
Nutritional Calcium Supply Dependent Calcium Balance, Bone Calcification and Calcium Isotope Ratios in Rats

 , , ,
, , ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Total Body Ca Balance
2.2. Femur Ca Content
2.3. Tibia Micro Computed Tomography (µCT)
2.4. Tibia Histomorphometry
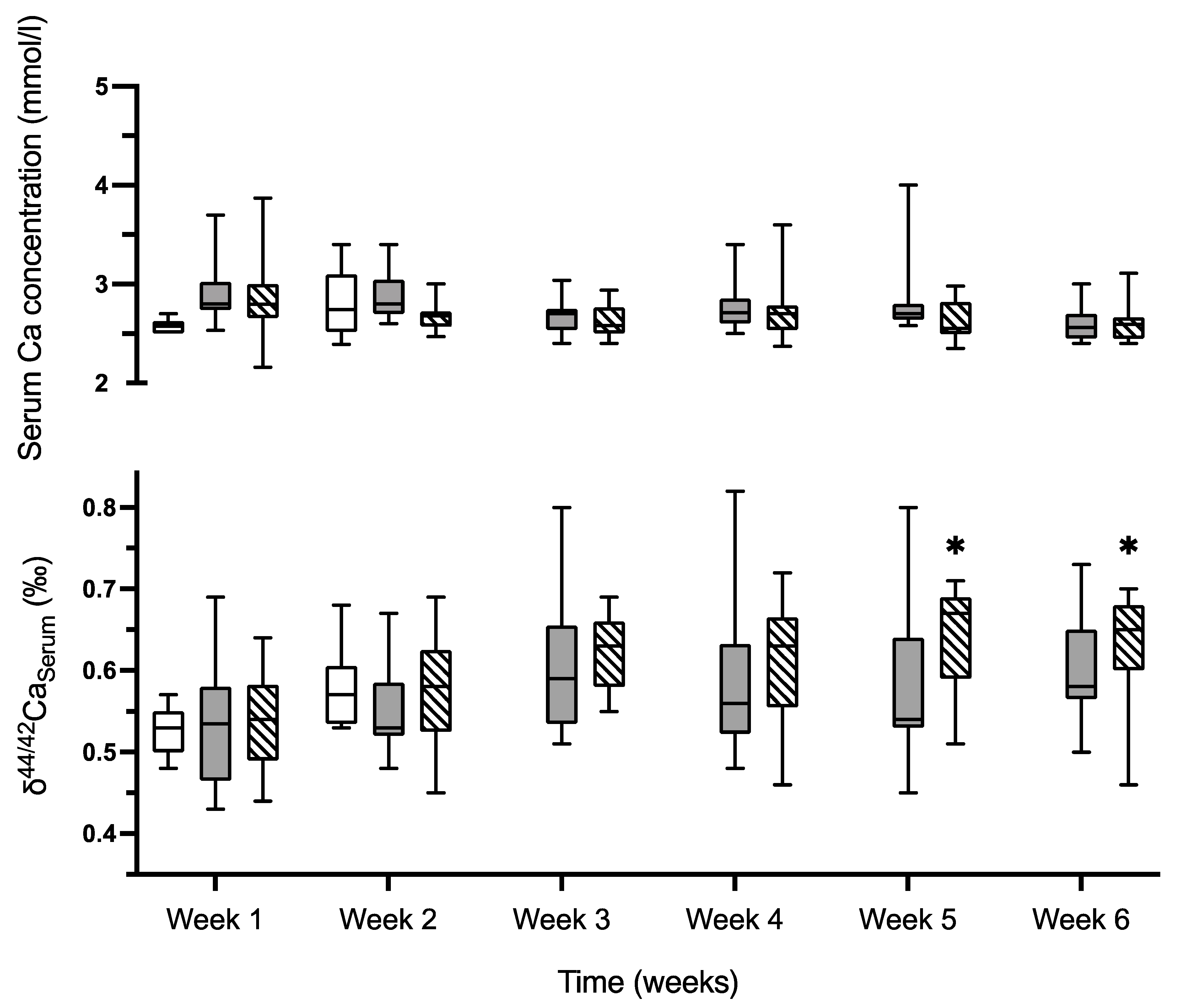
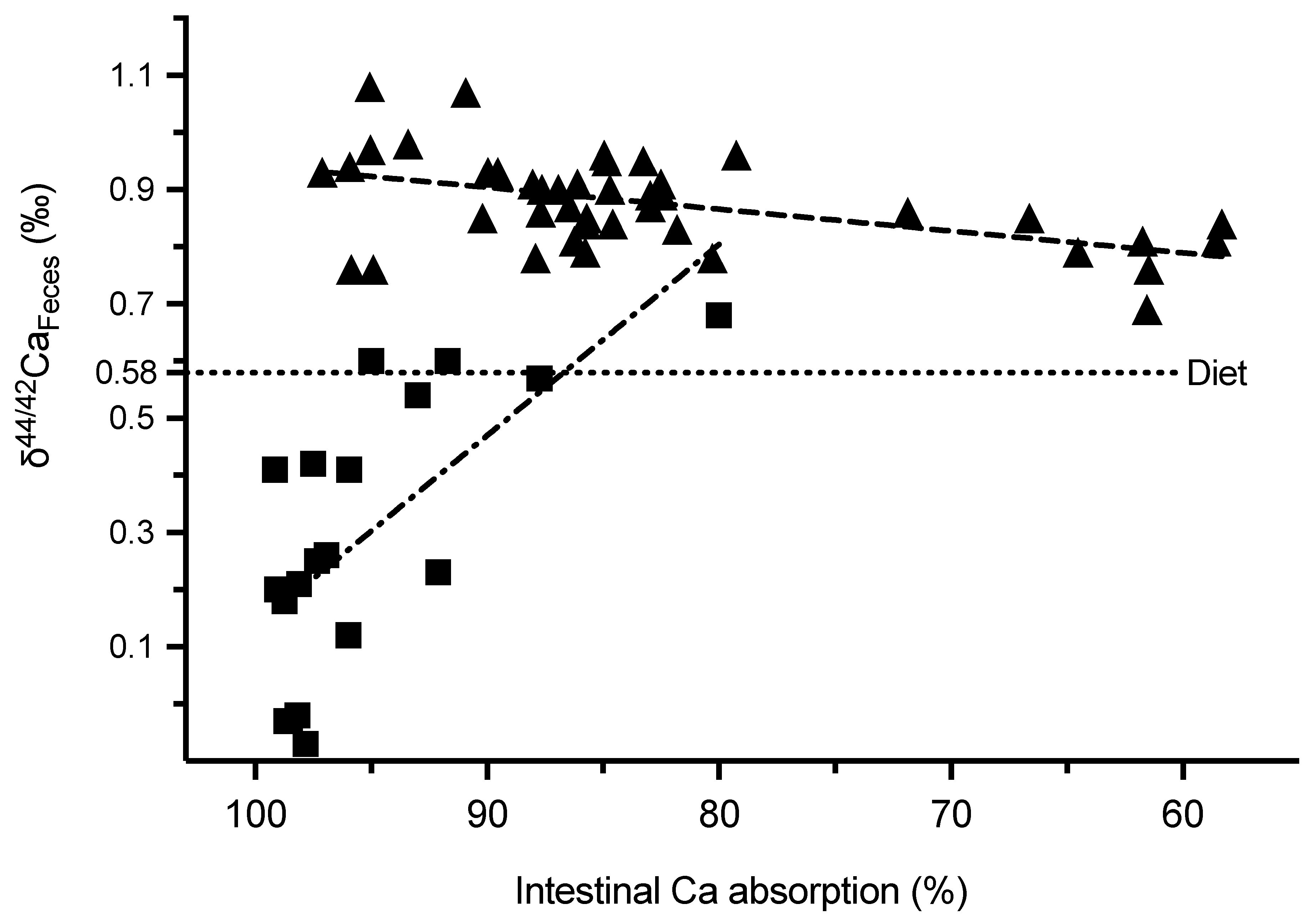
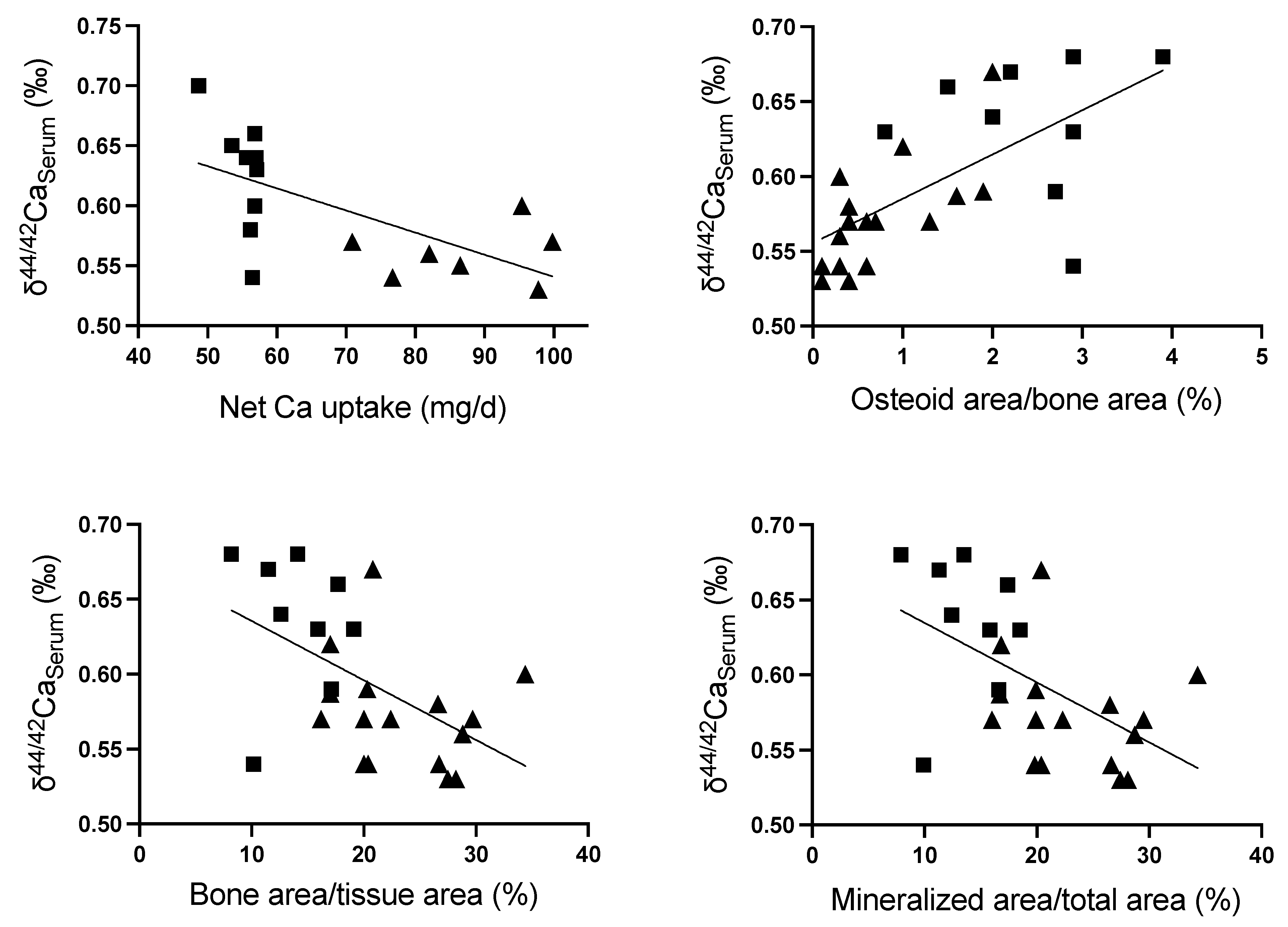
2.5. δ44/42Ca Isotope Studies
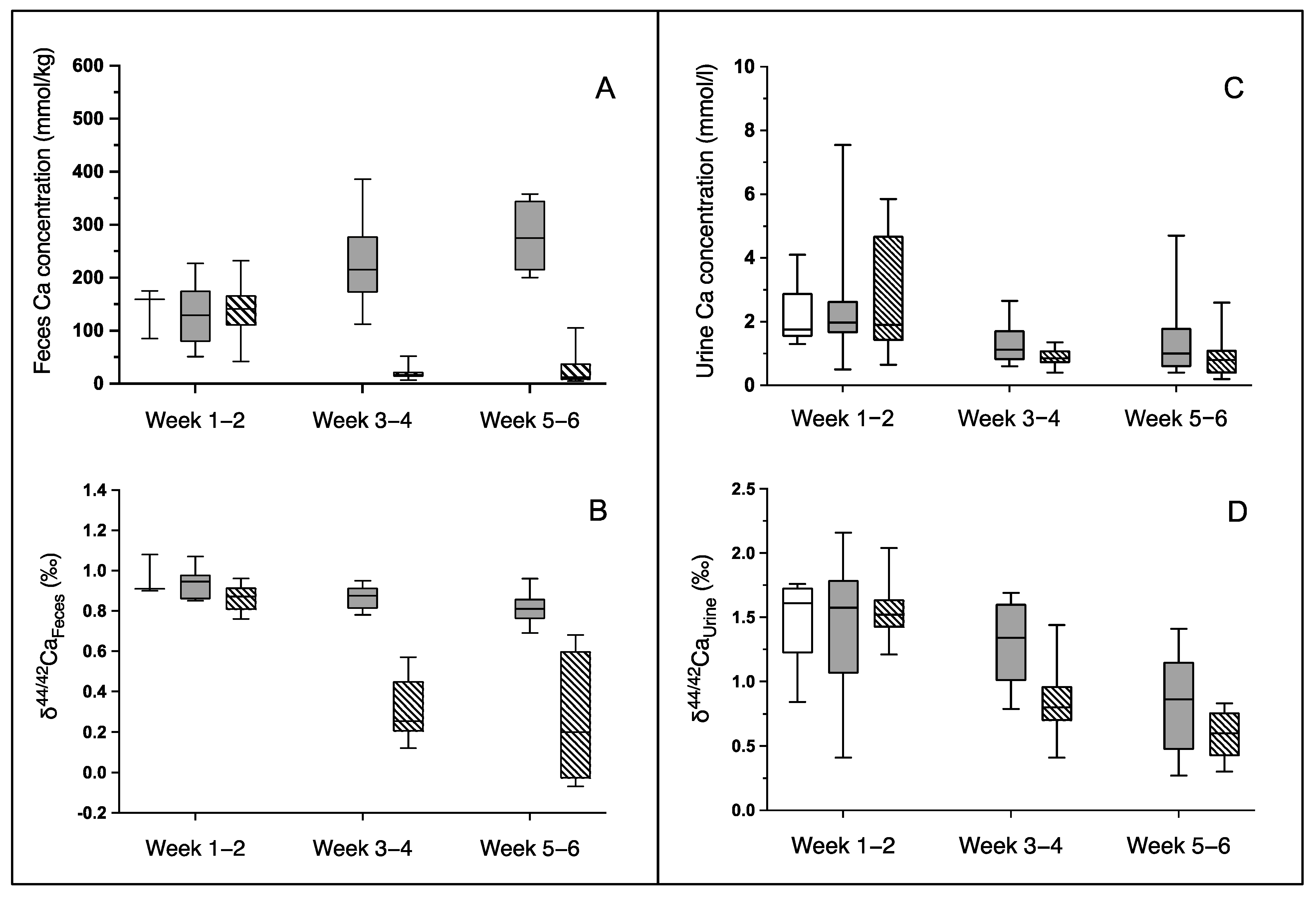
2.5.1. δ44/42Ca in Serum, Feces and Urine
2.5.2. δ44/42Ca in Bone
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Protocol
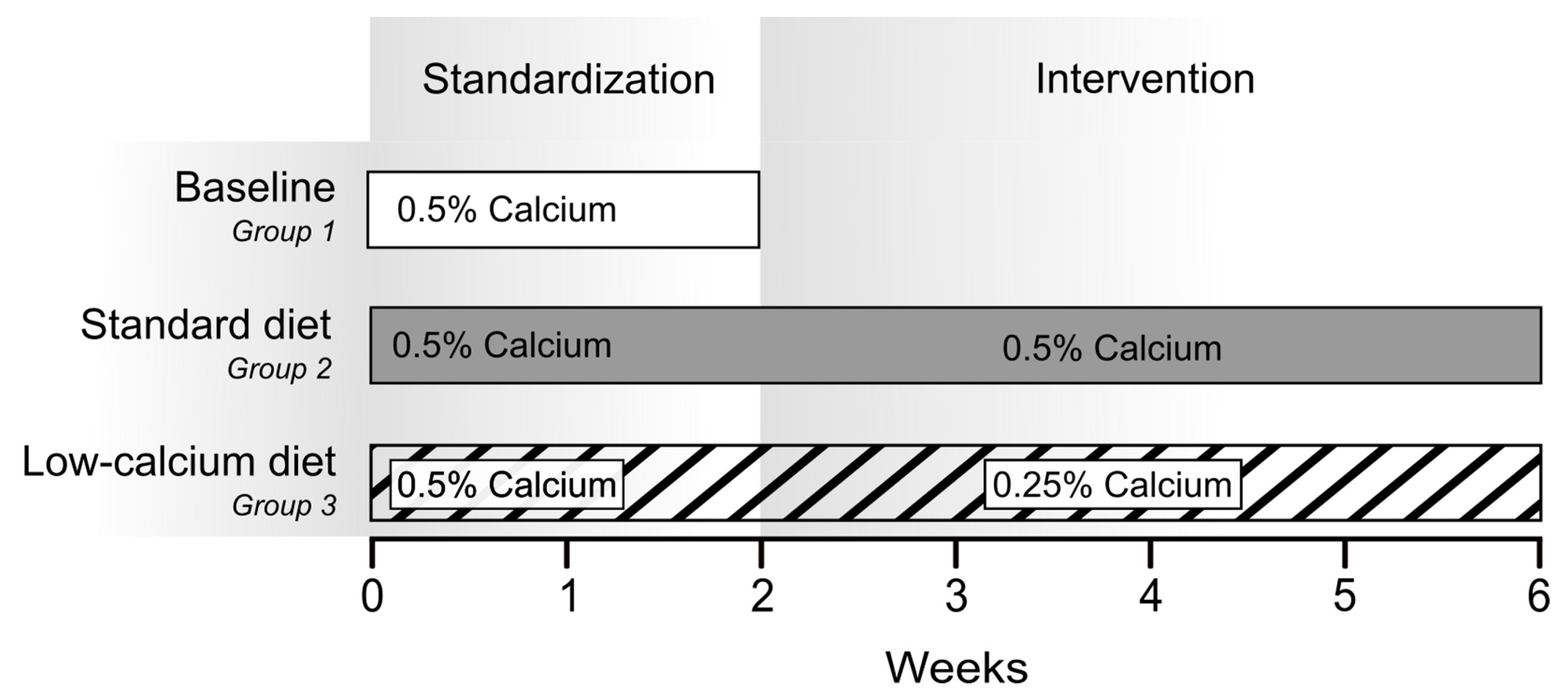
4.3. Methods
4.3.1. Bone Ca Quantification
4.3.2. Computed Tomography Imaging
4.3.3. Bone Histology
4.3.4. Ca Isotope Determination
Sample Digestion and Separation
Mass Spectrometry and Data Evaluation
4.4. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lüllmann-Rauch, R.; Asan, E. Taschenlehrbuch Histologie; Georg Thieme Verlag: Stuttgart, Germany, 2019. [Google Scholar]
- Skulan, J.; DePaolo, D.J. Calcium isotope fractionation between soft and mineralized tissues as a monitor of calcium use in vertebrates. Proc. Natl. Acad. Sci. USA 1999, 96, 13709–13713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuser, A.; Eisenhauer, A. A pilot study on the use of natural calcium isotope (44Ca/40Ca) fractionation in urine as a proxy for the human body calcium balance. Bone 2010, 46, 889–896. [Google Scholar] [CrossRef]
- Bartl, R.; Bartl, C. Bone Disorders: Biology, Diagnosis, Prevention, Therapy; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Gussone, N.; Dietzel, M.; Heuser, A.; Schiller, M.; Schmitt, A.-D.; Tipper, E.; Wombacher, F. Calcium Stable Isotope Geochemistry. In Advances in Isotope Geochemistry, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef]
- Eisenhauer, A.; Muller, M.; Heuser, A.; Kolevica, A.; Gluer, C.C.; Both, M.; Laue, C.; Hehn, U.V.; Kloth, S.; Shroff, R.; et al. Calcium isotope ratios in blood and urine: A new biomarker for the diagnosis of osteoporosis. Bone Rep. 2019, 10, 100200. [Google Scholar] [CrossRef]
- Hoefs, J. Stable Isotope Geochemistry, 5th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2004; 244p. [Google Scholar]
- Eisenhauer, A.; Nägler, T.F.; Stille, P.; Kramers, J.; Gussone, N.; Bock, B.; Fietzke, J.; Hippler, D.; Schmitt, A.-D. Proposal for International Agreement on Ca Notation Resulting from Discussions at Workshops on Stable Isotope Measurements Held in Davos (Goldschmidt 2002) and Nice (EGS-AGU-EUG 2003). Geostand. Geoanalytical Res. 2004, 28, 149–151. [Google Scholar] [CrossRef]
- Heuser, A.; Eisenhauer, A.; Scholz-Ahrens, K.E.; Schrezenmeir, J. Biological fractionation of stable Ca isotopes in Gottingen minipigs as a physiological model for Ca homeostasis in humans. Isot. Environ. Health Stud. 2016, 52, 633–648. [Google Scholar] [CrossRef]
- Heaney, R. Calcium Kinetics in Plasma: As they Apply to the Measurements of Bone Formation and Resporption Rates. In Calcification and Physiology, 2nd ed.; Bourne, G.H., Ed.; Academic Press: Cambridge, MA, USA, 1976; pp. 105–133. [Google Scholar]
- Ott, S.M.; Murano, R.; Lewellen, T.K.; Nelp, W.B.; Chesnut, C.M., 3rd. Total body calcium by neutron activation analysis in normals and osteoporotic populations: A discriminator of significant bone mass loss. J. Lab. Clin. Med. 1983, 102, 637–645. [Google Scholar] [PubMed]
- Heuser, A.; Frings-Meuthen, P.; Rittweger, J.; Galer, S.J.G. Calcium Isotopes in Human Urine as a Diagnostic Tool for Bone Loss: Additional Evidence for Time Delays in Bone Response to Experimental Bed Rest. Front. Physiol. 2019, 10, 12. [Google Scholar] [CrossRef]
- Channon, M.B.; Gordon, G.W.; Morgan, J.L.; Skulan, J.L.; Smith, S.M.; Anbar, A.D. Using natural, stable calcium isotopes of human blood to detect and monitor changes in bone mineral balance. Bone 2015, 77, 69–74. [Google Scholar] [CrossRef]
- Skulan, J.; Bullen, T.; Anbar, A.D.; Puzas, J.E.; Shackelford, L.; LeBlanc, A.; Smith, S.M. Natural calcium isotopic composition of urine as a marker of bone mineral balance. Clin. Chem. 2007, 53, 1155–1158. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.L.; Gordon, G.W.; Arrua, R.C.; Skulan, J.L.; Anbar, A.D.; Bullen, T.D. High-precision measurement of variations in calcium isotope ratios in urine by multiple collector inductively coupled plasma mass spectrometry. Anal. Chem. 2011, 83, 6956–6962. [Google Scholar] [CrossRef]
- Schreiber, J.J.; Anderson, P.A.; Hsu, W.K. Use of computed tomography for assessing bone mineral density. Neurosurg. Focus 2014, 37, E4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, J.J.; Anderson, P.A.; Rosas, H.G.; Buchholz, A.L.; Au, A.G. Hounsfield units for assessing bone mineral density and strength: A tool for osteoporosis management. J. Bone Jt. Surg. Am. 2011, 93, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Gordon, G.W.; Monge, J.; Channon, M.B.; Wu, Q.; Skulan, J.L.; Anbar, A.D.; Fonseca, R. Predicting multiple myeloma disease activity by analyzing natural calcium isotopic composition. Leukemia 2014, 28, 2112–2115. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.; Skulan, J.L.; Gordon, G.W.; Romaniello, S.J.; Smith, S.M.; Anbar, A.D. Rapidly assessing changes in bone mineral balance using natural stable calcium isotopes. Proc. Natl. Acad. Sci. USA 2012, 109, 9989–9994. [Google Scholar] [CrossRef] [Green Version]
- Rangarajan, R.; Mondal, S.; Thankachan, P.; Chakrabarti, R.; Kurpad, A.V. Assessing bone mineral changes in response to vitamin D supplementation using natural variability in stable isotopes of Calcium in Urine. Sci. Rep. 2018, 8, 16751. [Google Scholar] [CrossRef] [Green Version]
- Shroff, R.; Fewtrell, M.; Heuser, A.; Kolevica, A.; Lalayiannis, A.; McAlister, L.; Silva, S.; Goodman, N.; Schmitt, C.P.; Biassoni, L.; et al. Naturally Occurring Stable Calcium Isotope Ratios in Body Compartments Provide a Novel Biomarker of Bone Mineral Balance in Children and Young Adults. J. Bone Miner. Res. 2021, 36, 133–142. [Google Scholar] [CrossRef]
- Toepfer, E.T.; Rott, J.; Bartosova, M.; Kolevica, A.; Machuca-Gayet, I.; Heuser, A.; Rabe, M.; Shroff, R.; Bacchetta, J.; Zarogiannis, S.G.; et al. Calcium isotope fractionation by osteoblasts and osteoclasts, across endothelial and epithelial cell barriers, and with binding to proteins. Am. J. Physiol. Integr. Comp. Physiol. 2021, 321, R29–R40. [Google Scholar] [CrossRef]
- Christian, M.S.; Hoberman, A.M.; Johnson, M.D.; Brown, W.R.; Bucci, T.J. Effect of dietary optimization on growth, survival, tumor incidences and clinical pathology parameters in CD Sprague-Dawley and Fischer-344 rats: A 104-week study. Drug Chem. Toxicol. 1998, 21, 97–117. [Google Scholar] [CrossRef]
- Chu, N.; Henderson, G.; Hedges, R. Ca isotope variations in modern dietary systems and their potential to assess the importance dairying in past cultures. Geophys. Res. Abstr. 2005, 7, 047426. [Google Scholar]
- Chu, N.-C.; Henderson, G.M.; Belshaw, N.S.; Hedges, R.E.M. Establishing the potential of Ca isotopes as proxy for consumption of dairy products. Appl. Geochem. 2006, 21, 1656–1667. [Google Scholar] [CrossRef]
- Bronner, F. Mechanisms of intestinal calcium absorption. J. Cell. Biochem. 2003, 88, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.; Nilius, B.; Bindels, R.J. Calcium absorption across epithelia. Physiol. Rev. 2005, 85, 373–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, D.A.; Miller, D.H. Extrusion of colonic epithelial cells in vitro. J. Electron Microsc. Tech. 1990, 16, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Heaney, R.P.; Abrams, S.A. Improved estimation of the calcium content of total digestive secretions. J. Clin. Endocrinol. Metab. 2004, 89, 1193–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lieshout, E.M.; Van Doesburg, W.; Van der Meer, R. Real-time PCR of host DNA in feces to study differential exfoliation of colonocytes between rats and humans. Scand. J. Gastroenterol. 2004, 39, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Beggs, M.R.; Alexander, R.T. Intestinal absorption and renal reabsorption of calcium throughout postnatal development. Exp. Biol. Med. 2017, 242, 840–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronner, F. Calcium absorption—A paradigm for mineral absorption. J. Nutr. 1998, 128, 917–920. [Google Scholar] [CrossRef]
- Gloux, A.; Le Roy, N.; Brionne, A.; Bonin, E.; Juanchich, A.; Benzoni, G.; Piketty, M.L.; Prie, D.; Nys, Y.; Gautron, J.; et al. Candidate genes of the transcellular and paracellular calcium absorption pathways in the small intestine of laying hens. Poult. Sci. 2019, 98, 6005–6018. [Google Scholar] [CrossRef]
- Bronner, F.; Pansu, D.; Stein, W.D. An analysis of intestinal calcium transport across the rat intestine. Am. J. Physiol. 1986, 250, G561–G569. [Google Scholar] [CrossRef]
- Belkacemi, L.; Gariepy, G.; Mounier, C.; Simoneau, L.; Lafond, J. Calbindin-D9k (CaBP9k) localization and levels of expression in trophoblast cells from human term placenta. Cell Tissue Res. 2004, 315, 107–117. [Google Scholar] [CrossRef]
- Edwards, A. Regulation of calcium reabsorption along the rat nephron: A modeling study. Am. J. Physiol. Renal Physiol. 2015, 308, F553–F566. [Google Scholar] [CrossRef] [PubMed]
- Granjon, D.; Bonny, O.; Edwards, A. A model of calcium homeostasis in the rat. Am. J. Physiol. Renal Physiol. 2016, 311, F1047–F1062. [Google Scholar] [CrossRef] [PubMed]
- Skulan, J.; DePaolo, D.J.; Owens, T.L. Biological control of calcium isotopic abundances in the global calcium cycle. Geochim. Cosmochim. Acta 1997, 61, 2505–2510. [Google Scholar] [CrossRef]
- Tanaka, Y.K.; Yajima, N.; Higuchi, Y.; Yamato, H.; Hirata, T. Calcium isotope signature: New proxy for net change in bone volume for chronic kidney disease and diabetic rats. Metallomics 2017, 9, 1745–1755. [Google Scholar] [CrossRef]
- Ekbote, V.H.; Khadilkar, A.V.; Khadilkar, V.V.; Chiplonkar, S.A.; Mughal, Z. Dietary patterns with special reference to calcium intake in 2-16-year-old Urban Western Indian children. Indian J. Public Health 2017, 61, 188–193. [Google Scholar] [CrossRef]
- Kudlacek, S.; Schneider, B.; Peterlik, M.; Leb, G.; Klaushofer, K.; Weber, K.; Woloszczuk, W.; Willvonseder, R.; Austrian Study Group on Normative Values of Bone. Assessment of vitamin D and calcium status in healthy adult Austrians. Eur. J. Clin. Investig. 2003, 33, 323–331. [Google Scholar] [CrossRef]
- Wolfensohn, S.; Lloyd, M. Handbook of Laboratory Animal Management and Welfare, 3rd ed.; Blackwell Pub.: Oxford, UK; Ames, Iowa, 2003; 416p. [Google Scholar]
- Stadtwerke-Heidelberg. Trinkwasserqualität im Versorgungsgebiet Heidelberg Hartwasserzone Nördlich des Neckars. Available online: https://www.swhd.de/wasser (accessed on 27 February 2020).
- Behets, G.J.; Spasovski, G.; Sterling, L.R.; Goodman, W.G.; Spiegel, D.M.; De Broe, M.E.; D’Haese, P.C. Bone histomorphometry before and after long-term treatment with cinacalcet in dialysis patients with secondary hyperparathyroidism. Kidney Int. 2015, 87, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Parfitt, A.M.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R. Bone histomorphometry: Standardization of nomenclature, symbols, and units. Report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 1987, 2, 595–610. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Romaniello, S.J.; Field, M.P.; Smith, H.B.; Gordon, G.W.; Kim, M.H.; Anbar, A.D. Fully automated chromatographic purification of Sr and Ca for isotopic analysis. J. Anal. At. Spectrom. 2015, 30, 1906–1912. [Google Scholar] [CrossRef]
- Wieser, M.E.; Buhl, D.; Bouman, C.; Schwieters, J. High precision calcium isotope ratio measurements using a magnetic sector multiple collector inductively coupled plasma mass spectrometer. J. Anal. At. Spectrom. 2004, 19, 844–851. [Google Scholar] [CrossRef]
- Delwatta, S.L.; Gunatilake, M.; Baumans, V.; Seneviratne, M.D.; Dissanayaka, M.L.B.; Batagoda, S.S.; Udagedara, A.H.; Walpola, P.B. Reference values for selected hematological, biochemical and physiological parameters of Sprague-Dawley rats at the Animal House, Faculty of Medicine, University of Colombo, Sri Lanka. Anim. Model. Exp. Med. 2018, 1, 250–254. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline (Group 1, n = 12) | Standard-Ca-Diet (Group 2, n = 19) | Low-Ca-Diet (Group 3, n = 20) | p-Value a | ||
|---|---|---|---|---|---|
| Week 1–2 | Food intake (g/day) Dietary Ca intake (mg/day) | 22.1 ± 1.0 | 22.3 ± 1.7 | 21.5 ± 0.4 | 0.44 |
| 110.6 ± 5.0 | 111.5 ± 8.3 | 107.4 ± 2.1 | 0.44 | ||
| Fluid intake (mL/day) | 28.8 ± 1.0 | 29.4 ± 1.5 | 29.8 ± 1.2 | 0.38 | |
| Fluid Ca intake (mg/day) | 3.2 ± 0.1 | 3.2 ± 0.2 | 3.3 ± 0.1 | 0.38 | |
| Feces (g/day) Fecal Ca loss (mg/day) | 2.8 ± 0.7 | 2.7 ± 1.0 | 3.0 ± 1.0 | 0.76 | |
| 12.9 ± 6.4 | 12.9 ± 9.7 | 15.2 ± 5.0 | 0.77 | ||
| Intestinal Ca absorption b (%) | 88.6 ± 5.6 | 88.8 ± 8.5 | 86.2 ± 4.5 | 0.68 | |
| Intestinal Ca absorption (mg/day) | 100.9 ± 6.4 | 101.8 ± 9.7 | 95.4 ± 5.4 | 0.18 | |
| Urine excretion (mL/day) Urine Ca excretion (mg/day) | 8.5 ± 3.2 | 7.9 ± 2.8 | 10.2 ± 3.3 | 0.08 | |
| 0.7 ± 0.4 | 1.1 ± 0.9 | 0.8 ± 0.7 | 0.47 | ||
| Net body Ca uptake c (mg/day) | 100.4 ± 6.6 | 100.8 ± 9.8 | 94.6 ± 4.9 | 0.99 | |
| Week 3–4 | Food intake (g/day) Dietary Ca intake (mg/day) | 22.3 ± 1.7 | 21.5 ± 0.7 | 0.40 | |
| 111.5 ± 8.3 | 53.7 ± 1.9 | <0.0001 | |||
| Fluid intake (mL/day) | 38.7 ±1.6 | 39.6 ± 2.2 | 0.10 | ||
| Fluid Ca intake (mg/day) | 4.3 ± 0.2 | 4.4 ± 0.2 | 0.10 | ||
| Feces (g/day) Fecal Ca loss (mg/day) | 2.8 ± 1.1 | 3.2 ± 1.2 | 0.23 | ||
| 25.9 ± 16.2 | 2.7 ± 2.0 | 0.0003 | |||
| Intestinal Ca absorption b (%) | 76.6 ± 14.4 | 95.3 ± 3.4 | 0.0009 | ||
| Intestinal Ca absorption (mg/day) | 88.7 ± 16.7 | 55.3 ± 2.0 | <0.0001 | ||
| Urine excretion (mL/day) Urine Ca excretion (mg/day) | 11.6 ± 6.0 | 13.9 ± 5.4 | 0.22 | ||
| 0.6 ± 0.2 | 0.4 ± 0.1 | 0.09 | |||
| Net body Ca uptake c (mg/day) | 89.4 ± 16.3 | 54.9 ± 1.9 | <0.0001 | ||
| Week 5–6 | Food intake (g/day) Dietary Ca intake (mg) | 22.3 ± 1.7 | 21.5 ± 0.7 | 0.40 | |
| 111.5 ± 8.3 | 53.7 ± 1.9 | <0.0001 | |||
| Water intake (mL/day) | 46.1 ± 1.6 | 446.3 ± 2.9 | 0.67 | ||
| Fluid Ca intake (mg/day) | 5.1 ± 0.2 | 5.1 ± 0.3 | 0.67 | ||
| Feces (g/day) Fecal Ca loss (mg/day) | 2.1 ± 0.9 | 2.6 ± 1.5 | 0.30 | ||
| 25.3 ± 13.5 | 2.7 ± 3.7 | 0.0002 | |||
| Intestinal Ca absorption b (%) | 78.3 ± 11.6 | 95.3 ± 6.2 | 0.001 | ||
| Intestinal Ca absorption (mg/day) | 91.2 ± 13.5 | 56.1 ± 3.7 | <0.0001 | ||
| Urine excretion (mL/day) Urine Ca excretion (mg/day) | 17.2 ± 5.8 | 13.2 ± 6.9 | 0.14 | ||
| 1.0 ± 0.9 | 0.3 ± 0.3 | 0.001 | |||
| Net body Ca uptake c (mg/day) | 90.6 ± 13.5 | 55.7 ± 3.6 | <0.0001 | ||
| Week 1–6 | Net body Ca uptake (mg/6 weeks) | 3931 | 2872 | ||
| Week 3–6 | Net body Ca uptake (mg/4 weeks) | 2520 | 1548 | <0.0001 | |
| Baseline Week 2 (Group 1; n = 6) | Standard-Ca-Diet Week 6 (Group 2, n = 19) | Low-Ca-Diet Week 6 (Group 3, n = 19) | p-Value a | ||
|---|---|---|---|---|---|
| G1 vs. G2 | G2 vs. G3 | ||||
| Femur length (mm) | 32.3 ± 0.4 | 37.5 ± 0.3 | 38.1 ± 0.9 | <0.0001 | 0.17 |
| Femur Ca content (mg/femur) | 81.8 ± 4.5 | 121 ± 15 | 109 ± 16 | <0.0001 | 0.02 |
| Relative Ca content (mg/g bone) | 97.4 ± 5.3 | 104 ± 12 | 92.9 ± 14 | 0.21 | 0.03 |
| ∆ Ca gain femur (mg) | 39.6 ± 15 | 27.0 ± 16 | 0.02 | ||
| Left Tibia | Baseline Week 2 (Group 1, n = 12) | Standard-Ca-Diet Week 6 (Group 2, n = 11) | Low-Ca-Diet Week 6 (Group 3, n = 12) | p-Value a |
|---|---|---|---|---|
| Bone area/Tissue area (%) | 22.5 ± 6.4 | 20.4 ± 5.2 | 14.0 ± 3.2 | 0.002 |
| Trabecular number (/mm) | 5.6 ± 1.2 | 4.1 ± 0.6 | 3.2 ± 0.7 | 0.003 |
| Trabecular separation (mm) | 136 (96–184) | 170 (150–209) | 255 (209–288) | 0.003 b |
| Trabecular thickness (mm) | 51 ± 4.4 | 62 ± 8.6 | 57 ± 5.8 | 0.09 |
| Mineralized area/ total area (%) | 22.4 ± 6.4 | 20.2 ± 5.2 | 13.7 ± 3.1 | 0.001 |
| Osteoid area/ bone area (%) | 0.5 (0.2–1.1) | 0.6 (0.4–0.9) | 2.2 (1.6–2.9) | <0.0001 b |
| Osteoid perimeter/ Total perimeter (%) | 1.4 (0.7–2.5) | 4.1 (2.7–6.1) | 13 (9.4–15) | <0.0001 b |
| Osteoid width (µm) | 4.9 ± 1.5 | 4.5 ± 0.7 | 4.9 ± 1.3 | 0.41 |
| Eroded parameter/ Total parameter (%) | 6.9 ± 3.7 | 5.3 ± 3.0 | 4.8 ± 3.2 | 0.64 |
| Osteoblast perimeter/ Osteoid perimeter (%) | 25.1 ± 28.5 | 19.3 ± 20.7 | 37.7 ± 23.0 | 0.06 |
| Osteoblast perimeter/Total perimeter (%) | 0.4 (0.0–1.5) | 0.7 (0.00–1.4) | 4.2 (1.8–8.3) | 0.0002 b |
| Osteoclast perimeter/ Eroded perimeter (%) | 24.1 ± 12.1 | 17.6 ± 10.4 | 22.1 ± 13.7 | 0.39 |
| Osteoclast perimeter/ Total perimeter (%) | 1.7 ± 1.1 | 1.1 ± 0.0 | 1.2 ± 0.8 | 0.82 |
| (1) | |||||||
|---|---|---|---|---|---|---|---|
| δ44/42CaBone (‰) | Baseline Week 2 (Group 1, n = 9) | Standard-Ca-Diet Week 6 (Group 2, n = 6) | Low-Ca-Diet Week 6 (Group 3, n = 10) | p-Value | |||
| G1 vs. G2 a | G2 vs. G3 a | ||||||
| Weight-bearing bone (femora) | 0.15 ± 0.05 | 0.24 ± 0.02 | 0.29 ± 0.03 | 0.0008 | 0.01 | ||
| non-weight-bearing bone (lowest rib) | 0.13 ± 0.05 | 0.24 ± 0.01 | 0.29 ± 0.03 | 0.0003 | 0.002 | ||
| p-value a | 0.61 | 0.89 | 0.75 | ||||
| (2) | |||||||
| Diaphysis (tibia) | 0.18 ± 0.06 | 0.21 ± 0.09 | 0.28 ± 0.03 | 0.71 | 0.05 | ||
| Metaphysis (tibia) | 0.27 ± 0.07 | 0.26 ± 0.07 | 0.33 ± 0.03 | 0.67 | 0.006 | ||
| Epiphysis (tibia) | 0.26 ± 0.04 | 0.24 ± 0.07 | 0.32 ± 0.03 | 0.71 | 0.009 | ||
| p-value a | Diaphysis vs. Metaphysis | 0.016 | 0.37 | 0.0006 | |||
| Diaphysis vs. Epiphysis | 0.007 | 0.54 | 0.008 | ||||
| Metaphysis vs. Epiphysis | 0.55 | 0.74 | 0.40 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rott, J.; Toepfer, E.T.; Bartosova, M.; Kolevica, A.; Heuser, A.; Rabe, M.; Behets, G.; D’Haese, P.C.; Eichwald, V.; Jugold, M.; et al. Nutritional Calcium Supply Dependent Calcium Balance, Bone Calcification and Calcium Isotope Ratios in Rats. Int. J. Mol. Sci. 2022, 23, 7796. https://doi.org/10.3390/ijms23147796
Rott J, Toepfer ET, Bartosova M, Kolevica A, Heuser A, Rabe M, Behets G, D’Haese PC, Eichwald V, Jugold M, et al. Nutritional Calcium Supply Dependent Calcium Balance, Bone Calcification and Calcium Isotope Ratios in Rats. International Journal of Molecular Sciences. 2022; 23(14):7796. https://doi.org/10.3390/ijms23147796
Chicago/Turabian StyleRott, Jeremy, Eva Teresa Toepfer, Maria Bartosova, Ana Kolevica, Alexander Heuser, Michael Rabe, Geert Behets, Patrick C. D’Haese, Viktoria Eichwald, Manfred Jugold, and et al. 2022. "Nutritional Calcium Supply Dependent Calcium Balance, Bone Calcification and Calcium Isotope Ratios in Rats" International Journal of Molecular Sciences 23, no. 14: 7796. https://doi.org/10.3390/ijms23147796