; Farhi, M.; Masci, T.; Aravena-Calvo, J.; Ovadis, M.; Vainstein, A. Two showy traits, scent emission and pigmentation, are finely coregulated by the MYB transcription factor PH4 in petunia flowers. New Phytol. 2015, 208, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Colquhoun, T.A.; Kim, J.Y.; Wedde, A.E.; Levin, L.A.; Schmitt, K.C.; Schuurink, R.C.; Clark, D.G. PhMYB4 fine-tunes the floral volatile signature of Petunia× hybrida through PhC4H. J. Exp. Bot. 2010, 62, 1133–1143. [Google Scholar] [CrossRef]
- Xu, Y.-H.; Wang, J.-W.; Wang, S.; Wang, J.-Y.; Chen, X.-Y. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-δ-cadinene synthase-A. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.C.; Hung, Y.C.; Tsai, W.C.; Chen, W.H.; Chen, H.H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zvi, M.M.B.; Shklarman, E.; Masci, T.; Kalev, H.; Debener, T.; Shafir, S.; Ovadis, M.; Vainstein, A. PAP1 transcription factor enhances production of phenylpropanoid and terpenoid scent compounds in rose flowers. New Phytol. 2012, 195, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zvi, M.M.B.; Negre-Zakharov, F.; Masci, T.; Ovadis, M.; Shklarman, E.; Ben-Meir, H.; Tzfira, T.; Dudareva, N.; Vainstein, A. Interlinking showy traits: Co-engineering of scent and colour biosynthesis in flowers. Plant Biotechnol. J. 2008, 6, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P. Molecular and biochemical studies of fragrance biosynthesis in rose. Ph.D. Thesis, Universiteit van Amsterdam, Amsterdam, The Netherlands, 2017. [Google Scholar]
- Baudino, S.; Caissard, J.-C.; Bergougnoux, V.; Jullien, F.; Magnard, J.-L.; Scalliet, G.; Cock, J.M.; Hugueney, P. Production and emission of volatile compounds by petal cells. Plant Signal. Behav. 2007, 2, 525–526. [Google Scholar] [CrossRef] [Green Version]
- Bergougnoux, V.; Caissard, J.-C.; Jullien, F.; Magnard, J.-L.; Scalliet, G.; Cock, J.M.; Hugueney, P.; Baudino, S. Both the adaxial and abaxial epidermal layers of the rose petal emit volatile scent compounds. Planta 2007, 226, 853–866. [Google Scholar] [CrossRef]
- Comba, L.; Corbet, S.; Hunt, H.; Outram, S.; Parker, J.; Glover, B. The role of genes influencing the corolla in pollination of Antirrhinum majus. Plant Cell Environ. 2000, 23, 639–647. [Google Scholar] [CrossRef]
- Ben-Nissan, G.; Weiss, D. The petunia homologue of tomato gast1: Transcript accumulation coincides with gibberellin-induced corolla cell elongation. Plant Mol. Biol. 1996, 32, 1067–1074. [Google Scholar] [CrossRef]
- Martin, C.; Gerats, T. Control of pigment biosynthesis genes during petal development. Plant Cell 1993, 5, 1253. [Google Scholar] [CrossRef]
- Shalit, M.; Shafir, S.; Larkov, O.; Bar, E.; Kaslassi, D.; Adam, Z.; Zamir, D.; Vainstein, A.; Weiss, D.; Ravid, U. Volatile compounds emitted by rose cultivars: Fragrance perception by man and honeybees. Isr. J. Plant Sci. 2004, 52, 245–255. [Google Scholar] [CrossRef]
- Karami, A.; Niazi, A.; Kavoosi, G.; Khosh-Khui, M.; Salehi, H. Temporal characterization of 2-phenylethanol in strongly and weakly scented genotypes of damask rose. Physiol. Mol. Biol. Plants 2015, 21, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Baldermann, S.; Cao, S.; Lu, Y.; Liu, C.; Hirata, H.; Watanabe, N. Developmental patterns of emission of scent compounds and related gene expression in roses of the cultivar Rosa x hybrida cv. ‘Yves Piaget’. Plant Physiol. Biochem. 2015, 87, 109–114. [Google Scholar] [CrossRef]
- Raguso, R.A.; Levin, R.A.; Foose, S.E.; Holmberg, M.W.; McDade, L.A. Fragrance chemistry, nocturnal rhythms and pollination “syndromes” in Nicotiana. Phytochemistry 2003, 63, 265–284. [Google Scholar] [CrossRef]
- Effmert, U.; Große, J.; Röse, U.S.; Ehrig, F.; Kägi, R.; Piechulla, B. Volatile composition, emission pattern, and localization of floral scent emission in Mirabilis jalapa (Nyctaginaceae). Am. J. Bot. 2005, 92, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Hoballah, M.E.; Stuurman, J.; Turlings, T.C.; Guerin, P.M.; Connetable, S.; Kuhlemeier, C. The composition and timing of flower odour emission by wild Petunia axillaris coincide with the antennal perception and nocturnal activity of the pollinator Manduca sexta. Planta 2005, 222, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Saona, C.; Parra, L.; Quiroz, A.; Isaacs, R. Variation in highbush blueberry floral volatile profiles as a function of pollination status, cultivar, time of day and flower part: Implications for flower visitation by bees. Ann. Bot. 2011, 107, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Helsper, J.P.; Davies, J.A.; Bouwmeester, H.J.; Krol, A.F.; van Kampen, M.H. Circadian rhythmicity in emission of volatile compounds by flowers of Rosa hybrida L. cv. Honesty. Planta 1998, 207, 88–95. [Google Scholar] [CrossRef]
- Picone, J.M.; Clery, R.A.; Watanabe, N.; MacTavish, H.S.; Turnbull, C.G. Rhythmic emission of floral volatiles from Rosa damascena semperflorens cv. ‘Quatre Saisons’. Planta 2004, 219, 468–478. [Google Scholar]
- Zu, P.; Blanckenhorn, W.U.; Schiestl, F.P. Heritability of floral volatiles and pleiotropic responses to artificial selection in Brassica rapa. New Phytol. 2016, 209, 1208–1219. [Google Scholar] [CrossRef]
- Sagae, M.; Oyama-Okubo, N.; Ando, T.; Marchesi, E.; Nakayama, M. Effect of temperature on the floral scent emission and endogenous volatile profile of Petunia axillaris. Biosci. Biotechnol. Biochem. 2008, 72, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Ravid, J.; Spitzer-Rimon, B.; Takebayashi, Y.; Seo, M.; Cna’ani, A.; Aravena-Calvo, J.; Masci, T.; Farhi, M.; Vainstein, A. GA as a regulatory link between the showy floral traits color and scent. New Phytol. 2017, 215, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Negre, F.; Kish, C.M.; Boatright, J.; Underwood, B.; Shibuya, K.; Wagner, C.; Clark, D.G.; Dudareva, N. Regulation of methylbenzoate emission after pollination in snapdragon and petunia flowers. Plant Cell 2003, 15, 2992–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, B.A.; Tieman, D.M.; Shibuya, K.; Dexter, R.J.; Loucas, H.M.; Simkin, A.J.; Sims, C.A.; Schmelz, E.A.; Klee, H.J.; Clark, D.G. Ethylene-regulated floral volatile synthesis in petunia corollas. Plant Physiol. 2005, 138, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sexton, R.; Stopford, A.P.; Moodie, W.T.; Porter, A.E. Aroma production from cut sweet pea flowers (Lathyrus odoratus): The role of ethylene. Physiol. Plant. 2005, 124, 381–389. [Google Scholar] [CrossRef]
- Kondo, M.; Oyama-Okubo, N.; Ando, T.; Marchesi, E.; Nakayama, M. Floral scent diversity is differently expressed in emitted and endogenous components in Petunia axillaris lines. Ann. Bot. 2006, 98, 1253–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyama-Okubo, N.; Ando, T.; Watanabe, N.; Marchesi, E.; Uchida, K.; Nakayama, M. Emission mechanism of floral scent in Petunia axillaris. Biosci. Biotechnol. Biochem. 2005, 69, 773–777. [Google Scholar] [CrossRef] [Green Version]
- Loughrin, J.H.; Hamilton-Kemp, T.R.; Burton, H.R.; Andersen, R.A.; Hildebrand, D.F. Glycosidically bound volatile components of Nicotiana sylvestris and N suaveolens flowers. Phytochemistry 1992, 31, 1537–1540. [Google Scholar] [CrossRef]
- Ogawa, K.; Ijima, Y.; Guo, W.; Watanabe, N.; Usui, T.; Dong, S.; Tong, Q.; Sakata, K. Purification of a β-primeverosidase concerned with alcoholic aroma formation in tea leaves (cv. Shuixian) to be processed to oolong tea. J. Agric. Food Chem. 1997, 45, 877–882. [Google Scholar]
- Reuveni, M.; Sagi, Z.; Evnor, D.; Hetzroni, A. β-Glucosidase activity is involved in scent production in Narcissus flowers. Plant Sci. 1999, 147, 19–24. [Google Scholar] [CrossRef]
- Schulz, G.; Stahl-Biskup, E. Essential oils and glycosidic bound volatiles from leaves, stems, flowers and roots of Hyssopus officinalis L. (Lamiaceae). Flavour Fragrance J. 1991, 6, 69–73. [Google Scholar] [CrossRef]
- Hayashi, S.; Yagi, K.; Ishikawa, T.; Kawasaki, M.; Asai, T.; Picone, J.; Turnbull, C.; Hiratake, J.; Sakata, K.; Takada, M. Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2 H 8] L-phenylalanine in rose flowers. Tetrahedron 2004, 60, 7005–7013. [Google Scholar] [CrossRef]
- Jetter, R. Examination of the processes involved in the emission of scent volatiles from flowers. In Biology of Floral Scent; Natalia, D., Eran, P., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 125–144. [Google Scholar]
- Adebesin, F.; Widhalm, J.R.; Boachon, B.; Lefèvre, F.; Pierman, B.; Lynch, J.H.; Alam, I.; Junqueira, B.; Benke, R.; Ray, S. Emission of volatile organic compounds from petunia flowers is facilitated by an ABC transporter. Science 2017, 356, 1386–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Zu, P.; Schiestl, F.P. The molecular bases of floral scent evolution under artificial selection: Insights from a transcriptome analysis in Brassica rapa. Sci. Rep. 2016, 6, 36966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreier, K.; Spiller, M.; Linde, M.; Kaufmann, H.; Berger, R.; Krings, U.; Debener, T.; Bretzke, M. Genetic and molecular analyses of key loci involved in self incompatibility and floral scent in roses. In Proceedings of the V International Symposium on Rose Research and Cultivation 870, Gifu, Japan, 24–29 May 2009; ISHS Acta: Gifu, Japan, 2009. [Google Scholar]
- Crespel, L.; Chirollet, M.; Durel, C.; Zhang, D.; Meynet, J.; Gudin, S. Mapping of qualitative and quantitative phenotypic traits in Rosa using AFLP markers. Theor. Appl. Genet. 2002, 105, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Linde, M.; Hattendorf, A.; Kaufmann, H.; Debener, T. Powdery mildew resistance in roses: QTL mapping in different environments using selective genotyping. Theor. Appl. Genet. 2006, 113, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Spiller, M.; Linde, M.; Hibrand-Saint Oyant, L.; Tsai, C.-J.; Byrne, D.H.; Smulders, M.J.; Foucher, F.; Debener, T. Towards a unified genetic map for diploid roses. Theor. Appl. Genet. 2011, 122, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Denneboom, C.; Hattendorf, A.; Dolstra, O.; Debener, T.; Stam, P.; Visser, P. Construction of an integrated map of rose with AFLP, SSR, PK, RGA, RFLP, SCAR and morphological markers. Theor. Appl. Genet. 2005, 110, 766–777. [Google Scholar] [CrossRef]
- Yan, Z.; Visser, P.; Hendriks, T.; Prins, T.; Stam, P.; Dolstra, O. QTL analysis of variation for vigour in rose. Euphytica 2007, 154, 53–62. [Google Scholar] [CrossRef]
- Cherri-Martin, M.; Jullien, F.; Heizmann, P.; Baudino, S. Fragrance heritability in Hybrid Tea roses. Sci. Hortic. 2007, 113, 177–181. [Google Scholar] [CrossRef]
- Nowak, R. Chemical Composition of Hips Essential Oils of Some Rosa L. Species December 13, 2004. Z. Für Nat. C 2005, 60, 369–378. [Google Scholar] [CrossRef]
- Fridman, E.; Pichersky, E. Metabolomics, genomics, proteomics, and the identification of enzymes and their substrates and products. Curr. Opin. Plant Biol. 2005, 8, 242–248. [Google Scholar] [CrossRef]
- Spitzer, B.; Zvi, M.M.; Ovadis, M.; Marhevka, E.; Barkai, O.; Edelbaum, O.; Marton, I.; Masci, T.; Alon, M.; Morin, S.; et al. Reverse genetics of floral scent: Application of tobacco rattle virus-based gene silencing in Petunia. Plant Physiol. 2007, 145, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Shi, S.; Ma, N.; Cao, X.; Zhang, H.; Qiu, X.; Wang, Q.; Jian, H.; Zhou, N.; Zhang, Z. Graft-accelerated virus-induced gene silencing facilitates functional genomics in rose flowers. J. Integr. Plant Biol. 2018, 60, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; Alvarez-Huerta, M.; Neef, E.; Verstappen, F.W.; Bouwmeester, H.J.; Aharoni, A. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant Physiol. 2004, 135, 1865–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, A.; Remay, A.; Raymond, O.; Balzergue, S.; Chauvet, A.; Maene, M.; Pecrix, Y.; Yang, S.H.; Jeauffre, J.; Thouroude, T.; et al. Genomic approach to study floral development genes in Rosa sp. PLoS ONE 2011, 6, e28455. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wan, H.; Cheng, T.; Wang, J.; Yang, W.; Pan, H.; Zhang, Q. Comparative RNA-seq analysis of transcriptome dynamics during petal development in Rosa chinensis. Sci. Rep. 2017, 7, 43382. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Park, J.H.; Lim, C.J.; Lim, J.Y.; Ryu, J.Y.; Lee, B.W.; Choi, J.P.; Kim, W.B.; Lee, H.Y.; Choi, Y.; et al. Small RNA and transcriptome deep sequencing proffers insight into floral gene regulation in Rosa cultivars. BMC Genom. 2012, 13, 657. [Google Scholar] [CrossRef] [Green Version]
- Pei, H.; Ma, N.; Chen, J.; Zheng, Y.; Tian, J.; Li, J.; Zhang, S.; Fei, Z.; Gao, J. Integrative analysis of miRNA and mRNA profiles in response to ethylene in rose petals during flower opening. PLoS ONE 2013, 8, e64290. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, H.; Chen, M.; Jian, H.; Baudino, S.; Caissard, J.C.; Bendahmane, M.; Li, S.; Zhang, T.; Zhou, N.; et al. Transcriptome and gene expression analysis during flower blooming in Rosa chinensis ‘Pallida’. Gene 2014, 540, 96–103. [Google Scholar] [CrossRef]
- Nakamura, N.; Hirakawa, H.; Sato, S.; Otagaki, S.; Matsumoto, S.; Tabata, S.; Tanaka, Y. Genome structure of Rosa multiflora, a wild ancestor of cultivated roses. DNA Res. 2018, 25, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Piola, F.; Chessel, D.; Jay, M.; Heizmann, P. The domestication process of the Modern Rose: Genetic structure and allelic composition of the rose complex. Theor. Appl. Genet. 2001, 102, 398–404. [Google Scholar] [CrossRef]
- Pécrix, Y.; Rallo, G.; Folzer, H.; Cigna, M.; Gudin, S.; Le Bris, M. Polyploidization mechanisms: Temperature environment can induce diploid gamete formation in Rosa sp. J. Exp. Bot. 2011, 62, 3587–3597. [Google Scholar] [CrossRef] [Green Version]
- Gar, O.; Sargent, D.J.; Tsai, C.-J.; Pleban, T.; Shalev, G.; Byrne, D.H.; Zamir, D. An autotetraploid linkage map of rose (Rosa hybrida) validated using the strawberry (Fragaria vesca) genome sequence. PLoS ONE 2011, 6, e20463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajapakse, S.; Byrne, D.; Zhang, L.; Anderson, N.; Arumuganathan, K.; Ballard, R. Two genetic linkage maps of tetraploid roses. Theor. Appl. Genet. 2001, 103, 575–583. [Google Scholar] [CrossRef]
- Yu, C.; Luo, L.; Pan, H.; Guo, X.; Wan, H.; Zhang, Q. Filling gaps with construction of a genetic linkage map in tetraploid roses. Front. Plant Sci. 2014, 5, 796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Byrne, D.; Ballard, R.; Rajapakse, S. Microsatellite marker development in rose and its application in tetraploid mapping. J. Am. Soc. Hort. Sci. 2006, 131, 380–387. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Compound Variety | Compounds | Odor | References |
|---|---|---|---|
| terpenes | β-cubebene | citrus, fruity, radish | [25,31] |
| β-elemene | herbal, waxy, fresh | [26,31] | |
| δ-cadinene | thyme, herbal, woody | [27] | |
| germacrene D | woody, spice | [31] | |
| geraniol | rose-like, sweet | [28,31,32] | |
| citronellol | fresh rosy | [27,28,31] | |
| nerol | lemon-like, floral | [27,28,32] | |
| linalool | citrus and floral | [27,32] | |
| farnesyl acetate | green-floral rose | [27] | |
| geranyl acetate | lavender | [27,31] | |
| citronellyl acetate | fresh, rose, fruity odor | [25,31] | |
| neryl acetate | rose and lavender-like | [25,31] | |
| citral | citrus and lemon | [25,27] | |
| dihydro-β-ionone | violet-like and earthy | [27] | |
| rose oxide | herbal, green floral, earthy | [30] | |
| Phenylpropanoids/ benzenoids | 2-phenylethanol | honey-like | [28,30,31] |
| 2-phenylethyl acetate | sweet, honey, rosy, with a slight yeasty honey note | [27,30,31,32] | |
| 1,3,5-Trimethoxybenzene (TMB) | phenolic spicy, earthy note | [28,33,34] | |
| dimethoxytoluene (DMT) | fresh, earthy, phenolic spicy | [28,33] | |
| benzyl acetate | floral, fruity, sweet, fresh | [27,32] | |
| eugenol | clove, carnation | [28,35] | |
| methyl eugenol | clove, carnation | [27,34] | |
| methyl isoeugenol | clove, carnation, woody | [27,34] | |
| fatty-acid derivatives | cis-3-hexenyl-1-alcohol | fresh and leafy green | [25,28,32] |
| 2-hexenyl acetate | fresh, fruity green | [27,31,32] | |
| cis-3-hexenyl acetate | fresh and leafy green | [27,31] |
| Pathway | Gene | Species | References |
|---|---|---|---|
| Terpenoids | RhGDS | R. hybrid ‘Fragrant Cloud’ | [47] |
| RhCCD1 | R. damascene | [48] | |
| RhCCD4 | R. damascene | [49] | |
| RrLIS | R. rugosa Thunb. ‘Tangzi’ | [50] | |
| RhAAT | R. hybrida ‘Fragrant Cloud’ | [32,51] | |
| RrAAT | R. rugosa Thunb. ‘Tangzi’ | [50] | |
| RrDXS | R. rugosa Thunb. ‘Tangzi’ | [50] | |
| RrDXR | R. rugosa Thunb. ‘Tangzi’ | [50] | |
| RcGDS | R. chinensis ‘Old Blush’ | [42] | |
| RhNUDX1 | R. chinensis ‘Old Blush’ | [18] | |
| Phenylpropanoids /benzenoids | RhPAAS | R. hybrida ‘Fragrant Cloud’ | [52] |
| Rose-PAR | R.×damascena Mill | [53] | |
| RyAAAT3 | R. hybrida ‘Yves Piaget’ | [54] | |
| RyPPDC | R. hybrida ‘Yves Piaget’ | [55] | |
| RcPOMT | R. chinensis Jacq.var. spontanea | [56] | |
| RcEGS1 | R. chinensis ‘Old Blush’ | [57] | |
| RcOOMT1 | R. hybrida ‘Fragrant Cloud’ and ‘Golden Gate’; R. chinensis Jacq.var. spontanea | [34,35] | |
| RcOOMT2 | R. hybrida ‘Fragrant Cloud’ and ‘Golden Gate’; R. chinensis Jacq.var. spontanea | [34,35] | |
| RcOMT 1 | R. chinensis var. spontanea | [34] | |
| RcOMT 2 | R. chinensis var. spontanea | [34] | |
| AADC | R. ‘Hoh-Jun’ | [58] | |
| RhMYB1 | R. hybrida ‘Jinyindao’ | [59] |
| Pathways | Scent-Related Genes or Traits | LG | Population | Rose Species | References |
|---|---|---|---|---|---|
| terpenoids | geraniol (QTL) | LG1 | Linkage groups 94/1 | R. multiflora | [114] |
| TPS-L (Terpene synthase-like) | LG1 | Linkage groups 94/1 | R. multiflora | [46] | |
| RhCCD1 | LG1 | Linkage groups 94/1 | R. multiflora | [46,114] | |
| RhAAT1 | LG2 | Linkage groups 94/1 + Linkage groups 97/7 | R. multiflora | [46,114] | |
| geranyl acetate | LG2 | Linkage groups 94/1 | R. multiflora | [114] | |
| nerol | LG3 | Linkage groups 94/1 | R. multiflora | [114] | |
| β-citronellol (QTL) | LG3 | Linkage groups 94/1 | R. multiflora | [114] | |
| neryl acetate | LG4 | Linkage groups 94/1 | R. multiflora | [114] | |
| TPS-L (Terpene synthase-like, Farnesyltransferase) | LG4 | Linkage groups 94/1 | R. multiflora | [46] | |
| GDS | LG5 | Linkage groups 94/1 + Linkage groups 97/7 | R. multiflora | [46,114] | |
| TPS-L (Terpene synthase-like) | LG5 | Linkage groups 94/1 | R. multiflora | [46] | |
| alcohol acetate | LG7 | Linkage groups 94/1 | R. multiflora | [114] | |
| Phenylpropanoids | RhPAR | LG1 | Linkage groups 94/1 | R. multiflora | [46,114] |
| RhOOMT1 | LG2 | Linkage groups 94/1 | R. multiflora | [114] | |
| RhOOMT2 | LG2 | Linkage groups 94/1 | R. multiflora | [46] | |
| RcOMT3-1 | LG2 | Linkage groups 94/1 + Linkage groups 97/7 | R. multiflora | [114] | |
| RcOMT3-265 | LG2 | Linkage groups 94/1 | R. multiflora | [46] | |
| BEAT-L | LG2 | Linkage groups 94/1 | R. multiflora | [46] | |
| NMT-L (N-methyltransferase) | LG2 | Linkage groups 97/7 | R. multiflora | [46] | |
| BEAT-L | LG4 | Linkage groups 97/7 | R. multiflora | [46] | |
| RcOMT3-2 | LG4 | Linkage groups 94/1 + Linkage groups 97/7 | R. multiflora | [114] | |
| RcOMT3-280 | LG4 | Linkage groups 97/7 | R. multiflora | [46] | |
| RcOMT1 | LG4 | Linkage groups 97/7 | R. multiflora | [46,114] | |
| phenylethanol (QTL) | LG5 | Linkage groups 94/1 | R. multiflora | [114] | |
| RhAADC | LG5 | Linkage groups 97/7 | R. multiflora | [114] | |
| POMT | LG6 | Linkage groups 94/1 + Linkage groups 97/7 | R. multiflora | [46,114] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, S.; Zhang, Z. Genetic and Biochemical Aspects of Floral Scents in Roses. Int. J. Mol. Sci. 2022, 23, 8014. https://doi.org/10.3390/ijms23148014
Shi S, Zhang Z. Genetic and Biochemical Aspects of Floral Scents in Roses. International Journal of Molecular Sciences. 2022; 23(14):8014. https://doi.org/10.3390/ijms23148014
Chicago/Turabian StyleShi, Shaochuan, and Zhao Zhang. 2022. "Genetic and Biochemical Aspects of Floral Scents in Roses" International Journal of Molecular Sciences 23, no. 14: 8014. https://doi.org/10.3390/ijms23148014