
Characterization, Expression Profiling, and Biochemical Analyses of the Cinnamoyl-CoA Reductase Gene Family for Lignin Synthesis in Alfalfa Plants


Abstract
:1. Introduction
2. Results
2.1. Identification and Analysis of Cinnamoyl-CoA Reductase (CCR) Genes in M. sativa Genome
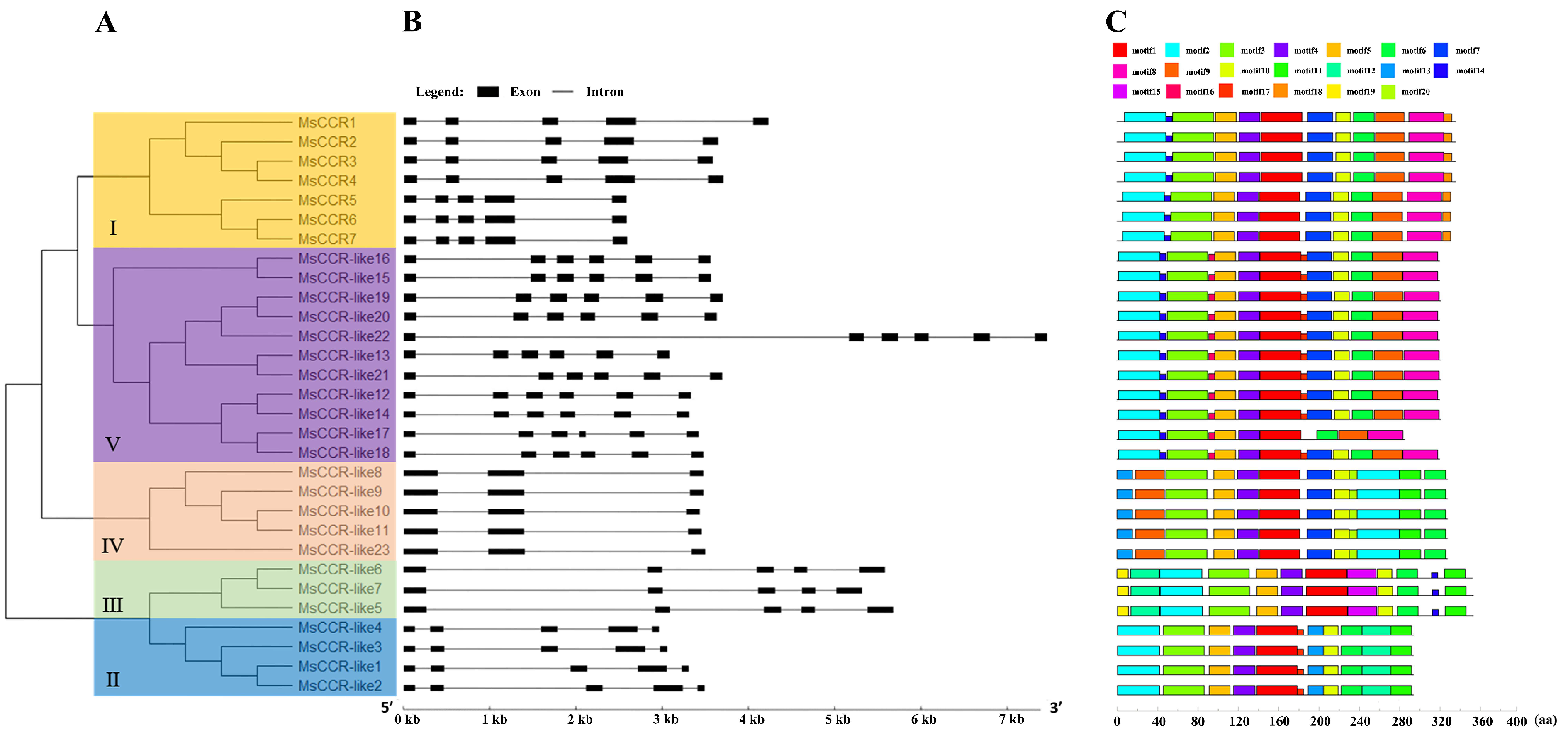
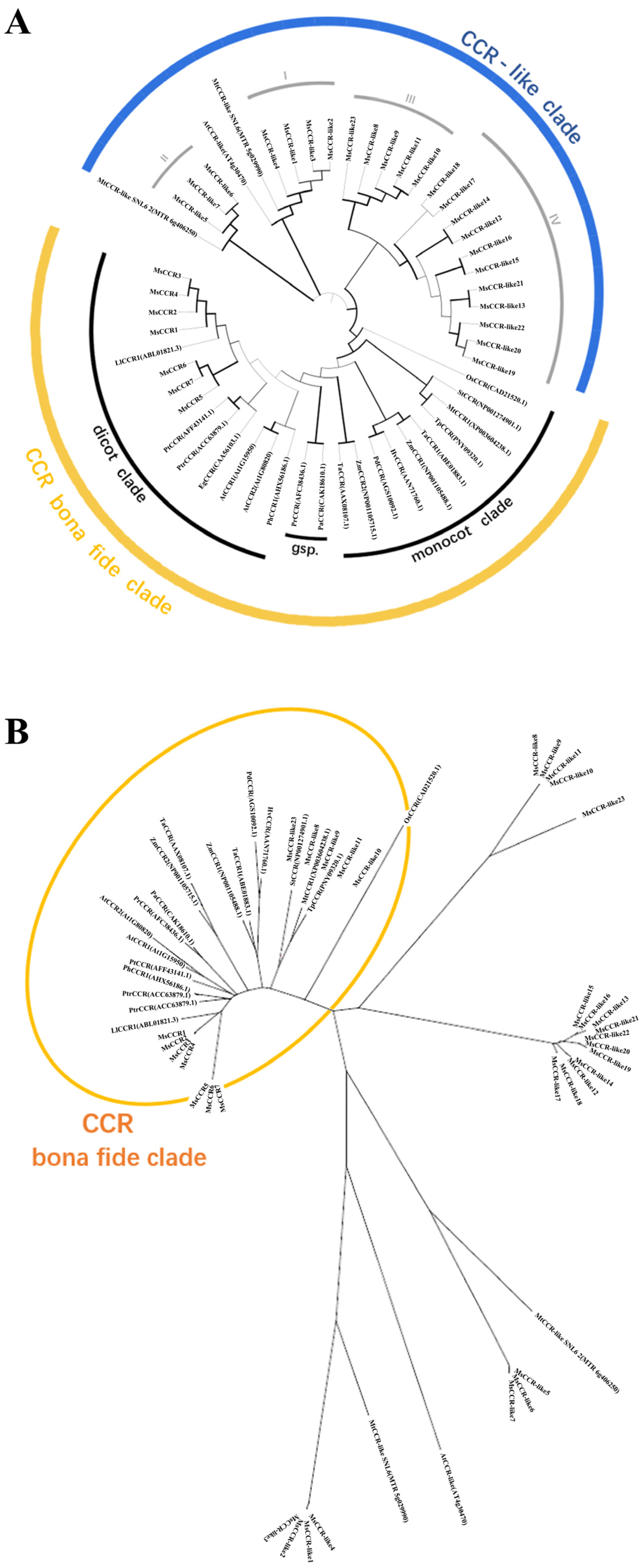
2.2. Gene Structure, Evolution, and Conserved Motif Analysis of MsCCRs/MsCCR-like Genes
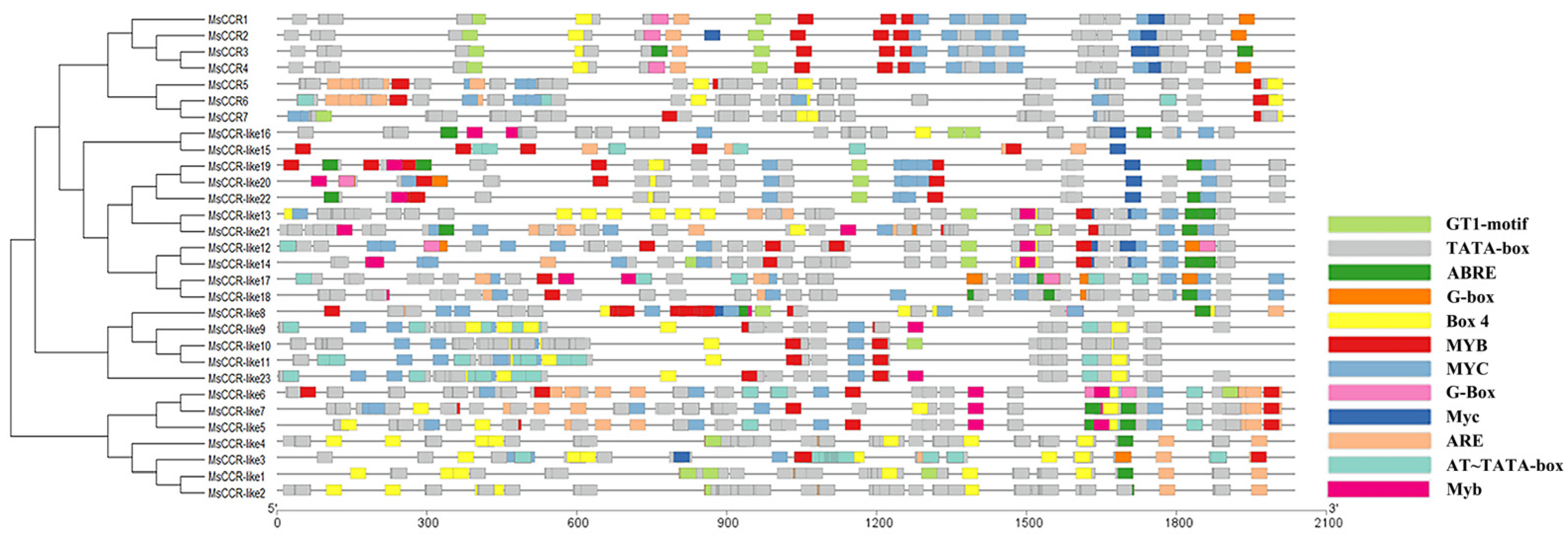
2.3. The Cis-Acting Regulatory Elements in the Promoter of MsCCRs
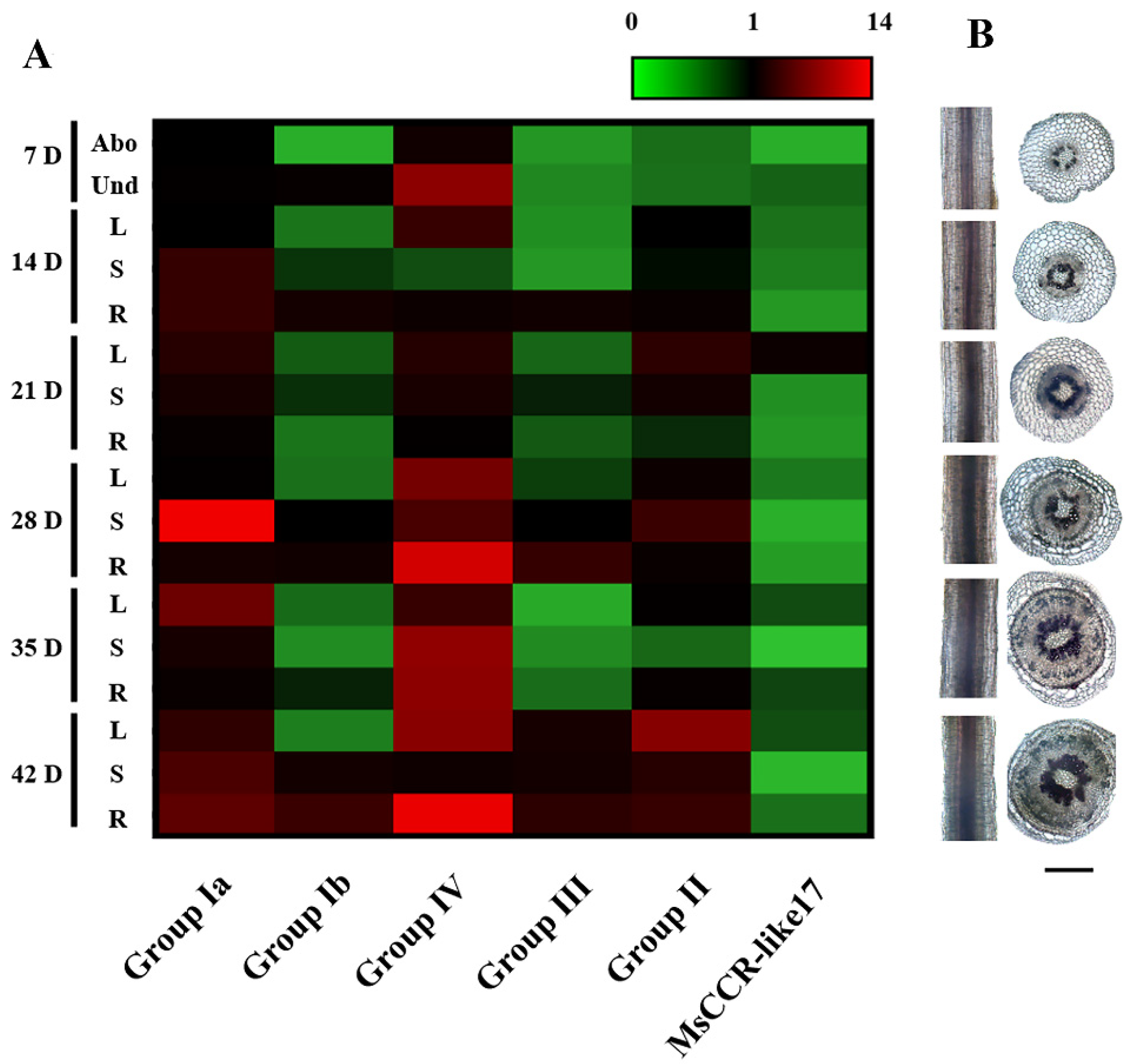
2.4. The Expression Patterns of MsCCR/MsCCR-like Genes and Lignin Content Analysis during Different Growth Stage
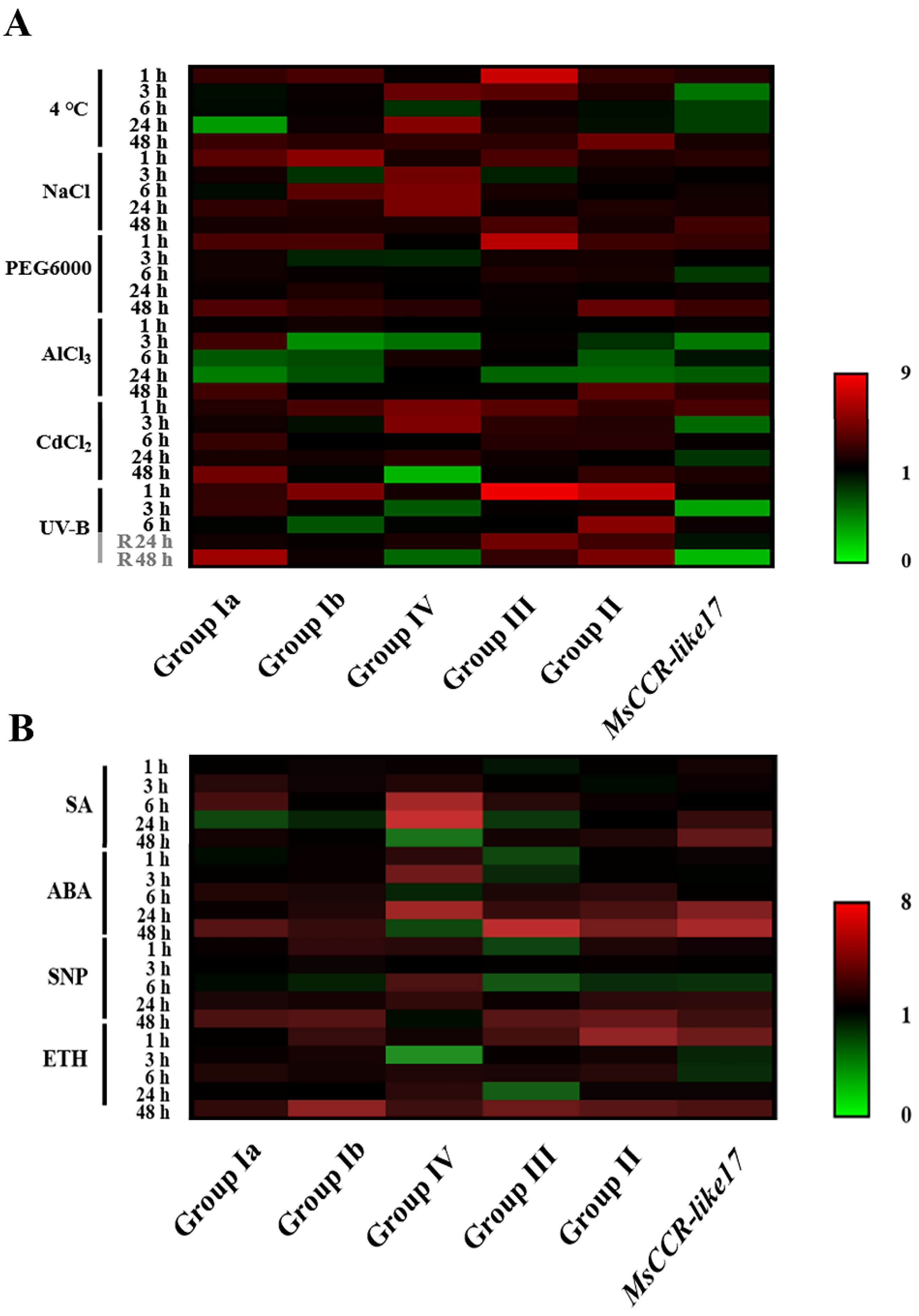
2.5. The Expression Patterns of MsCCRs/MsCCR-likes under Abiotic Stresses and Hormone Treatment
2.6. Cloning, Heterologous Expression, and Enzymatic Assay of MsCCR1 and MsCCR-like1 Gene
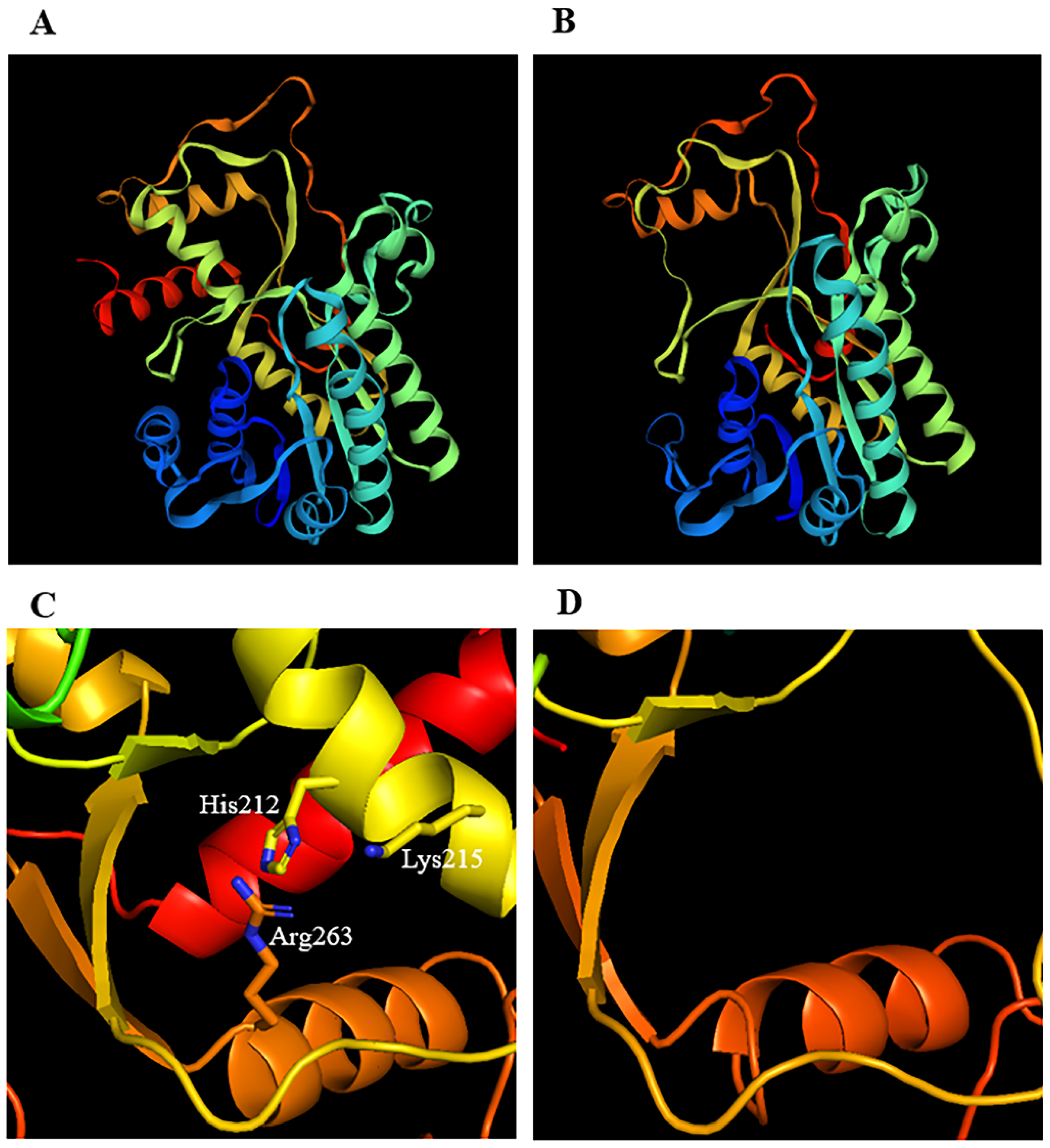
2.7. Three-Dimensional Structure Analysis of CCRs in M. sativa and Other Plants
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. MsCCR Identification and Classification
4.3. Motif Analysis and Gene Structure Visualization
4.4. Multiple Sequence Alignment and Phylogenic Analysis Evolutionary Analysis
4.5. Cis-regulatory Element Analysis and 3D Structure Prediction
4.6. Collinearity Analysis and Chromosome Localization
4.7. RNA Isolation and RT-qPCR Analysis
4.8. Lignin Content and Staining Analysis
4.9. Cloning, Expression, and Purification of Recombinant MsCCR
4.10. CCR Activity and Enzyme Kinetic Parameter Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prasad, N.; Vindal, V.; Kumar, V.; Kabra, A.; Phogat, N.; Kujmar, M. Structural and docking studies of Leucaena leucocephala cinnamoyl CoA reductase. J. Mol. Model. 2011, 17, 533–541. [Google Scholar] [CrossRef]
- Jung, H.J.G.; Samac, D.A.; Sarath, G. Modifying crops to increase cell wall digestibility. Plant Sci. 2012, 185, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, J.; Temple, S.; Dixon, R.A. Development and commercialization of reduced lignin alfalfa. Curr. Opin. Biotech. 2019, 56, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Auh, C.K.; Dowling, P.; Bell, J.; Lehmann, D.; Wang, Z.Y. Transgenic down-regulation of caffeic acid O-methyltransferase (COMT) led to improved digestibility in tall fescue (Festuca arundinacea). Funct. Plant Biol. 2004, 31, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Grev, A.M.; Hathaway, M.R.; Sheaffer, C.C.; Wells, M.S.; Reiter, A.S.; Martinson, K.L. Apparent digestibility, fecal particle size, and mean retention time of reduced lignin alfalfa hay fed to horses. J. Anim. Sci. 2021, 99, skab158. [Google Scholar] [CrossRef]
- Reddy, M.S.; Chen, F.; Shadle, G.; Jackson, L.; Aljoe, H.; Dixon, R.A. Targeted down-regulation of cytochrome P450 enzymes for forage quality improvement in alfalfa (Medicago sativa L.). Proc. Natl. Acad. Sci. USA 2005, 102, 16573–16578. [Google Scholar] [CrossRef] [Green Version]
- Carocha, V.; Soler, M.; Hefer, C.; Cassan-Wang, H.; Fevereiro, P.; Myburg, A.A.; Paiva, J.A.P.; Grima-Pettenati, J.G. Genome-wide analysis of the lignin toolbox of Eucalyptus grandis. New Phytol. 2015, 206, 1297–1313. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Dixon, R.A. Lignin modification improves fermentable sugar yields for biofuel production. Nat. Biotechnol. 2007, 25, 759–761. [Google Scholar] [CrossRef]
- Simmons, B.A.; Loqué, D.; Ralph, J. Advances in modifying lignin for enhanced biofuel production. Curr. Opin. Plant Biol. 2010, 13, 313–320. [Google Scholar] [CrossRef]
- Umezawa, T. Lignin modification in planta for valorization. Phytochem. Rev. 2018, 17, 1305–1327. [Google Scholar] [CrossRef]
- Hisano, H.; Nandakumar, R.; Wang, Z.Y. Genetic modification of lignin biosynthesis for improved biofuel production. In Vitro Cell. Dev. Biol. 2009, 45, 306–313. [Google Scholar] [CrossRef]
- De Meester, B.; Calderón, B.M.; de Vries, L.; Pollier, J.; Goeminne, G.; Van Doorsselaere, J.; Chen, M.; Ralph, J.; Vanholme, R.; Boerjan, W. Tailoring poplar lignin without yield penalty by combining a null and haploinsufficient CINNAMOYL-CoA REDUCTASE2 allele. Nat. Commun. 2020, 11, 5020. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, J.; Serk, H.; Granlund, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.; Ennos, A.R.; Turner, S.R. Cloning and characterization of irregular xylem4 (irx4): A severely lignin-deficient mutant of Arabidopsis. Plant J. 2001, 26, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Khakdan, F.; Nasiri, J.; Ranjbar, M.; Alizadeh, H. Water deficit stress fluctuates expression profiles of 4Cl, C3H, COMT, CVOMT and EOMT genes involved in the biosynthesis pathway of volatile phenylpropanoids alongside accumulation of methylchavicol and methleugenol in different Iranian cultivars of basil. J. Plant Physiol. 2017, 218, 74–83. [Google Scholar] [CrossRef]
- Ali, M.B.; Singh, N.; Shohael, A.M.; Hahn, E.J.; Paek, K.Y. Phenolics metabolism and lignin synthesis in root suspension cultures of Panax ginseng in response to copper stress. Plant Sci. 2006, 171, 147–154. [Google Scholar] [CrossRef]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species- and jasmonic acid-dependent process in Arabidopsis. Plant Physiol. 2011, 156, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Liu, X.B.; Shen, H.Z.; Gao, Z.; Ruan, R.W.; Wu, D.Q.; Yi, Z.L. The lignin synthesis related genes and lodging resistance of Fagopyrum esculentum. Biol. Plantarum. 2017, 61, 138–146. [Google Scholar] [CrossRef]
- Begović, L.; Abičić, I.; Lalić, A.; Lepeduš, H.; Cesar, V.; Leljak-Levanić, D. Lignin synthesis and accumulation in barley cultivars differing in their resistance to lodging. Plant Physiol. Biochem. 2018, 133, 142–148. [Google Scholar] [CrossRef]
- Baxter, H.L.; Stewart, N., Jr. Effects of altered lignin biosynthesis on phenylpropanoid metabolism and plant stress. Biofuels 2013, 4, 635–650. [Google Scholar] [CrossRef]
- Gou, M.; Yang, X.; Zhao, Y.; Ran, X.; Song, Y.; Liu, C.J. Cytochrome b5 is an obligate electron shuttle protein for syringyl lignin biosynthesis in Arabidopsis. Plant Cell 2019, 31, 1344–1366. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.L.; Bhoo, S.H.; Kwon, M.; Lee, S.W.; Cho, M.H. Biochemical and expression analyses of the rice cinamoyl-CoA reductase gene family. Front. Plant Sci. 2017, 8, 2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, J.; Chen, F.; Jackson, L.; Shadle, G.; Dixon, R. Multi-site genetic modification of monolignols biosynthesis in alfalfa (Medicago sativa): Effects on lignin composition in specific cell types. New Phytol. 2008, 179, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Tamasloukht, B.; Lam, M.S.-J.W.Q.; Martinez, Y.; Tozo, K.; Barbier, O.; Jourda, C.; Jauneau, A.; Borderies, G.; Balzergue, S.; Renou, J.P.; et al. Characterization of a cinnamoyl-CoA reductase 1 (CCR1) mutant in maize: Effects on lignification, fibre development, and global gene expression. J. Exp. Bot. 2011, 62, 3837–3848. [Google Scholar] [CrossRef]
- Tu, Y.; Rochfort, S.; Liu, Z.; Ran, Y.; Griffith, M.; Badenhorst, P.; Louie, G.V.; Bowman, M.E.; Smith, K.F.; Noel, J.P.; et al. Functional analyses of caffeic acid O-methyltransferase and cinnamoyl-CoA-reductase genes from perennial ryegrass (Lolium perenne). Plant Cell 2010, 22, 3357–3373. [Google Scholar] [CrossRef] [Green Version]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E.; Poby, D.; Grima Pettenati, J. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar] [CrossRef]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van de Peer, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [Green Version]
- Thévenin, J.; Pollet, B.; Letarnec, B.; Saulnier, L.; Gissot, L.; Maia-Grondard, A.; Lapierre, C.; Jounanin, L. The simultaneous repression of CCR and CAD, two enzymes of the lignin biosynthetic pathway, results in sterility and dwarfism in Arabidopsis thaliana. Mol. Plant 2011, 4, 70–82. [Google Scholar] [CrossRef]
- De Meester, B.; de Vries, L.; Özparpucu, M.; Gierlinger, N.; Corneillie, S.; Pallidis, A.; Goeminne, G.; Morreel, K.; De Bruyne, M.; De Rycke, R.; et al. Vessel-specific reintroduction of CINNAMOYL-COA REDUCTASE1 (CCR1) in dwarfed ccr1 mutants restores vessel and xylary fiber integrity and increases biomass. Plant Physiol. 2018, 176, 611–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escamilla-Treviño, L.L.; Shen, H.; Uppalapati, S.R.; Ray, T.; Tang, Y.; Hernandez, T.; Yin, Y.; Xu, Y.; Dixon, R.A. Switchgrass (Panicum virgatum) possesses a divergent family of cinnamoyl CoA reductases with distinct biochemical properties. New Phytol. 2010, 185, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, R.; Sun, Y.H.; Li, Q.; Heber, S.; Sederoff, R.; Chiang, V.L. Towards a systems approach for lignin biosynthesis in Populus trichocarpa: Transcript abundance and specificity of the monolignols biosynthetic genes. Plant Cell Physiol. 2010, 51, 144–163. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Jackson, L.; Shadle, G.; Nakashima, J.; Temple, S.; Chen, F.; Dixon, R.A. Distinct cinnamoyl CoA reductases involved in parallel routes to lignin in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2010, 107, 17803–17808. [Google Scholar] [CrossRef] [Green Version]
- Borah, P.; Khurana, J.P. The OsFBK1 E3 ligase subunit affects anther and root secondary cell wall thickenings by mediating turnover of a cinnamoyl-CoA reductase. Plant Physiol. 2018, 176, 2148–2165. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, Y.; Zhang, X.; Zhang, R.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef]
- The Protein Database from NCBI. Available online: https://ncbi.nlm.nih.gov/protein (accessed on 30 April 2021).
- Chao, N.; Jiang, W.T.; Wang, X.C.; Jiang, X.N.; Gai, Y. Novel motif is capable of determining CCR and CCR-like proteins based on the divergence of CCRs in plants. Tree Physiol. 2019, 39, 2019–2026. [Google Scholar] [CrossRef]
- Xia, H.; Liu, X.; Lin, Z.; Zhang, X.; Wu, M.; Wang, T.; Deng, H.; Wang, J.; Lin, L.; Deng, Q.; et al. Genome-wide identification of MYB transcription factors and screening of members involved in stress response in Actinidia. Int. J. Mol. Sci. 2022, 23, 2323. [Google Scholar] [CrossRef]
- Beckers, B.; De Beek, M.O.; Weyens, N.; Van Acker, R.; Van Montagu, M.; Boerjan, W.; Vangronsveld, J. Lignin engineering in field-grown polar trees affects the endosphere bacterial microbiome. Proc. Natl. Acad. Sci. USA 2016, 113, 2312–2317. [Google Scholar] [CrossRef] [Green Version]
- Halpin, C. Lignin engineering to improve saccharification and digestibility in grasses. Curr. Opin. Biotechnol. 2019, 56, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, J.H.; Chen, F.; Torres-Jerez, I.; Tang, Y.; Wang, M.; Du, Q.; Cheng, X.; Wen, J.; Dixon, R. Transcriptome analysis of secondary cell wall development in Medicago truncatula. BMC Genomics 2016, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.A.; Collins, R.E.; Anterola, A.M.; Cochrane, F.C.; Davin, L.B.; Lewis, N.G. An in silico assessment of gene function and organization of the phenylpropanoid pathway metabolic networks in Arabidopsis thaliana and limitations thereof. Phytochemistry 2003, 64, 1097–1112. [Google Scholar] [CrossRef]
- Chao, N.; Li, N.; Qi, Q.; Li, S.; Lv, S.; Jiang, X.N.; Gai, Y. Characterization of the cinnamol-CoA reductase (CCR) gene family in Populus tomentosa reveals the enzymeatic active sites and evolution of CCR. Planta 2017, 245, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Zhao, Y.; Wang, H.; Li, G.; Cheng, X.; Jin, Q.; Cai, Y. Transcriptomic analysis of early fruit development in Chinese white pear (Pyrus bretschneideri Rehd.) and functional identification of PbCCR1 in lignin biosynthesis. BMC Plant Biol. 2019, 19, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Niu, Y.; Zheng, Y. Multiple functions of MYB transcription factors in abiotic stress responses. Int. J. Mol. Sci. 2021, 22, 6125. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zhang, C.; Guo, X.; Li, H.; Lu, H. MYB transcription factors and its regulation in secondary cell wall formation and lignin biosynthesis during xylem development. Int. J. Mol. Sci. 2021, 22, 3560. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wang, D.; Gao, Y.; Li, C.; Jiang, H.; Zhu, X.; Zhang, H. Changes of lignin biosynthesis in tobacco leaves during maturation. Funct. Plant Biol. 2021, 48, 624–633. [Google Scholar] [CrossRef]
- Wang, Y.X.; Teng, R.M.; Wang, W.L.; Wang, Y.; Shen, W.; Zhuang, J. Identification of genes revealed differential expression profiles and lignin accumulation during leaf and stem development in tea plant (Camellia sinensis (L.) O. Kuntze). Protoplasma 2019, 256, 359–370. [Google Scholar] [CrossRef]
- Yoon, J.; Choi, H.; An, G. Roles of lignin biosynthesis and regulatory genes in plant development. J. Integr. Plant Biol. 2015, 57, 902–912. [Google Scholar] [CrossRef]
- Rao, X.; Chen, X.; Shen, H.; Ma, Q.; Li, G.; Tang, Y.; Pena, M.; York, W.; Frazier, T.P.; Lenaghan, S.; et al. Gene regulatory networks for lignin biosynthesis in switchgrass (Panicum virgatum). Plant Biotechnol. J. 2019, 17, 580–593. [Google Scholar] [CrossRef]
- Cheng, X.; Li, M.; Li, D.; Zhang, J.; Jin, Q.; Sheng, L.; Cai, Y.; Lin, Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit. Biol. Open 2017, 6, 1602–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Wang, L.; Zhang, Y.; Huang, D. Identification of early response genes to salt stress in roots of melon (Cucumis melo L.) seedlings. Mol. Biol. Rep. 2013, 40, 2915–2926. [Google Scholar] [CrossRef] [PubMed]
- Hori, C.; Yu, X.; Mortimer, J.C.; Sano, R.; Matsumoto, T.; Kikuchi, J.; Demura, T.; Ohtani, M. Impact of abiotic stress on the regulation of cell wall biosynthesis in Populus trichocarpa. Plant Biotech. 2020, 37, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wu, J.; Lin, L.; Li, P.; Wang, Y.; Li, J.; Sun, Q.; Liang, J.; Wang, Y. Overexpression of cinnamoyl-CoA reductase 2 in Brassica napus increases resistance to sclerotinia sclerotiorum by affecting lignin biosynthesis. Front. Plant Sci. 2021, 12, 732733. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.W.; Choi, S.; Jin, X.; Jung, S.E.; Choi, J.W.; Seo, J.S.; Kim, J.K. Transcriptional activation of rice CINNAMOYL-CoA REDUCTASE 10 by OsNAC5, contributes to drought tolerance by modulating lignin accumulation in roots. Plant Biotechnol. J. 2022, 20, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Yassin, N.B.M.; Park, J.S.; Choi, A.; Herr, J.; Carlson, J.E. Comparative and phylogenimic analyses of cinnamoyl-CoA reductase and cinnamoyl-CoA-reductase-like gene family in land plants. Plant Sci. 2011, 181, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Xio, Y.; Lv, Z.; Tan, H.; Chen, R.; Li, Q.; Chen, J.; Wang, Y.; Yin, J.; Zhang, L.; et al. AP2/ERF transcription factor, Ii049, positively regulates lignin biosynthesis in Isatic indigotica through activating salicylic acid signaling and lignin/lignin pathway genes. Front. Plant Sci. 2017, 8, 1361. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cheng, C.; Zhang, X.; Zhou, S.; Wang, C.; Ma, C.; Yang, S. PpNAC187 enhances lignin synthesis in “Whangkeumbae” pear (Pyrus pyrifolia) “Hard-End” Fruit. Molecules 2019, 24, 4338. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, N.M.; Marina, M.; Nardi, C.F.; Civello, P.M.; Martínez, G.A. Novel insights of ethylene role in strawberry cell wall metabolism. Plant Sci. 2016, 252, 1–11. [Google Scholar] [CrossRef]
- Lan, Y.; Chai, Y.; Xing, C.; Wu, K.; Wang, L.; Cai, M. Nitric oxide reduces the aluminum-binding capacity in rice root tips by regulating the cell wall composition and enhancing antioxidant enzymes. Ecotoxicol. Environ. Saf. 2021, 208, 111499. [Google Scholar] [CrossRef]
- Lacombe, E.; Hawkins, S.; Doorsselaere, J.; Piquemal, J.; Goffner, D.; Poeydomenge, O.; Boudet, A.M.; Grima-Pettenati, J. Cinnamoyl CoA reductase, the first committed enzyme of the lignin branch biosynthetic pathway: Cloning, expression and phylogenetic relationships. Plant J. 1997, 11, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Baltas, M.; Lapeyre, C.; Bedos-Belval, F.; Maturano, M.; Saint-Aguet, P.; Roussel, L.; Duran, H.; Grima-Pettenati, J. Kinetic and inhibition studies of cinnamoyl-CoA reductase 1 from Arabidopsis thaliana. Plant Physiol. Biochem. 2005, 43, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhou, R.; Louie, G.V.; Mühlemann, J.K.; Bomati, E.K.; Bowman, M.E.; Dudareva, N.; Dixon, R.A.; Noel, J.P.; Wang, X. Structural studies of cinnamoyl-CoA reductase and cinnamyl-alcohol dehydrogenase, key enzymes of monolignol biosynthesis. Plant Cell. 2014, 26, 3709–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bart, R.S.; Chern, M.; Vega-Sánchez, M.E.; Canlas, P.; Ronald, P.C. Rice Snl6, a cinnamoyl-CoA reductase-like gene family member, is required for NH1-mediated immunity to Xanthomonas oryzae pv. oryzae. PLoS Genet. 2010, 6, e1001123. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.; Yao, P.; Pan, J.; Dai, C.; Cao, H.; Chen, Z.; Zhang, S.; Xu, S.; Shen, W. Transcriptome analysis reveals insight into molecular hydrogen-induced cadmium tolerance in alfalfa: The prominent role of sulfur and (homo)glutathione metabolism. BMC Plant Biol. 2020, 20, 58. [Google Scholar] [CrossRef]
- Hdira, S.; Haddoudi, L.; Hanana, M.; Romero, I.; Mahjoub, A.; Ben Jouira, H.; Ludidi, N.; Sanchez-Ballesta, M.T.; Abdelly, C.; Badri, M. Morpho-Physiological, Biochemical, and Genetic Responses to Salinity in Medicago truncatula. Plants 2021, 10, 808. [Google Scholar] [CrossRef]
- Chen, T.; Yang, Q.; Zhang, X.; Ding, W.; Gruber, M. An alfalfa (Medicago sativa L.) ethylene response factor gene, MsERF11, enhances salt tolerance in transgenic Arabidopsis. Plant Cell Rep. 2012, 31, 1737–1746. [Google Scholar] [CrossRef]
- Chen, M.; Cui, W.; Zhu, K.; Xie, Y.; Zhang, C.; Shen, W. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, W.; Duan, X.; Dai, C.; Zhang, Y.; Cui, W.; Wang, R.; Shen, W. Hydrogen-rich water-alleviated ultraviolet-B-triggered oxidative damage is partially associated with the manipulation of the metabolism of (iso)flavonoids and antioxidant defence in Medicago sativa. Funct. Plant Biol. 2015, 42, 1141–1157. [Google Scholar] [CrossRef]
- Dražić, G.; Mihailović, N.; Lojić, M. Cadmium accumulation in Medicago sativa seedlings treated with salicylic acid. Biol. Plant. 2006, 50, 239–244. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Abscisic acid increases anaerobic tolerance in alfalfa seedlings. Physiol. Plantarum 1999, 107, 395–398. [Google Scholar] [CrossRef]
- The Alfalfa (M. sativa L.) Genomic Data Base. Available online: https://figshare.com/projects/whole_genome_sequencing_and_assembly_of_Medicago_sativa/66380 (accessed on 20 March 2021).
- The ExPASy Compute pI/MW Tool. Available online: https://www.expasy.org/resources/compute-pi-mw (accessed on 26 April 2021).
- The Multiple Em for Motif Elicitation Tool. Available online: http://meme-suite.org (accessed on 26 April 2021).
- Hossain, M.S.; Ahmed, B.; Ullah, M.W.; Haque, M.S.; Islam, M.S. Genome-wide identification and characterization of expansin genes in jute. Trop. Plant Biol. 2022, 15, 40–54. [Google Scholar] [CrossRef]
- Chakraborty, A.; Viswanath, A.; Malipatil, R.; Semalaiyappan, J.; Shah, P.; Ronanki, S.; Rathore, A.; Singh, S.P.; Govindaraj, M.; Tonapi, V.A.; et al. Identification of candidate genes regulating drought tolerance in Pearl Millet. Int. J. Mol. Sci. 2022, 23, 6907. [Google Scholar] [CrossRef] [PubMed]
- The GSDS Tool. Available online: http://gsds.cbi.pku.edu.cn/ (accessed on 26 April 2021).
- The MEGA-X Tool. Available online: https://www.megasoftware.net/ (accessed on 30 April 2021).
- The iTOL Software. Available online: https://itol.embl.de/ (accessed on 16 May 2022).
- The BioEdit Software. Available online: https://www.nucleics.com/DNA_sequencing_support/Trace_viewer_reviews/BioEdit (accessed on 26 April 2021).
- The PlantCARE Tool. Available online: https//bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 11 May 2021).
- The SWISS-MODEL Tool. Available online: https://swissmodel.expasy.org/ (accessed on 29 April 2021).
- The PyMOL Tool. Available online: https://pymol.org/2/ (accessed on 12 May 2022).
- The TBtools. Available online: https://github.com/CJ-Chen/TBtools/releases (accessed on 12 May 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbols | Feruloyl-CoA | p-Coumaroyl-CoA | Caffeoyl-CoA | Sinapoyl-CoA | ||||
|---|---|---|---|---|---|---|---|---|
| Km | Kcat/Km | Km | Kcat/Km | Km | Kcat/Km | Km | Kcat/Km | |
| MsCCR1 | 45.20 ± 11.42 ab | 2.09 ± 0.18 B | 24.22 ± 4.51 bc | 2.58 ± 0.11 B | ND | ND | 10.57 ± 1.25 c | 5.72 ± 0.36 A |
| MsCCR-like1 | ND | ND | 44.14 ± 7.12 ab | 1.09 ± 0.19 C | 48.16 ± 1.75 a | 1.09 ± 0.11 C | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, W.; Zhuang, Z.; Jiang, P.; Pan, J.; Zhao, G.; Xu, S.; Shen, W. Characterization, Expression Profiling, and Biochemical Analyses of the Cinnamoyl-CoA Reductase Gene Family for Lignin Synthesis in Alfalfa Plants. Int. J. Mol. Sci. 2022, 23, 7762. https://doi.org/10.3390/ijms23147762
Cui W, Zhuang Z, Jiang P, Pan J, Zhao G, Xu S, Shen W. Characterization, Expression Profiling, and Biochemical Analyses of the Cinnamoyl-CoA Reductase Gene Family for Lignin Synthesis in Alfalfa Plants. International Journal of Molecular Sciences. 2022; 23(14):7762. https://doi.org/10.3390/ijms23147762
Chicago/Turabian StyleCui, Weiti, Zihan Zhuang, Peihao Jiang, Jincheng Pan, Gan Zhao, Sheng Xu, and Wenbiao Shen. 2022. "Characterization, Expression Profiling, and Biochemical Analyses of the Cinnamoyl-CoA Reductase Gene Family for Lignin Synthesis in Alfalfa Plants" International Journal of Molecular Sciences 23, no. 14: 7762. https://doi.org/10.3390/ijms23147762