
3′5-Dimaleamylbenzoic Acid Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 3′5-DMBA Attenuates BLM-Induced Pulmonary Fibrosis in Mice
2.2. 3′5-DMBA Reduces Pulmonary Fibrosis-Related Protein Expression in Mice
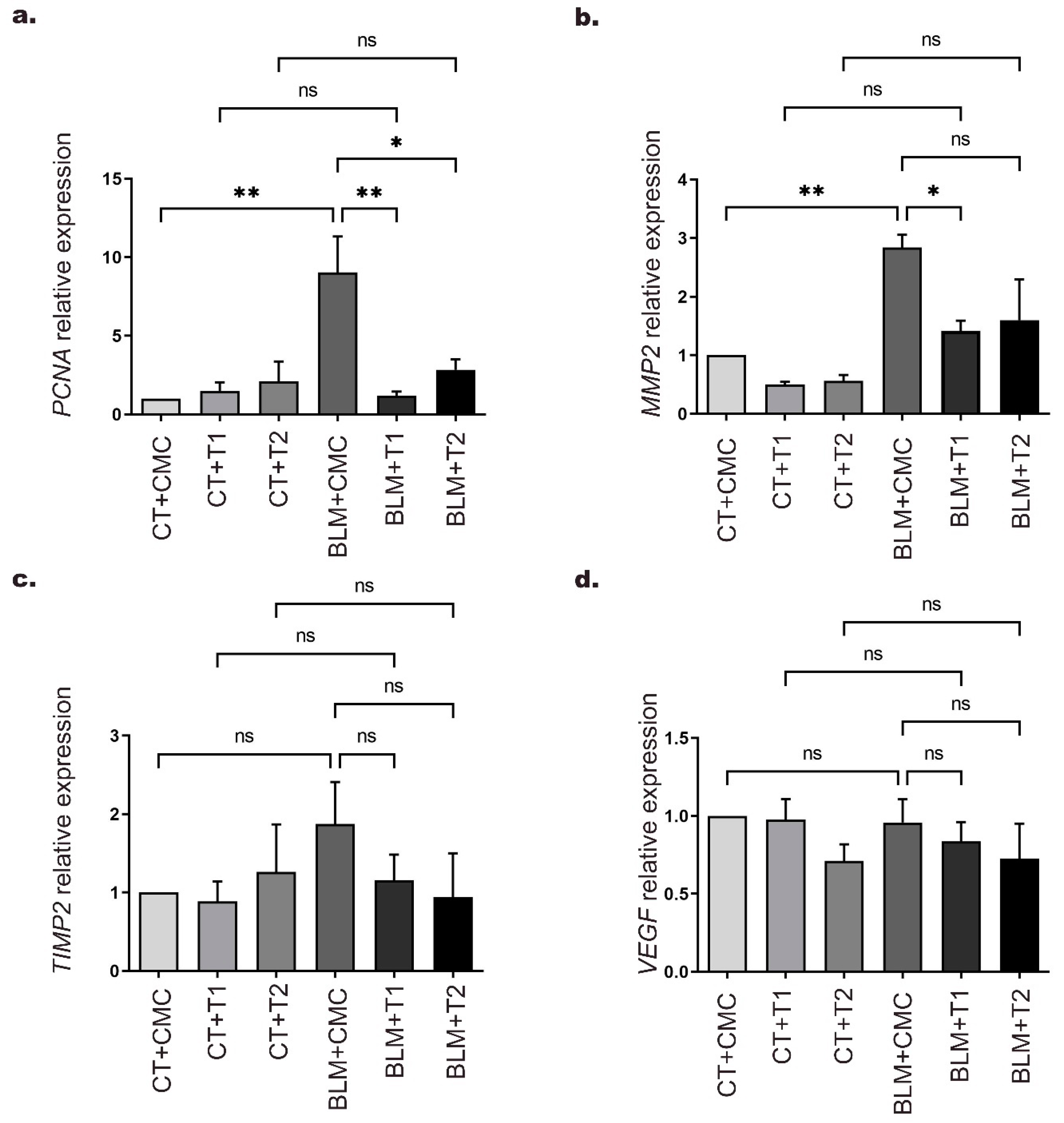
2.3. 3′5-DMBA Modulates Gene Expression Related to BLM-Induced Pulmonary Fibrosis in Mice
2.4. 3′5-DMBA Modulates the Expression of Genes Related to Cell Proliferation and Remodeling of the ECM during Its Antifibrotic Effect
2.5. 3′5-DMBA Exerts Pro-Oxidant Effects without Promoting Increased Lipid Peroxidation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Tissue Preparation
4.4. Histological Analysis
4.5. Immunohistochemical Analysis
4.6. RT-qPCR
4.7. Total, Reduced (GSH), and Oxidized (GSSG) Glutathione Quantification
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lass, D.S.; Grossfeld, D.; Renna, H.A.; Agarwala, P.; Spiegler, P.; Kasselman, L.J.; Glass, A.D.; DeLeon, J.; Reiss, A.B. Idiopathic pulmonary fibrosis: Molecular mechanisms and potential treatment approaches. Respir. Investig. 2020, 58, 320–335. [Google Scholar] [CrossRef]
- Lederer, D.J.; Martinez, F.J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2018, 378, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Du Bois, R.M. An earlier and more confident diagnosis of idiopathic pulmonary fibrosis. Eur. Respir. Rev. 2012, 21, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Sgalla, G.; Iovene, B.; Calvello, M.; Ori, M.; Varone, F.; Richeldi, L. Idiopathic pulmonary fibrosis: Pathogenesis and management. Respir. Res. 2018, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Sauleda, J.; Núñez, B.; Sala, E.; Soriano, J.B. Idiopathic Pulmonary Fibrosis: Epidemiology, Natural History, Phenotypes. Med. Sci. 2018, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinde, A.V.; Humeres, C.; Frangogiannis, N.G. The role of α-smooth muscle actin in fibroblast-mediated matrix contraction and remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Willis, B.C.; Borok, Z. TGF-beta-induced EMT: Mechanisms and implications for fibrotic lung disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L525–L534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergeron, A.; Soler, P.; Kambouchner, M.; Loiseau, P.; Milleron, B.; Valeyre, D.; Hance, A.J.; Tazi, A. Cytokine profiles in idiopathic pulmonary fibrosis suggest an important role for TGF-beta and IL-10. Eur. Respir. J. 2003, 22, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Jobling, M.F.; Mott, J.D.; Finnegan, M.T.; Jurukovski, V.; Erickson, A.C.; Walian, P.J.; Taylor, S.E.; Ledbetter, S.; Lawrence, C.M.; Rifkin, D.B.; et al. Isoform-specific activation of latent transforming growth factor beta (LTGF-beta) by reactive oxygen species. Radiat. Res. 2006, 166, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Bocchino, M.; Agnese, S.; Fagone, E.; Svegliati, S.; Grieco, D.; Vancheri, C.; Gabrielli, A.; Sanduzzi, A.; Avvedimento, E.V. Reactive oxygen species are required for maintenance and differentiation of primary lung fibroblasts in idiopathic pulmonary fibrosis. PLoS ONE 2010, 5, e14003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazaki, K.; Matsuno, Y.; Yoshida, K.; Sherpa, M.; Nakajima, M.; Matsuyama, M.; Kiwamoto, T.; Morishima, Y.; Ishii, Y.; Hizawa, N. ROS-Nrf2 pathway mediates the development of TGF-β1-induced epithelial-mesenchymal transition through the activation of Notch signaling. Eur. J. Cell Biol. 2021, 100, 151181. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Jiménez, E.; Ramírez-Hernández, A.A.; Santos-Álvarez, J.C.; Velázquez-Enríquez, J.M.; Pina-Canseco, S.; Baltiérrez-Hoyos, R.; Vásquez-Garzón, V.R. Involvement of 4-hydroxy-2-nonenal in the pathogenesis of pulmonary fibrosis. Mol. Cell. Biochem. 2021, 476, 4405–4419. [Google Scholar] [CrossRef] [PubMed]
- Tsubouchi, K.; Araya, J.; Yoshida, M.; Sakamoto, T.; Koumura, T.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; et al. Involvement of GPx4-Regulated Lipid Peroxidation in Idiopathic Pulmonary Fibrosis Pathogenesis. J. Immunol. 2019, 203, 2076–2087. [Google Scholar] [CrossRef]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signaling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [Green Version]
- Asensi, M.; Sastre, J.; Pallardo, F.V.; Lloret, A.; Lehner, M.; Garcia-de-la Asuncion, J.; Viña, J. Ratio of reduced to oxidized glutathione as indicator of oxidative stress status and DNA damage. Methods Enzymol. 1999, 299, 267–276. [Google Scholar] [CrossRef]
- Miura, Y.; Lam, M.; Bourke, J.E.; Kanazawa, S. Bimodal fibrosis in a novel mouse model of bleomycin-induced usual interstitial pneumonia. Life Sci. Alliance 2022, 5. [Google Scholar] [CrossRef]
- Liu, T.; De Los Santos, F.G.; Phan, S.H. The Bleomycin Model of Pulmonary Fibrosis. Methods Mol. Biol. 2017, 1627, 27–42. [Google Scholar] [CrossRef]
- Mouratis, M.A.; Aidinis, V. Modeling pulmonary fibrosis with bleomycin. Curr. Opin. Pulm. Med. 2011, 17, 355–361. [Google Scholar] [CrossRef]
- Ravanetti, F.; Ferrini, E.; Ragionieri, L.; Khalajzeyqami, Z.; Nicastro, M.; Ridwan, Y.; Kleinjan, A.; Villetti, G.; Grandi, A.; Stellari, F.F. SSC-ILD mouse model induced by osmotic minipump delivered bleomycin: Effect of Nintedanib. Sci. Rep. 2021, 11, 18513. [Google Scholar] [CrossRef]
- Ravanetti, F.; Ragionieri, L.; Ciccimarra, R.; Ruscitti, F.; Pompilio, D.; Gazza, F.; Villetti, G.; Cacchioli, A.; Stellari, F.F. Modeling pulmonary fibrosis through bleomycin delivered by osmotic minipump: A new histomorphometric method of evaluation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L376–L385. [Google Scholar] [CrossRef] [PubMed]
- Fortier, S.M.; Penke, L.R.; King, D.; Pham, T.X.; Ligresti, G.; Peters-Golden, M. Myofibroblast dedifferentiation proceeds via distinct transcriptomic and phenotypic transitions. JCI Insight 2021, 6, e144799. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Shao, Y.; Zhao, X.; Hong, C.S.; Wang, F.; Lu, X.; Li, J.; Ye, G.; Yan, M.; Zhuang, Z.; et al. Integration of Metabolomics and Transcriptomics Reveals Major Metabolic Pathways and Potential Biomarker Involved in Prostate Cancer. Mol. Cell. Proteom. 2016, 15, 154–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, B.; Lagares, D. Evasion of apoptosis by myofibroblasts: A hallmark of fibrotic diseases. Nat. Rev. Rheumatol. 2020, 16, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Staab-Weijnitz, C.; Fernandez, I.; Schiller, H.; Heinzelmann, K.; Sterclova, M.; Vasakova, M.; Mann, M.; Eickelberg, O. Compartment-specific overexpression of glutathione peroxidases during lung fibrosis. Eur. Respir. J. 2014, 44, 2022. [Google Scholar]
- Huang, P.R.; Yeh, Y.M.; Pao, C.C.; Chen, C.Y.; Wang, T.C. N-(1-Pyrenyl) maleimide inhibits telomerase activity in a cell free system and induces apoptosis in Jurkat cells. Mol. Biol. Rep. 2012, 39, 8899–8905. [Google Scholar] [CrossRef]
- Ali, B.; Kanda Kupa, L.D.; Heluany, C.S.; Drewes, C.C.; Vasconcelos, S.N.S.; Farsky, S.H.P.; Stefani, H.A. Cytotoxic effects of a novel maleimide derivative on epithelial and tumor cells. Bioorg. Chem. 2017, 72, 199–207. [Google Scholar] [CrossRef]
- Andrade-Jorge, E.; Godínez-Victoria, M.; Sánchez-Torres, L.E.; Fabila-Castillo, L.H.; Trujillo-Ferrara, J.G. Aryl Maleimides as Apoptosis Inducers on L5178-Y Murine Leukemia Cells (in silico, in vitro and ex vivo Study). Anticancer Agents Med. Chem. 2016, 16, 1615–1621. [Google Scholar] [CrossRef]
- Carrasco-Torres, G.; Baltiérrez-Hoyos, R.; Andrade-Jorge, E.; Villa-Treviño, S.; Trujillo-Ferrara, J.G.; Vásquez-Garzón, V.R. Cytotoxicity, Oxidative Stress, Cell Cycle Arrest, and Mitochondrial Apoptosis after Combined Treatment of Hepatocarcinoma Cells with Maleic Anhydride Derivatives and Quercetin. Oxid. Med. Cell. Longev. 2017, 2017, 2734976. [Google Scholar] [CrossRef]
- García-Alvarez, J.; Ramirez, R.; Sampieri, C.L.; Nuttall, R.K.; Edwards, D.R.; Selman, M.; Pardo, A. Membrane type-matrix metalloproteinases in idiopathic pulmonary fibrosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2006, 23, 13–21. [Google Scholar]
- Barratt, S.L.; Blythe, T.; Jarrett, C.; Ourradi, K.; Shelley-Fraser, G.; Day, M.J.; Qiu, Y.; Harper, S.; Maher, T.M.; Oltean, S.; et al. Differential Expression of VEGF-A(xxx) Isoforms Is Critical for Development of Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2017, 196, 479–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izbicki, G.; Segel, M.J.; Christensen, T.G.; Conner, M.W.; Breuer, R. Time course of bleomycin-induced lung fibrosis. Int. J. Exp. Pathol. 2002, 83, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, R.G.; Moore, B.B.; Chambers, R.C.; Eickelberg, O.; Königshoff, M.; Kolb, M.; Laurent, G.J.; Nanthakumar, C.B.; Olman, M.A.; Pardo, A.; et al. An Official American Thoracic Society Workshop Report: Use of Animal Models for the Preclinical Assessment of Potential Therapies for Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 2017, 56, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo-Ferrara, J.; Santillan, R.; Beltrán, H.I.; Farfán, N.; Höpfl, H. 1H and 13C NMR spectra for a series of arylmaleamic acids, arylmaleimides, arylsuccinamic acids and arylsuccinimides. Magn. Reson. Chem. 1999, 37, 682–686. [Google Scholar] [CrossRef]
- Arora, A.; Bhuria, V.; Hazari, P.P.; Pathak, U.; Mathur, S.; Roy, B.G.; Sandhir, R.; Soni, R.; Dwarakanath, B.S.; Bhatt, A.N. Amifostine Analog, DRDE-30, Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Front. Pharmacol. 2018, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Tong, M.; Ou, B.; Liu, C.; Hu, C.; Yang, Y. Isorhamnetin protects against bleomycin-induced pulmonary fibrosis by inhibiting endoplasmic reticulum stress and epithelial-mesenchymal transition. Int. J. Mol. Med. 2019, 43, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Eyden, B. The myofibroblast: Phenotypic characterization as a prerequisite to understanding its functions in translational medicine. J. Cell. Mol. Med. 2008, 12, 22–37. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Watanabe, K.; Nabeshima, K.; Hamasaki, M.; Iwasaki, H. Prognostic significance of fibroblastic foci in usual interstitial pneumonia and non-specific interstitial pneumonia. Respirology 2013, 18, 278–283. [Google Scholar] [CrossRef]
- Yamashita, M.; Yamauchi, K.; Chiba, R.; Iwama, N.; Date, F.; Shibata, N.; Kumagai, H.; Risteli, J.; Sato, S.; Takahashi, T.; et al. The definition of fibrogenic processes in fibroblastic foci of idiopathic pulmonary fibrosis based on morphometric quantification of extracellular matrices. Hum. Pathol. 2009, 40, 1278–1287. [Google Scholar] [CrossRef]
- Merkt, W.; Zhou, Y.; Han, H.; Lagares, D. Myofibroblast fate plasticity in tissue repair and fibrosis: Deactivation, apoptosis, senescence and reprogramming. Wound Repair Regen. 2021, 29, 678–691. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Togo, S.; Kadoya, K.; Tulafu, M.; Namba, Y.; Iwai, M.; Watanabe, J.; Nagahama, K.; Okabe, T.; Hidayat, M.; et al. Pirfenidone attenuates lung fibrotic fibroblast responses to transforming growth factor-β1. Respir. Res. 2019, 20, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaru, A.; Gabazza, E.; Kobayashi, T.; Kobayashi, H.; Taguchi, O.; Takagi, T.; Oonishi, M.; Fujiwara, K.; Gabazza, C.D.A.; Takahashi, Y. Matrix metalloproteinase-2 is protective in bleomycin-induced pulmonary fibrosis. Eur. Respir. J. 2015, 46, 1903. [Google Scholar]
- Kubben, F.J.; Peeters-Haesevoets, A.; Engels, L.G.; Baeten, C.G.; Schutte, B.; Arends, J.W.; Stockbrügger, R.W.; Blijham, G.H. Proliferating cell nuclear antigen (PCNA): A new marker to study human colonic cell proliferation. Gut 1994, 35, 530–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Silva, M.A.; Pires, K.M.; Trajano, E.T.; Martins, V.; Nesi, R.T.; Benjamin, C.F.; Caetano, M.S.; Sternberg, C.; Machado, M.N.; Zin, W.A.; et al. Redox imbalance and pulmonary function in bleomycin-induced fibrosis in C57BL/6, DBA/2, and BALB/c mice. Toxicol. Pathol. 2012, 40, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Xiong, M.; Zhao, Y.; Mo, H.; Yang, H.; Yue, F.; Hu, K. Intermittent hypoxia increases ROS/HIF-1α ‘related oxidative stress and inflammation and worsens bleomycin-induced pulmonary fibrosis in adult male C57BL/6J mice. Int. Immunopharmacol. 2021, 100, 108165. [Google Scholar] [CrossRef]
- Zitka, O.; Skalickova, S.; Gumulec, J.; Masarik, M.; Adam, V.; Hubalek, J.; Trnkova, L.; Kruseova, J.; Eckschlager, T.; Kizek, R. Redox status expressed as GSH:GSSG ratio as a marker for oxidative stress in paediatric tumour patients. Oncol. Lett. 2012, 4, 1247–1253. [Google Scholar] [CrossRef] [Green Version]
- Drozd, E.; Gruber, B.; Marczewska, J.; Drozd, J.; Anuszewska, E. Intracellular glutathione level and efflux in human melanoma and cervical cancer cells differing in doxorubicin resistance. Postepy Hig. Med. Dosw. 2016, 70, 319–328. [Google Scholar] [CrossRef]
- Petruska, J.M.; Leslie, K.O.; Mossman, B.T. Enhanced lipid peroxidation in lung lavage of rats after inhalation of asbestos. Free Radic. Biol. Med. 1991, 11, 425–432. [Google Scholar] [CrossRef]
- Rahman, I.; van Schadewijk, A.A.; Crowther, A.J.; Hiemstra, P.S.; Stolk, J.; MacNee, W.; De Boer, W.I. 4-Hydroxy-2-nonenal, a specific lipid peroxidation product, is elevated in lungs of patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2002, 166, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Leonarduzzi, G.; Scavazza, A.; Biasi, F.; Chiarpotto, E.; Camandola, S.; Vogel, S.; Dargel, R.; Poli, G. The lipid peroxidation end product 4-hydroxy-2,3-nonenal up-regulates transforming growth factor beta1 expression in the macrophage lineage: A link between oxidative injury and fibrosclerosis. Faseb J. 1997, 11, 851–857. [Google Scholar] [CrossRef]
- Hirata, Y.; Kuwabara, K.; Takashima, M.; Murai, T. Hormetic Effects of Binaphthyl Phosphonothioates as Pro-oxidants and Antioxidants. Chem. Res. Toxicol. 2020, 33, 2892–2902. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Garzón, V.R.; Arellanes-Robledo, J.; García-Román, R.; Aparicio-Rautista, D.I.; Villa-Treviño, S. Inhibition of reactive oxygen species and pre-neoplastic lesions by quercetin through an antioxidant defense mechanism. Free Radic. Res. 2009, 43, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Arbeeny, C.M.; Ling, H.; Smith, M.M.; O’Brien, S.; Wawersik, S.; Ledbetter, S.R.; McAlexander, A.; Schopfer, F.J.; Willette, R.N.; Jorkasky, D.K. CXA-10, a Nitrated Fatty Acid, Is Renoprotective in Deoxycorticosterone Acetate-Salt Nephropathy. J. Pharmacol. Exp. Ther. 2019, 369, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.G.; Lee, T.H.; Hong, Y.; Ryoo, J.; Heo, J.W.; Gil, B.M.; Kang, H.S.; Kwon, S.S.; Kim, Y.H. Effects of low-dose pirfenidone on survival and lung function decline in patients with idiopathic pulmonary fibrosis (IPF): Results from a real-world study. PLoS ONE 2021, 16, e0261684. [Google Scholar] [CrossRef]
- Lee, R.; Reese, C.; Bonner, M.; Tourkina, E.; Hajdu, Z.; Riemer, E.C.; Silver, R.M.; Visconti, R.P.; Hoffman, S. Bleomycin delivery by osmotic minipump: Similarity to human scleroderma interstitial lung disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L736–L748. [Google Scholar] [CrossRef] [Green Version]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-García, K.; López-Martínez, A.; Velázquez-Enríquez, J.M.; Zertuche-Martínez, C.; Carrasco-Torres, G.; Sánchez-Navarro, L.M.; Villa-Treviño, S.; Baltiérrez-Hoyos, R.; Vásquez-Garzón, V.R. 3′5-Dimaleamylbenzoic Acid Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Int. J. Mol. Sci. 2022, 23, 7943. https://doi.org/10.3390/ijms23147943
González-García K, López-Martínez A, Velázquez-Enríquez JM, Zertuche-Martínez C, Carrasco-Torres G, Sánchez-Navarro LM, Villa-Treviño S, Baltiérrez-Hoyos R, Vásquez-Garzón VR. 3′5-Dimaleamylbenzoic Acid Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. International Journal of Molecular Sciences. 2022; 23(14):7943. https://doi.org/10.3390/ijms23147943
Chicago/Turabian StyleGonzález-García, Karina, Armando López-Martínez, Juan Manuel Velázquez-Enríquez, Cecilia Zertuche-Martínez, Gabriela Carrasco-Torres, Luis Manuel Sánchez-Navarro, Saúl Villa-Treviño, Rafael Baltiérrez-Hoyos, and Verónica Rocío Vásquez-Garzón. 2022. "3′5-Dimaleamylbenzoic Acid Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice" International Journal of Molecular Sciences 23, no. 14: 7943. https://doi.org/10.3390/ijms23147943