
Impact of the Exposome on the Epigenome in Inflammatory Bowel Disease Patients and Animal Models

 , , , ,
, , , ,
Abstract
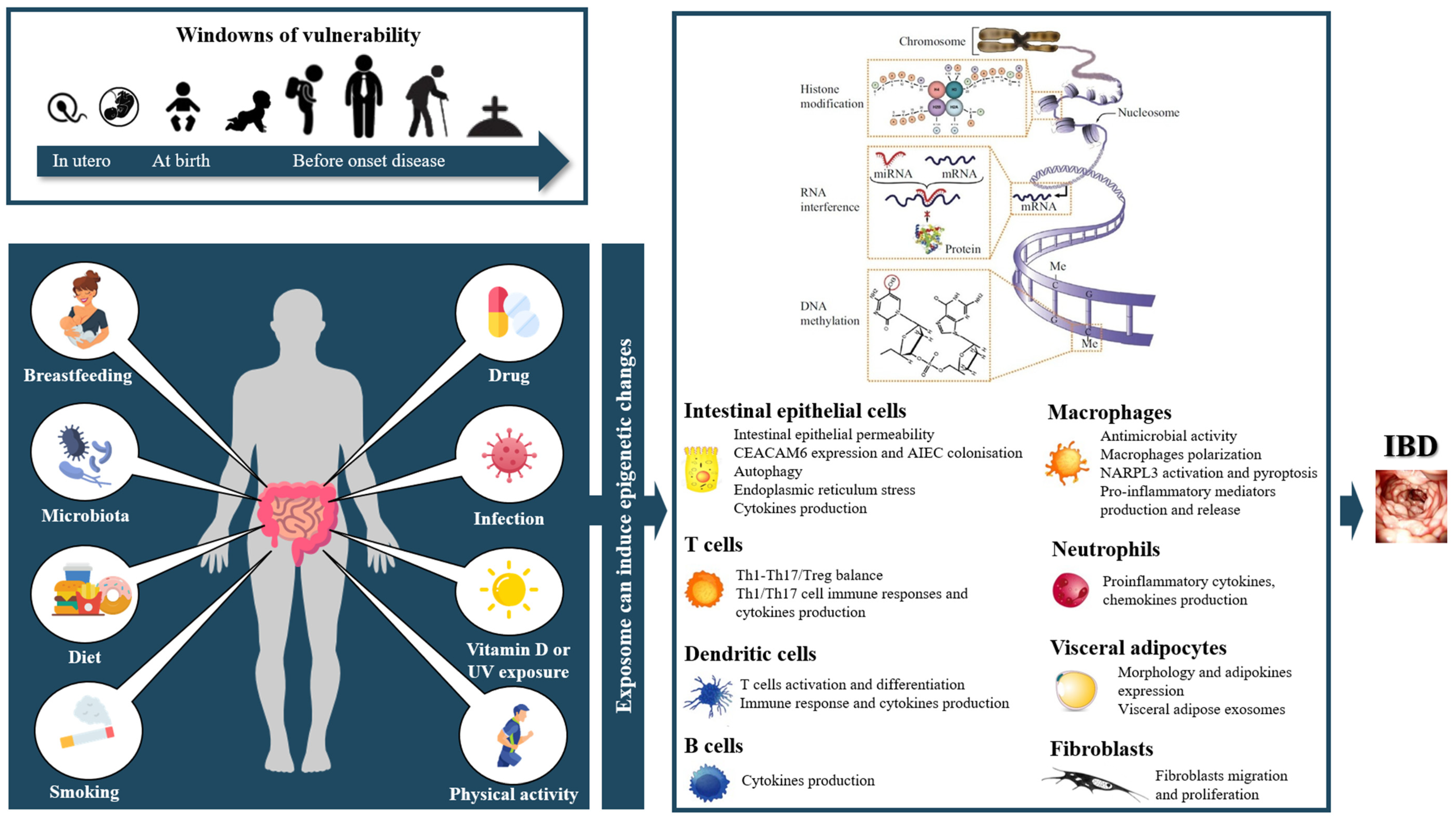
:1. Introduction
2. Epigenetics in IBD
3. Methods
4. Results
4.1. Parental Exposition
4.2. Microbiota

{kind=link}
| Germ | Activity | Epigenetic Mechanism | Tissue/Cells | Mechanism | Model | Author |
|---|---|---|---|---|---|---|
| Commensal bacteria | ||||||
| Commensal bacteria | Anti-inflammatory activity | miR-10a | DCs | Negatively regulates host miR-10a expression, which contribute to the intestinal homeostasis maintenance by targeting IL-12/IL-23p40 expression | C57BL/6 (B6) mice | Xue X, et al. (2011) [125] |
| Commensal flora | Proinflammatory activity | miR-107 | DCs and macrophages | Downregulates miR-107 expression, known to represses the expression of IL-23p19, thereby favouring IL-23 expression | IECs, lamina propria CD11c+ myeloid cells including dendritic cells and macrophages, and T cells; DSS-induced colitis in mice | Xue X, et al. (2014) [126] |
| Commensal bacteria | Anti-inflammatory activity | miR-10a | DCs | Inhibits human DCs miR-10a expression, which downregulates mucosal inflammatory response through inhibition of IL-12/IL-23p40 and NOD2 expression, and blockade of Th1/Th17 cell immune responses | Human monocyte-derived dendritic cells | Wu W, et al. (2015) [127] |
| Commensal microbiome-dependent (Bacteroides acidifaciens and Lactobacillus johnsonii | Anti-inflammatory activity | miR-21-5p | IECs | Commensal microbiome-dependent miR-21-5p expression in IECs regulates intestinal epithelial permeability via ADP Ribosylation Factor 4 (ARF4) | HT-29 and Caco-2 cells | Nakata K., et al. (2017) [128] |
| Cluster(s) | ||||||
| ↓ of Bacteroidetes and ↑ of protective Firmicutes and Clostridia | Anti-inflammatory activity | miR-21 | Colonic mucosae | Leads to miR-21 reduction, known to influence the pathogenesis of intestinal inflammation by causing propagation of a disrupted gut microbiota | WT and miR-21−/− mice | Johnston DGW, et al. (2018) [129] |
| Cluster enriched in Bacteroides fragilis | - | DNA methylation | Intestinal mucosa | Induces 33 and 19 significantly hyper-methylated or hypomethylated sites, including hyper-methylated signals in the gene body of Notch Receptor 4 (NOTCH4) | 50 CD; 80 UC; 31 controls | Ryan FJ, et al. (2020) [130] |
| Cluster enriched in Escherichia/Shigella/Klebsiella and Ruminococcus gnavus | Proinflammatory activity | DNA methylation | Intestinal mucosa | Larger number of differentially methylated CpG sites (131 hyper- and 475 hypomethylated), including hypomethylation in CCDC88B (recently correlated with risk of CD) and Transporter 2 (TAP2), involved in genetic heterogeneity of CD | ||
| Cluster enriched in B. vulgatus | - | DNA methylation | Intestinal mucosa | Induces 23 hyper- and 18 hypomethylated sites, significant hyper-methylation was observed in the gene body of DNA Damage Regulated Autophagy Modulator 1 (DRAM1) | ||
| Specific germ | ||||||
| Adherent-invasive Escherichia coli (AIEC) | Proinflammatory activity | miR-30c and miR-130a | IECs | Upregulates levels of miR-30c and miR-130a in IECs (by activating NF-κB), reducing the levels of ATG5 and ATG16L1 and inhibiting autophagy, leading to increased numbers of intracellular AIEC and an increased inflammatory response | Cultured IECs and mouse enterocytes | Nguyen HT, et al. (2014) [115] |
| AIEC | Proinflammatory activity | let-7b | IECs | Instigates excessive mucosal immune response against gut microbiota via miR let-7b/TLR4 signaling pathway | WT and IL-10 KO mice; T84 cells | Guo Z, et al. (2018) [117] |
| AIEC | Proinflammatory activity | miR-30c and miR-130a | IECs | AIEC-infected IECs secretes exosomes that can transfer specific miRs (miR-30c and miR-130a) to recipient IECs, inhibiting autophagy-mediated clearance of intracellular AIEC | T84 cells | Larabi A, et al. (2020) [116] |
| Mycobacterium avium subspecies paratuberculosis (MAP) | Proinflammatory activity | miR-21 | Macrophages | MAP upregulates miR-21 in macrophages, a change that results in diminished macrophages clearance ability and favours pathogens survival within the cells | THP-1 cells | Mostoufi-Afshar S, et al. (2018) [119] |
| Lactobacillus rhamnosus GG | Anti-inflammatory activity | miR-146a and miR-155 | DCs | Induces a significant downregulation of miR-146a expression, a negative regulator of immune response, and upupregulation of on miR-155 | Cultured DCs | Giahi L., et al. (2012) |
| Lactobacillus acidophilus | Anti-inflammatory activity | miRs | Colonic mucosae | L. acidophilus induce miRs expression | DSS-induced colitis in mice | Kim WK, et al. (2021) [131] |
| Faecalibacterium prausnitzii | Anti-inflammatory activity | HDAC1 inhibition | T cells | Inhibits HDAC1, promotes Foxp3 and blocks the IL-6/STAT3/IL-17 downstream pathway contributing to the maintain of Th17/Treg balance | IBD patients (n = 9) and healthy control (n = 6); DSS-induced colitis in mice | Zhou L, et al. (2018) [132] |
| Faecalibacterium prausnitzii | Anti-inflammatory activity | HDAC3 inhibition | T cells | Produces butyrate to decrease Th17 differentiation and attenuate colitis through inhibiting HDAC3 and c-Myc-related metabolism in T cells | IBD patients; TNBS-induced colitis in mice | Zhang M, et al. (2019) [133] |
| Trichinella spiralis | Anti-inflammatory activity | miRs | T cells | Extra-vesicles-derived miR are involved in the regulation of the host immune response, including inflammation, including increase of Th2 and Treg cells | TNBS-induced colitis in mice | Yang Y, et al. (2020) [134] |
| Enterotoxigenic Bacteroides fragilis (ETBF) | Proinflammatory activity | miR-149-3p | T cells | Downregulates miR-149-3p, which play a role in modulation of T-helper type 17 cell differentiation (with increased number of T-helper type 17 cell contributing to intestinal inflammation) | ETBF cells | Cao Y, et al. (2021) [135] |
| Bacterial component | ||||||
| Roseburia intestinalis-derived flagellin | Anti-inflammatory activity | lncRNA | IECs | Flagellin induces p38-stat1 activation, activated HIF1A-AS2 promotor, induced HIF1A-AS2 (a lncRNA) expression in gut epithelium in a dose- and time-dependent manner. HIF1A-AS2 inactivates NF-κB/Jnk pathway and thus inhibits inflammatory responses | DSS/Flagellin-challenged mice; Caco-2 cells | Quan Y, et al. (2018) [124] |
| Roseburia intestinalis-derived flagellin | Anti-inflammatory activity | miR-223-3p | Macrophages | Flagellin inhibited activation of the NLRP3 inflammasome and pyroptosis via miR-223-3p/NLRP3 signaling in macrophages | DSS-induced colitis model in C57Bl/6 mice and the LPS/ATP-induced THP-1 macrophages | Wu X, et al. (2020) [123] |
| LPS | Proinflammatory activity | H3K4me1, H3K4me3, and H3K27ac histone | Macrophages | Increases H3K4me1, H3K4me3, and H3K27ac histone marks, particularly in genes associated with an inflammatory response such as IL-12a and IL-18 | IL-10-deficient (Il10(−/−)) mice | Simon JM, et al. (2016) [136] |
| LPS and flagellin | Anti-inflammatory activity | miR-146 | IECs | Stimulate miR-146a overexpression in IECs, induces immune tolerance, inhibiting cytokine production (MCP-1 and GROα/IL-8) | TNBS and DSS-induced colitis in mice | Anzola A, et al. (2018) [137] |
| LPS | Proinflammatory activity | lncRNA H19 | IECs | Increases levels of H19 lncRNA in epithelial cells in the intestine. H19 lncRNA bound to p53 and miR (miR-34a and let-7) that inhibit cell proliferation (alters regeneration of the epithelium) | Intestinal tissues of UC patients and mice | Geng H, et al. (2018) [138] |
| LPS | Proinflammatory activity | miR-19b | IECs | LPS significantly induces cell inflammatory injury, downregulated miR-19b expression and activates NF-κB and PI3K/AKT pathway | Caco2 cells | Qiao CX, et al. (2018) [120] |
| LPS | Proinflammatory activity | lncRNA | Monocytes/macrophages | LPS promotes a downregulation of the lncRNA growth arrest-specific transcript 5 (GAS5), could mediate tissue damage by modulating the expression of matrix metalloproteinases | IBD patients (n = 25) | Lucafò M, et al. (2019) [139] |
| LPS | Proinflammatory activity | miR-215 | Fibroblasts | LPS upregulates the expression of miR-215, increases oxidative stress in LPS-treated intestinal fibroblast by downregulating GDF11 (Growth differentiation factor 11) expression and activating the TLR4/NF-κB and JNK/p38 signaling pathways | CCD-18Co cells | Sun B, et al. (2020) [122] |
| LPS | Proinflammatory activity | miR-506 and DNMT1 modification | IECs | LPS inhibits miR-506, leading to reduced expression of anion exchange protein 2 and inositol-1,4,5-trisphosphate-receptor but was accompanied by a substantial increase in DNMT1 and SPHK1 (sphingosine kinase 1) expression. The enhanced levels of kinase SPHK1 resulte in upregulation of bioactive sphingosine-1-phosphate (S1P) which led to further activation of S1P-dependent signaling pathways. The net effect of these responses is severe inflammation | Patients with PSC, PSC with concurrent UC (PSC + UC), UC alone, and healthy controls (n = 10 each); Caco2 cells | Kempinska-Podhorodecka A, et al. (2021) [140] |
| LPS | Proinflammatory activity | miR-497 | Macrophages | Reduces miR-497, promotes the activation of NF-κB pathway and the release of cytokines | IBD patients, mice with colitis and LPS-treated RAW264.7 cells | Zhang M, et al. (2021) [121] |
4.3. Gut Microbiota-Derived Metabolites
4.4. Diet
| Food | Activity | Epigenetic Mechanism | Tissue/Cells | Mechanism | Model | Author |
|---|---|---|---|---|---|---|
| Diet | ||||||
| Western diet | Proinflammatory activity | miR-143, miR-145A, miR-148a, miR-152 | IECs | Leads to a decrease in miR-143/145a, miR-148a and miR-152 in colonocytes with a consequent increase in ADAM17 expression protein (these miRs regulating ADAM17) and aggravates colitis. | DSS-induced colitis in mice | Dougherty U, et al. (2021) [164] |
| High fat diet | Proinflammatory activity | miR-155 | Visceral adipocytes | High fat diet changes the miR profile (among which miR-155) of the visceral adipose exosomes, switching the exosomes from anti-inflammatory to a proinflammatory phenotype. | Macrophages | Wei M, et al. (2020) [193] |
| High fat diet rich in n-6 linoleic acid | Proinflammatory activity | DNA methylation | Colonic mucosae | Epigenetically modifies farnesoid-X-receptor (FXR), leading to the activation of downstream factors that participate in bile acid homeostasis and epigenetically activates prostaglandin-endoperoxide synthase-2 (Ptsg-2) coupled accumulation of c-JUN and proliferative cyclin D1(Ccnd1) and increase the risk of inflammation | C57BL/6J mice; Human colonic foetal cells | Romagnolo DF, et al. (2019) [194] |
| Methyl-deficient diet | Proinflammatory activity | Sirtuin 1 | IECs | Reduces sirtuin 1 (SIRT1) expression level and promotes greater acetylation of (heat shock factor protein 1) HSF1, in relation with a dramatic decrease of chaperones (binding immunoglobulin protein (BIP), heat shock protein (HSP)27 and HSP90) | DSS-induced colitis in mice; Caco-2 cells | Melhem H, et al. (2016) [165] |
| Low-methyl diet | Proinflammatory activity | DNA methylation | IECs | Low-methyl diet-dependent HRE demethylation led to abnormal gut expression of CEACAM6 (carcinoembryonic antigen-related cell adhesion molecule 6), favouring AIEC colonisation and subsequent inflammation | Transgenic mice; Caco-2, T-84 and sh-HIF1-α-T-84 cells | Denizot J, et al. (2015) [166] |
| Methyl-donnor supplemented diet (folate, B12 vitamin) | Anti-inflammatory activity | DNA methylation | IECs | Methyl-donor supplemented diet contributes to hypermethylation of CEACAM6 promoter in IECs, associated with a significant decrease in CEACAM6 expression contributing to less adherence of AIEC bacteria to the enterocytes | CEABAC10 mice | Gimier E, et al. (2020) [195] |
| Isolated food | ||||||
| Cow’s milk (commercial) | Anti-inflammatory activity | miR-21, miR-29b and miR-125b | Colonic mucosae | Extracellular vesicles (EVs) concentrated from commercial cow’s milk downregulates miR-21, miR-29b and miR-125b. MiR-125b was associated with a higher expression of the NF-κB inhibitor TNFAIP3 (A20) | DSS-induced colitis in mice | Benmoussa A, et al. (2019) [178] |
| Human milk derived exosomes | Anti-inflammatory activity | miR-320, miR-375, and Let-7 and DNMT1 and DNMT3 | Colonic mucosae | MiR highly express in milk, such as miR-320, 375, and Let-7, were found to be more abundant in the colon of milk derived exosomes-treated mice compared with untreated mice. These miR downregulate their target genes, mainly DNA methyltransferase 1 (DNMT1) and DNMT3 | DSS-induced colitis in mice; PBMC | Reif S, et al. (2020) [179] |
| Dietary depletion of milk exosomes and their microRNA cargos | Proinflammatory activity | miR-200a-3p | Cecum mucosae | Elicits a depletion of miR-200a-3p and elevated intestinal inflammation and chemokine (C-X-C Motif) ligand 9 expression | Mdr1a−/− mice | Wu D, et al. (2019) [180] |
| Saccharin sodium, Stevioside, and Sucralose (three common sweeteners) | Anti-inflammatory activity | miR-15b | IECs | Upregulate the expression of E-cadherin through the miR-15b/RECK/MMP-9 axis to improve intestinal barrier integrity. Saccharin exerts the most pronounced effect, followed by Stevioside and Sucralose | DSS-induced colitis in mice | Zhang X, et al. (2022) [181] |
| Galacto-oligosaccharides (GOS) | Anti-inflammatory activity | miR-19 | IECs | GOS increases of cell viability, the decrease of apoptosis, as well as the suppressed release of TNF-α, IFN-γ and IL-1β by upregulating miR-19b | Human colon epithelial FHC cells; Helicobacter hepaticus induced colitis in rats | Sun J, et al. (2019) [182] |
| Cinnamaldehyde (a major active compound from cinnamon) | Anti-inflammatory activity | miR-21 and miR-155 | Macrophages | Cinnamaldehyde inhibits NLRP3 inflammasome activation as well as miR-21 and miR-155 level in colon tissues and macrophage. The decrease in miR-21 and miR-155 suppresses levels of IL-1β and IL-6; | DSS-induced colitis in mice; macrophage cell line RAW264.7 and human monocytes U937 | Qu S, et al. (2018) [185] |
| Cinnamaldehyde | Anti-inflammatory activity | lncRNAs H19 | T cells | Cinnamaldehyde inhibits Th17 cell differentiation by regulating the expression of lncRNA H19 | DSS-induced colitis in mice and naïve CD4+ T cells | Qu SL, et al. (2021) [101] |
| Limonin (a triterpenoid extracted from citrus) | Anti-inflammatory activity | miR-124 | IECs | Downregulates p-STAT3/miR-214 signaling pathway and represses the productions of proinflammatory cytokines (such as TNF-α and IL-6) | DSS-induced colitis in mice; cultured normal colonic epithelial cells | Liu S, et al. (2019) [186] |
| Edible ginger | Anti-inflammatory activity | Contained around 125 miRNAs | IECs | Increases the survival and proliferation of IECs, reduces the proinflammatory cytokines (such as TNF-α, IL-6 and IL-1β), and increases the anti-inflammatory cytokines (including IL-10 and IL-22) in colitis | DSS-induced colitis in mice | Zhang M, et al. (2016) [187] |
| Ginsenoside Rh2 (active ingredient of ginseng) | Anti-inflammatory activity | miR-124 | IECs | Inhibits IL-6-induced STAT3 phosphorylation and miR-214 expression (which is an inflammatory effector molecule acting through NF-κB-IL6 pathway) | DSS-induced colitis in mice; cultured normal colonic epithelial cells | Chen X, et al. (2021) [188] |
| Black raspberries (BRBs) | Anti-inflammatory activity | Demethylation the promoter of dkk3; correction of promoter hypermethylation of suppressor genes | Colonic mucosae | BRBs exert their anti-inflammatory effects is through decreasing NF-κB p65 expression leading to decrease of DNMT3B expression (but also histone deacetylases 1 and 2 (HDAC1 and HDAC2) and methyl-binding domain 2 or MBD2), which in turn reverse aberrant DNA methylation of tumor suppressor genes, e.g., dkk2, dkk3, in the Wnt pathway, resulting in their enhanced mRNA expression locally in colon and systematically in spleen and bone marrow and thus in decreased translocation of β-catenin to the nucleus prohibiting the activation of the pathway | DSS-induced colitis in mice; splenocytes and bone marrow cells | Wang LS, et al. (2013) [189] |
| Black raspberries | Anti-inflammatory activity | Demethylation | Colonic mucosae | BRBs decreas the methylation of wif1, sox17, and qki gene promoters and thus increase their mRNA expression (contributing to Wnt signaling) | Interleukin-10 knockout mice | Wang LS, et al. (2013) [190] |
| Mastiha | Anti-inflammatory activity | miR-155 | T cells | Plays a role in circulating levels of miR-155, a critical player in T helper-17 (Th17) differentiation and function | UC patients (n = 35) | Amerikanou C, et al. (2021) [196] |
| Isoliquiritigenin | Anti-inflammatory activity | HDACs inhibition | IECs | Suppresses acetylated HMGB1 release via the induction of HDAC activity, which is one of the critical mediators of inflammation, which is actively secreted from inflammatory cytokine-stimulated immune or non-immune cells | HT-29 cells | Chi JH, et al. (2017) [197] |
| Chronic ethanol exposure | Proinflammatory activity | miR-122a | IECs | Increases the intestinal miR-122a expression, which decreased occludin (OCLN) expression leading to increased intestinal permeability | HT-29 cells | Chen Y, et al. (2013) [191] |
| Chronic alcohol feeding (but not acute alcohol binge) | Proinflammatory activity | miR-155 | Intestinal tissue | Increases miR-155 in the small bowel, which is a modulator of cytokine and T-cell immune response in the gut, leading to intestinal TNFα, and NF-κB activation | WT-mice | Lippai D, et al. (2014) [192] |
| Polyphenol | ||||||
| Polyphenolic red wine extract | Anti-inflammatory activity | miR-126 | Fibroblasts | Polyphenolic red wine extract downregulates miR-126, leading to downregulation of NF-kB, ICAM-1, VCAM-1, and PECAM-1 | CCD-18Co myofibroblasts cells | Angel-Morales G, et al. (2012) [198] |
| Polyphenolic extracts from cowpea (Vigna unguiculata) | Anti-inflammatory activity | miR-126 | Fibroblasts | Cowpea may exert their anti-inflammatory activities at least in part through induction of miR-126 that then downregulate VCAM-1 mRNA and protein expressions | CCD-18Co myofibroblasts cells | Ojwang LO, et al. (2015) [199] |
| Mango (Mangifera indica L.) polyphenolics | Anti-inflammatory activity | miR-126 | Fibroblasts | Mango polyphenols attenuates inflammatory response by modulating the PI3K/AKT/mTOR pathway at least in part through upregulation of miR-126 expression | CCD-18Co cells; DSS-induced colitis in rats | Kim H, et al. (2017) [200] |
| Baicalin (flavone) | Anti-inflammatory activity | miR-191a | IECs | Exerts a protective effect on IECs against TNF-α-induced injury, which is at least partly via inhibiting the expression of miR-191a, thus increasing ZO-1 expression | IEC-6 cells | Wang L, et al. (2017) [170] |
| Pomegranate (Punica granatum L.) polyphenolics | Anti-inflammatory activity | miR-145 | Myofibroblasts | Pomegranate polyphenols attenuate colitis by modulating the miR-145/p70S6K/HIF1α axis | DSS-induced colitis in rats; CCD-18Co colon-myofibroblastic cells | Kim H, et al. (2017) [201] |
| Alpinetin, a flavonoid compound extracted from the seeds of Alpinia katsumadai Hayata | Anti-inflammatory activity | miR-302 | T cells | Activates Aryl hydrocarbon receptor (AhR), promoting expression of miR-302, downregulating expression of DNA methyltransferase 1 (DNMT-1), reducing methylation level of Foxp3 promoter region, facilitating combination of CREB and promoter region of Foxp3, and upregulating the expression of Foxp3. Alpinetin ameliorates colitis in mice by recovering Th17/Treg balance. | DSS-induced colitis in mice | Lv Q, et al. (2018) [168] |
| Fortunellin, a citrus flavonoid | Anti-inflammatory activity | miR-374a | IECs | Fortunellin targets miR-374a, which is a negative regulator of PTEN, known to induce cell apoptosis | TNBS-induced colitis in rats | Xiong Y, et al. (2018) [169] |
| Quercetin (flavonoid) | Anti-inflammatory activity | miR-369-3p | DCs | Quercetin-induced miR-369-3p which reduce C/EBP-β, TNF-α, and IL-6 production | LPS-stimulated DCs | Galleggiante V, et al. (2019) [171] |
| Resveratrol (a natural plant product) | Anti-inflammatory activity | miR-31, Let7a, miR-132 | T cells | Resveratrol decreases the expression of several miRs (miR-31, Let7a, miR-132) that targets cytokines and transcription factors involved in anti-inflammatory T cell responses (Foxp3 and TGF-β). MiR-31 regulates the expression of Foxp3 with increase of CD4+ Foxp3+ regulatory T cells (Tregs) | TNBS-induced colitis in mice | Alrafas HR, et al. (2020) [176] |
| Resveratrol (an anti-oxidant) | Anti-inflammatory activity | HDACs inhibition | T cells | Inhibits HDACs, increases anti-inflammatory CD4+ FOXP3+ (Tregs) and CD4+ IL10+ cells, and decreases proinflammatory Th1 and Th17 cells | AOM and DSS-induced colitis in mice | Alrafas HR, et al. (2020) [175] |
| Chlorogenic acid (found in the coffee) | Anti-inflammatory activity | miR-155 | Macrophages | Downregulates miR-155 expression, inactivates the NF-κB/NLRP3 inflammasome pathway in macrophages and prevent colitis | DSS-induced colitis in mice; LPS/ATP-induced RAW264.7 cells | Zeng J, et al. (2020) [177] |
| Lonicerin (constituant of herb Lonicera japonica Thunb.) | Anti-inflammatory activity | H3K27me3 modification | Macrophages | Binds to enhancer of zeste homolog 2 (EZH2) histone methyltransferase, which mediate modification of H3K27me3 and promotes the expression of autophagy-related protein 5, which in turn leads to enhanced autophagy and accelerates autolysosome-mediated NLRP3 degradation | DSS-induced colitis in mice and isolated colonic macrophages and IECs; bone marrow-derived macrophages | Lv Q, et al. (2021) [174] |
| Pristimerin (Pris), which is a natural triterpenoid compound extracted from the Celastraceae plant | Anti-inflammatory activity | miR-155 | Colonic mucosae | Pris may reduce DSS-induced colitis in mice by inhibiting the expression of miR-155 | Blood and colon tissue of IBD patients; DSS-induced colitis in mice | Tian M, et al. (2021) [202] |
| Cardamonin is a naturally occurring chalcone (majorly from the Zingiberaceae family incluging a wide range of spices from India) | Anti-inflammatory activity | Modulation of miR expression | Macrophages | Cardamonin modulates miR expression, protects the mice from DSS-induced colitis, decreases the expression of iNOS, TNF-α, and IL-6, and inhibited NF-kB signaling which emphasizes the role of cardamonin as an anti-inflammatory molecule | RAW 264.7 Cells (monocyte/macrophage-like cells); DSS-induced colitis in mice | James S, et al. (2021) [172] |
| Berberine | Anti-inflammatory activity | miR-103a-3p | IECs | Represses Wnt/β-catenin pathway activation via modulating the miR-103a-3p/Bromodomain-containing protein 4 axis, thereby refraining pyroptosis and reducing the intestinal mucosal barrier defect induced via colitis | DSS-induced colitis in mice; Caco-2 cells and human NCM460 cells | Zhao X, et al. (2022) [173] |
4.5. Smoking
4.6. Drugs
4.7. Vitamin D
4.8. Physical Activity
5. Limitations to the Analysis of the Exposome Impact on the Epigenome in IBD
6. Conclusions and Challenge for the Future
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Danese, S.; Fiocchi, C. Etiopathogenesis of inflammatory bowel diseases. World J. Gastroenterol. 2006, 12, 4807–4812. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Wacholder, S.; Gail, M.H.; Peters, U.; Jacobs, K.B.; Chanock, S.J.; Chatterjee, N. Estimation of effect size distribution from genome-wide association studies and implications for future discoveries. Nat. Genet. 2010, 42, 570–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanahan, F. The gut microbiota-A clinical perspective on lessons learned. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 609–614. [Google Scholar] [CrossRef]
- Bernstein, C.N.; Shanahan, F. Disorders of a modern lifestyle: Reconciling the epidemiology of inflammatory bowel diseases. Gut 2008, 57, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Rubio, K.; Dobersch, S.; Barreto, G. Functional interactions between scaffold proteins, noncoding RNAs, and genome loci induce liquid-liquid phase separation as organizing principle for 3-dimensional nuclear architecture: Implications in cancer. FASEB J. 2019, 33, 5814–5822. [Google Scholar] [CrossRef]
- Dobersch, S.; Rubio, K.; Singh, I.; Günther, S.; Graumann, J.; Cordero, J.; Castillo-Negrete, R.; Huynh, M.B.; Mehta, A.; Braubach, P.; et al. Positioning of nucleosomes containing γ-H2AX precedes active DNA demethylation and transcription initiation. Nat. Commun. 2021, 12, 1072. [Google Scholar] [CrossRef] [PubMed]
- Rogler, G.; Vavricka, S. Exposome in IBD: Recent insights in environmental factors that influence the onset and course of IBD. Inflamm. Bowel Dis. 2015, 21, 400–408. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suñer, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.C.; Houseman, E.A.; Marsit, C.; Zheng, S.; Wrensch, M.R.; Wiemels, J.L.; Nelson, H.; Karagas, M.R.; Padbury, J.F.; Bueno, R.; et al. Aging and environmental exposures alter tissue-specific DNA methylation dependent upon CPG island context. PLoS Genet. 2009, 5, e1000602. [Google Scholar] [CrossRef] [Green Version]
- Szarc Vel Szic, K.; Declerck, K.; Vidaković, M.; Vanden Berghe, W. From inflammaging to healthy aging by dietary lifestyle choices: Is epigenetics the key to personalized nutrition? Clin. Epigenetics 2015, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Wild, C.P. Complementing the genome with an “exposome”: The outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappaport, S.M. Implications of the exposome for exposure science. J. Expo. Sci. Environ. Epidemiol. 2011, 21, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Waterland, R.A.; Michels, K.B. Epigenetic epidemiology of the developmental origins hypothesis. Annu. Rev. Nutr. 2007, 27, 363–388. [Google Scholar] [CrossRef]
- Barclay, A.R.; Russell, R.K.; Wilson, M.L.; Gilmour, W.H.; Satsangi, J.; Wilson, D.C. Systematic Review: The Role of Breastfeeding in the Development of Pediatric Inflammatory Bowel Disease. J. Pediatr. 2009, 155, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Tysk, C.; Lindberg, E.; Jarnerot, G.; Floderus-Myrhed, B. Ulcerative colitis and Crohn’s disease in an unselected population of monozygotic and dizygotic twins. A study of heritability and the influence of smoking. Gut 1988, 29, 990–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piovani, D.; Danese, S.; Peyrin-Biroulet, L.; Nikolopoulos, G.K.; Lytras, T.; Bonovas, S. Environmental Risk Factors for Inflammatory Bowel Diseases: An Umbrella Review of Meta-Analyses. Gastroenterology 2019, 157, 647–659.e4. [Google Scholar] [CrossRef] [Green Version]
- Boyko, E.J.; Theis, M.K.; Vaughan, T.L.; Nicol-blades, B. Increased risk of inflammatory bowel disease associated with oral contraceptive use. Am. J. Epidemiol. 1994, 140, 268–278. [Google Scholar] [CrossRef]
- Shaw, S.Y.; Blanchard, J.F.; Bernstein, C.N. Association between the use of antibiotics and new diagnoses of Crohn’s disease and ulcerative colitis. Am. J. Gastroenterol. 2011, 106, 2133–2142. [Google Scholar] [CrossRef]
- Bernstein, C.N. Psychological Stress and Depression: Risk Factors for IBD? Dig. Dis. 2016, 34, 58–63. [Google Scholar] [CrossRef]
- Singh, S.; Graff, L.A.; Bernstein, C.N. Do NSAIDs, antibiotics, infections, or stress trigger flares in IBD? Am. J. Gastroenterol. 2009, 104, 1298–1313. [Google Scholar] [CrossRef]
- DeFilippis, E.M.; Longman, R.; Harbus, M.; Dannenberg, K.; Scherl, E.J. Crohn’s Disease: Evolution, Epigenetics, and the Emerging Role of Microbiome-Targeted Therapies. Curr. Gastroenterol. Rep. 2016, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, C. Inflammatory bowel disease pathogenesis: Where are we? J. Gastroenterol. Hepatol. 2015, 30 (Suppl. 1), 12–18. [Google Scholar] [CrossRef] [PubMed]
- Ray, G.; Longworth, M.S. Epigenetics, DNA Organization, and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2019, 25, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.E.; Meissner, A.; Lander, E.S. The Mammalian Epigenome. Cell 2007, 128, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. Recent progress in the epigenetics and chromatin field. Cell Res. 2011, 21, 373–374. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, N.; Singh, I.; Mehta, A.; Braun, T.; Barreto, G. HMGA proteins as modulators of chromatin structure during transcriptional activation. Front. Cell Dev. Biol. 2014, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Richmond, T.J.; Finch, J.T.; Rushton, B.; Rhodes, D.; Klug, A. Structure of the nucleosome core particle at 7 resolution. Nature 1984, 311, 532–537. [Google Scholar] [CrossRef]
- Bentley, G.A.; Lewit-Bentley, A.; Finch, J.T.; Podjarny, A.D.; Roth, M. Crystal structure of the nucleosome core particle at 16 Å resolution. J. Mol. Biol. 1984, 176, 55–75. [Google Scholar] [CrossRef]
- Singh, I.; Contreras, A.; Cordero, J.; Rubio, K.; Dobersch, S.; Günther, S.; Jeratsch, S.; Mehta, A.; Krüger, M.; Graumann, J.; et al. MiCEE is a ncRNA-protein complex that mediates epigenetic silencing and nucleolar organization. Nat. Genet. 2018, 50, 990–1001. [Google Scholar] [CrossRef]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K. Environmental epigenetics and a unified theory of the molecular aspects of evolution: A neo-Lamarckian concept that facilitates neo-Darwinian evolution. Genome Biol. Evol. 2015, 7, 1296–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobersch, S.; Rubio, K.; Barreto, G. Pioneer Factors and Architectural Proteins Mediating Embryonic Expression Signatures in Cancer. Trends Mol. Med. 2019, 25, 287–302. [Google Scholar] [CrossRef]
- Esteller, M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat. Rev. Genet. 2007, 8, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar] [CrossRef]
- Bird, A.P.; Wolffe, A.P. Methylation-induced repression-belts, braces, and chromatin. Cell 1999, 99, 451–454. [Google Scholar] [CrossRef] [Green Version]
- Gardiner-Garden, M.; Frommer, M. CpG Islands in vertebrate genomes. J. Mol. Biol. 1987, 196, 261–282. [Google Scholar] [CrossRef]
- Lim, D.H.K.; Maher, E.R. DNA methylation: A form of epigenetic control of gene expression. Obstet. Gynaecol. 2010, 12, 37–42. [Google Scholar] [CrossRef]
- Yang, I.V.; Schwartz, D.A. Epigenetic control of gene expression in the lung. Am. J. Respir. Crit. Care Med. 2011, 183, 1295–1301. [Google Scholar] [CrossRef] [Green Version]
- Reik, W.; Dean, W.; Walter, J. Epigenetic Reprogramming in Mammalian Development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.A.; Liang, G. Rethinking how DNA methylation patterns are maintained. Nat. Rev. Genet. 2009, 10, 805–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef]
- Jin, B.; Robertson, K.D. DNA methyltransferases, DNA damage repair, and cancer. Adv. Exp. Med. Biol. 2013, 754, 3–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Nimmo, E.R.; Prendergast, J.G.; Aldhous, M.C.; Kennedy, N.A.; Henderson, P.; Drummond, H.E.; Ramsahoye, B.H.; Wilson, D.C.; Semple, C.A.; Satsangi, J. Genome-wide methylation profiling in Crohn’s disease identifies altered epigenetic regulation of key host defense mechanisms including the Th17 pathway. Inflamm. Bowel Dis. 2012, 18, 889–899. [Google Scholar] [CrossRef]
- Häsler, R.; Feng, Z.; Bäckdahl, L.; Spehlmann, M.E.; Franke, A.; Teschendorff, A.; Rakyan, V.K.; Down, T.A.; Wilson, G.A.; Feber, A.; et al. A functional methylome map of ulcerative colitis. Genome Res. 2012, 22, 2130–2137. [Google Scholar] [CrossRef] [Green Version]
- Cooke, J.; Zhang, H.; Greger, L.; Silva, A.L.; Massey, D.; Dawson, C. Mucosal genome-wide methylation changes in inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 2128–2137. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Kato, J.; Hiraoka, S.; Horii, J.; Suzuki, H.; Higashi, R.; Kaji, E.; Kondo, Y.; Yamamoto, K. DNA methylation of colon mucosa in ulcerative colitis patients: Correlation with inflammatory status. Inflamm. Bowel Dis. 2011, 17, 1955–1965. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.; Nagy-Szakal, R.; Mir, S.A.V.; Frank, E.; Szigeti, R.; Kaplan, J.L.; Bronsky, J.; Opekun, A.; Ferry, G.D.; Winter, H.; et al. DNA methylation-associated colonic mucosal immune and defense responses in treatmentnaïve pediatric ulcerative colitis. Epigenetics 2014, 9, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, K.; Alonso, S.; Miyaki, Y.; Okada, S.; Ogura, H.; Shiiya, N.; Konishi, F.; Taya, T.; Perucho, M.; Suzuki, K. Array-based identification of common DNA methylation alterations in ulcerative colitis. Int. J. Oncol. 2012, 40, 983–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Hegarty, J.P.; Yu, W.; Cappel, J.A.; Chen, X.; Faber, P.W.; Wang, Y.; Poritz, L.S.; Fan, J.-B.; Koltun, W.A. Identification of disease-associated DNA methylation in B cells from Crohn’s disease and ulcerative colitis patients. Dig. Dis. Sci. 2012, 57, 3145–3153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fischle, W.; Cheung, W.; Jacobs, S.; Khorasanizadeh, S.; Allis, C.D. Beyond the double helix: Writing and reading the histone code. Novartis Found. Symp. 2004, 259, 3–17; discussion 17–21, 163–169. [Google Scholar] [PubMed]
- Singh, I.; Ozturk, N.; Cordero, J.; Mehta, A.; Hasan, D.; Cosentino, C.; Sebastian, C.; Krüger, M.; Looso, M.; Carraro, G.; et al. High mobility group protein-mediated transcription requires DNA damage marker γ-H2AX. Cell Res. 2015, 25, 837–850. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhao, J.; Lv, Y.; Wang, W.; Feng, C.; Zou, W.; Su, L.; Jiao, J. Histone Variants and Histone Modifications in Neurogenesis. Trends Cell Biol. 2020, 30, 869–880. [Google Scholar] [CrossRef]
- Nakayama, T.; Takami, Y. Participation of histones and histone-modifying enzymes in cell functions through alterations in chromatin structure. J. Biochem. 2001, 129, 491–499. [Google Scholar] [CrossRef]
- Morgan, M.A.J.; Shilatifard, A. Reevaluating the roles of histone-modifying enzymes and their associated chromatin modifications in transcriptional regulation. Nat. Genet. 2020, 52, 1271–1281. [Google Scholar] [CrossRef]
- Stillman, B. Histone Modifications: Insights into Their Influence on Gene Expression. Cell 2018, 175, 6–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, L.; Wu, H.J.; Hsu, J.M.; Chang, S.S.; LaBaff, A.M.; Li, C.W.; Wang, Y.; Hsu, J.L.; Hung, M.C. Long non-coding RNAs: Versatile master regulators of gene expression and crucial players in cancer. Am. J. Transl. Res. 2012, 4, 127–150. [Google Scholar]
- Aalto, A.P.; Pasquinelli, A.E. Small non-coding RNAs mount a silent revolution in gene expression. Curr. Opin. Cell Biol. 2012, 24, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Pekow, J.R.; Kwon, J.H. MicroRNAs in inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Bjerrum, J.T.; Seidelin, J.B.; Nielsen, O.H. MicroRNAs in inflammatory bowel disease-pathogenesis, diagnostics and therapeutics. World J. Gastroenterol. 2012, 18, 4629–4634. [Google Scholar] [CrossRef] [PubMed]
- Annese, V. Genetics and epigenetics of IBD. Pharmacol. Res. 2020, 159, 104892. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, R.; Zilbauer, M. The Gut Microbiome and the Triple Environmental Hit Concept of Inflammatory Bowel Disease Pathogenesis. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Zong, D.; Liu, X.; Li, J.; Ouyang, R.; Chen, P. The role of cigarette smoke-induced epigenetic alterations in inflammation. Epigenetics Chromatin 2019, 12, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciniak, A.; Patro-Małysza, J.; Kimber-Trojnar, Ż.; Marciniak, B.; Oleszczuk, J.; Leszczyńska-Gorzelak, B. Fetal programming of the metabolic syndrome. Taiwan. J. Obstet. Gynecol. 2017, 56, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Dunford, A.R.; Sangster, J.M. Maternal and paternal periconceptional nutrition as an indicator of offspring metabolic syndrome risk in later life through epigenetic imprinting: A systematic review. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, S655–S662. [Google Scholar] [CrossRef]
- Rasmussen, L.; Knorr, S.; Antoniussen, C.S.; Bruun, J.M.; Ovesen, P.G.; Fuglsang, J.; Kampmann, U. The impact of lifestyle, diet and physical activity on epigenetic changes in the offspring—A systematic review. Nutrients 2021, 13, 2821. [Google Scholar] [CrossRef]
- Aguilera, O.; Fernández, A.F.; Muñoz, A.; Fraga, M.F. Epigenetics and environment: A complex relationship. J. Appl. Physiol. 2010, 109, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Tschurtschenthaler, M.; Kachroo, P.; Heinsen, F.A.; Adolph, T.E.; Rühlemann, M.C.; Klughammer, J.; Offner, F.A.; Ammerpohl, O.; Krueger, F.; Smallwood, S.; et al. Paternal chronic colitis causes epigenetic inheritance of susceptibility to colitis. Sci. Rep. 2016, 6, 31640. [Google Scholar] [CrossRef]
- Morgan, H.D.; Sutherland, H.G.E.; Martin, D.I.K.; Whitelaw, E. Epigenetic inheritance at the agouti locus in the mouse. Nat. Genet. 1999, 23, 314–318. [Google Scholar] [CrossRef]
- Youngson, N.A.; Whitelaw, E. Transgenerational epigenetic effects. Annu. Rev. Genom. Hum. Genet. 2008, 9, 233–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaible, T.D.; Harris, R.A.; Dowd, S.E.; Smith, C.W.; Kellermayer, R. Maternal methyl-donor supplementation induces prolonged murine offspring colitis susceptibility in association with mucosal epigenetic and microbiomic changes. Hum. Mol. Genet. 2011, 20, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterland, R.A.; Jirtle, R.L. Transposable Elements: Targets for Early Nutritional Effects on Epigenetic Gene Regulation. Mol. Cell. Biol. 2003, 23, 5293–5300. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.; Lumey, L.H.; Talens, R.P.; Kremer, D.; Putter, H.; Stein, A.; Slagboom, P.; Heijmans, B.T. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Lumey, L.H.; Slagboom, P.E. The epigenome: Archive of the prenatal environment. Epigenetics 2009, 4, 526–531. [Google Scholar] [CrossRef] [Green Version]
- Ventham, N.T.; Kennedy, N.A.; Nimmo, E.R.; Satsangi, J. Beyond gene discovery in inflammatory bowel disease: The emerging role of epigenetics. Gastroenterology 2013, 145, 293–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blewitt, M.E.; Vickaryous, N.K.; Paldi, A.; Koseki, H.; Whitelaw, E. Dynamic reprogramming of DNA methylation at an epigenetically sensitive allele in mice. PLoS Genet. 2006, 2, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Grossniklaus, U.; Kelly, B.; Ferguson-Smith, A.C.; Pembrey, M.; Lindquist, S. Transgenerational epigenetic inheritance: How important is it? Nat. Rev. Genet. 2013, 14, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, N.; Dean, W.; Erhardt, S.; Hajkova, P.; Surani, A.; Reik, W. Resistance of IAPs to methylation reprogramming may provide a mechanism for epigenetic inheritance in the mouse. Genesis 2003, 35, 88–93. [Google Scholar] [CrossRef]
- Petronis, A. Epigenetics as a unifying principle in the aetiology of complex traits and diseases. Nature 2010, 465, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Einstein, F.H. Multigenerational effects of maternal undernutrition. Cell Metab. 2014, 19, 893–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Sundquist, J.; Hemminki, K.; Sundquist, K. Risk of inflammatory bowel disease in first- and second-generation immigrants in Sweden: A nationwide follow-up study. Inflamm. Bowel Dis. 2011, 17, 1784–1791. [Google Scholar] [CrossRef]
- Pinsk, V.; Lemberg, D.A.; Grewal, K.; Barker, C.C.; Schreiber, R.A.; Jacobson, K. Inflammatory bowel disease in the South Asian pediatric population of British Columbia. Am. J. Gastroenterol. 2007, 102, 1077–1083. [Google Scholar] [CrossRef]
- Benchimol, E.I.; Mack, D.R.; Guttmann, A.; Nguyen, G.C.; To, T.; Mojaverian, N.; Quach, P.; Manuel, D.G. Inflammatory bowel disease in immigrants to Canada and their children: A population-based cohort study. Am. J. Gastroenterol. 2015, 110, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.I.; McFadden, T.; Link, V.M.; Han, S.J.; Karlsson, R.M.; Stacy, A.; Farley, T.K.; Lima-Junior, D.S.; Harrison, O.J.; Desai, J.V.; et al. Prenatal maternal infection promotes tissue-specific immunity and inflammation in offspring. Science 2021, 373, eabf3002. [Google Scholar] [CrossRef]
- Wiklund, P.; Karhunen, V.; Richmond, R.C.; Parmar, P.; Rodriguez, A.; De Silva, M.; Wielscher, M.; Rezwan, F.I.; Richardson, T.G.; Veijola, J.; et al. DNA methylation links prenatal smoking exposure to later life health outcomes in offspring. Clin. Epigenetics 2019, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure-Associated DNA Methylation in Human Populations. Annu. Rev. Public Health 2018, 39, 309–333. [Google Scholar] [CrossRef] [Green Version]
- Flom, J.D.; Ferris, J.S.; Liao, Y.; Tehranifar, P.; Richards, C.B.; Cho, Y.H.; Gonzalez, K.; Santella, R.M.; Terry, M.B. Prenatal smoke exposure and genomic DNA methylation in a multiethnic birth cohort. Cancer Epidemiol. Biomark. Prev. 2011, 20, 2518–2523. [Google Scholar] [CrossRef] [Green Version]
- Breton, C.V.; Byun, H.M.; Wenten, M.; Pan, F.; Yang, A.; Gilliland, F.D. Prenatal tobacco smoke exposure affects global and gene-specific DNA methylation. Am. J. Respir. Crit. Care Med. 2009, 180, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Preston, R.; Goldman, L.R.; Brebi-Mieville, P.; Ili, C.; Lebron, C.; Witter, F.R.; Apelberg, B.J.; Hernández-Roystacher, M.; Jaffe, A.; Halden, R.; et al. Global DNA hypomethylation is associated with in utero exposure to cotinine and perfluorinated alkyl compounds. Epigenetics 2010, 5, 539–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, B.G.; Gyselaers, W.; Byun, H.-M.; Roels, H.A.; Cuypers, A.; Baccarelli, A.A.; Nawrot, T.S. Placental mitochondrial DNA and CYP1A1 gene methylation as molecular signatures for tobacco smoke exposure in pregnant women and the relevance for birth weight. J. Transl. Med. 2017, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Begum, R.; Thota, S.; Batra, S. A systematic review of smoking-related epigenetic alterations. Arch. Toxicol. 2019, 93, 2715–2740. [Google Scholar] [CrossRef] [PubMed]
- Richmond, R.C.; Suderman, M.; Langdon, R.; Relton, C.L.; Smith, G.D. DNA methylation as a marker for prenatal smoke exposure in adults. Int. J. Epidemiol. 2018, 47, 1120–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herberth, G.; Bauer, M.; Gasch, M.; Hinz, D.; Röder, S.; Olek, S.; Kohajda, T.; Rolle-Kampczyk, U.; von Bergen, M.; Sack, U.; et al. Maternal and cord blood miR-223 expression associates with prenatal tobacco smoke exposure and low regulatory T-cell numbers. J. Allergy Clin. Immunol. 2014, 133, 543–550.e4. [Google Scholar] [CrossRef]
- Jenkins, T.G.; James, E.R.; Alonso, D.F.; Hoidal, J.R.; Murphy, P.J.; Hotaling, J.M.; Cairns, B.R.; Carrell, D.T.; Aston, K.I. Cigarette smoking significantly alters sperm DNA methylation patterns. Andrology 2017, 5, 1089–1099. [Google Scholar] [CrossRef]
- Knopik, V.S.; Maccani, M.A.; Francazio, S.; McGeary, J.E. The epigenetics of maternal cigarette smoking during pregnancy and effects on child development. Dev. Psychopathol. 2012, 24, 1377–1390. [Google Scholar] [CrossRef] [Green Version]
- Suter, M.; Ma, J.; Harris, A.S.; Patterson, L.; Brown, K.A.; Shope, C.; Showalter, L.; Abramovici, A.; Aagaard-Tillery, K.M. Maternal tobacco use modestly alters correlated epigenome-wide placental DNA methylation and gene expression. Epigenetics 2011, 6, 1284–1294. [Google Scholar] [CrossRef] [Green Version]
- Ventham, N.T.; IBD BIOM consortium; Kennedy, N.; Adams, A.T.; Kalla, R.; Heath, S.C.; O’Leary, K.R.; Drummond, H.E.; Wilson, D.C.; Gut, I.G.; et al. Integrative epigenome-wide analysis demonstrates that DNA methylation may mediate genetic risk in inflammatory bowel disease. Nat. Commun. 2016, 7, 13507. [Google Scholar] [CrossRef] [Green Version]
- Kellermayer, R. Epigenetics and the developmental origins of inflammatory bowel diseases. Can. J. Gastroenterol. 2012, 26, 909–915. [Google Scholar] [CrossRef]
- Guo, F.; Cai, D.; Li, Y.; Gu, H.; Qu, H.; Zong, Q.; Bao, W.; Chen, A.; Liu, H.-Y. How Early-Life Gut Microbiota Alteration Sets Trajectories for Health and Inflammatory Bowel Disease? Front. Nutr. 2021, 8, 690073. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Blaser, M.J.; Ley, R.E.; Knight, R. Development of the human gastrointestinal microbiota and insights from high-throughput sequencing. Gastroenterology 2011, 140, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.-H.; Gadkari, M.; Zhou, Q.; Yu, S.; Gao, N.; Guan, Y.; Schady, D.; Roshan, T.N.; Chen, M.-H.; Laritsky, E.; et al. Postnatal epigenetic regulation of intestinal stem cells requires DNA methylation and is guided by the microbiome. Genome Biol. 2015, 16, 211. [Google Scholar] [CrossRef] [Green Version]
- Poupeau, A.; Garde, C.; Sulek, K.; Citirikkaya, K.; Treebak, J.T.; Arumugam, M.; Simar, D.; Olofsson, L.E.; Bäckhed, F.; Barrès, R. Genes controlling the activation of natural killer lymphocytes are epigenetically remodeled in intestinal cells from germ-free mice. FASEB J. 2019, 33, 2719–2731. [Google Scholar] [CrossRef] [Green Version]
- Amatullah, H.; Jeffrey, K.L. Epigenome-metabolome-microbiome axis in health and IBD. Curr. Opin. Microbiol. 2020, 56, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, C.-Z.; Wan, J.-Y.; Yao, H.; Yuan, C.-S. Dissecting the interplay mechanism between epigenetics and gut microbiota: Health maintenance and disease prevention. Int. J. Mol. Sci. 2021, 22, 6933. [Google Scholar] [CrossRef]
- Liu, S.; da Cunha, A.P.; Rezende, R.M.; Cialic, R.; Wei, Z.; Bry, L.; Comstock, L.E.; Gandhi, R.; Weiner, H.L. The Host Shapes the Gut Microbiota via Fecal MicroRNA. Cell Host Microbe 2016, 19, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, I.; Raddatz, G.; Gutekunst, J.; Ridnik, M.; Cohen, D.; Abu-Remaileh, M.; Tuganbaev, T.; Shapiro, H.; Pikarsky, E.; Elinav, E.; et al. The microbiota programs DNA methylation to control intestinal homeostasis and inflammation. Nat. Microbiol. 2020, 5, 610–619. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species roseburia hominis and faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef]
- Frank, D.N.; St. Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate production by probiotic bacteria. Nutrients 2011, 3, 118–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, R.B. Microbial Influences in Inflammatory Bowel Diseases. Gastroenterology 2008, 134, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Meconi, S.; Vercellone, A.; Levillain, F.; Payré, B.; Al Saati, T.; Capilla, F.; Desreumaux, P.; Darfeuille-Michaud, A.; Altare, F. Adherent-invasive Escherichia coli isolated from Crohn’s disease patients induce granulomas in vitro. Cell. Microbiol. 2007, 9, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Dalmasso, G.; Müller, S.; Carrière, J.; Seibold, F.; Darfeuille-Michaud, A. Crohn’s disease-associated adherent invasive escherichia coli modulate levels of microRNAs in intestinal epithelial cells to reduce autophagy. Gastroenterology 2014, 146, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Larabi, A.; Dalmasso, G.; Delmas, J.; Barnich, N.; Nguyen, H.T.T. Exosomes transfer miRNAs from cell-to-cell to inhibit autophagy during infection with Crohn’s disease-associated adherent-invasive E. Coli. Gut Microbes 2020, 11, 1677–1694. [Google Scholar] [CrossRef]
- Guo, Z.; Cai, X.; Guo, X.; Xu, Y.; Gong, J.; Li, Y.; Zhu, W. Let-7b ameliorates Crohn’s disease-associated adherent-invasive E coli induced intestinal inflammation via modulating Toll-Like Receptor 4 expression in intestinal epithelial cells. Biochem. Pharmacol. 2018, 156, 196–203. [Google Scholar] [CrossRef]
- Timms, V.J.; Daskalopoulos, G.; Mitchell, H.M.; Neilan, B.A. The association of mycobacterium avium subsp. paratuberculosis with inflammatory bowel disease. PLoS ONE 2016, 11, e0148731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostoufi-Afshar, S.; Tabatabaei, M.; Ghahramani Seno, M.M. Mycobacterium avium subsp. Paratuberculosis induces differential cytosine methylation at miR-21 transcription start site region. Iran. J. Vet. Res. 2018, 19, 262–269. [Google Scholar]
- Qiao, C.X.; Xu, S.; Wang, D.D.; Gao, S.Y.; Zhao, S.F.; Zhang, M.L.; Yu, B.; Yin, Q.; Zhao, G. MicroRNA-19b alleviates lipopolysaccharide-induced inflammatory injury in human intestinal cells by up-regulation of Runx3. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5284–5294. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, D.; Yu, H.; Li, Q. MicroRNA-497 inhibits inflammation in DSS-induced IBD model mice and lipopolysaccharide-induced RAW264.7 cells via Wnt/β-catenin pathway. Int. Immunopharmacol. 2021, 101, 108318. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Xing, K.; Qi, C.; Yan, K.; Xu, Y. Down-regulation of mir-215 attenuates lipopolysaccharide-induced inflammatory injury in ccd-18co cells by targeting gdf11 through the tlr4/nf-kb and jnk/p38 signaling pathways. Histol. Histopathol. 2020, 35, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pan, S.; Luo, W.; Shen, Z.; Meng, X.; Xiao, M.; Tan, B.; Nie, K.; Tong, T.; Wang, X. Roseburia intestinalis-derived flagellin ameliorates colitis by targeting miR-223-3p-mediated activation of NLRP3 inflammasome and pyroptosis. Mol. Med. Rep. 2020, 22, 2695–2704. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Song, K.; Zhang, Y.; Zhu, C.; Shen, Z.; Wu, S.; Luo, W.; Tan, B.; Yang, Z.; Wang, X. Roseburia intestinalis-derived flagellin is a negative regulator of intestinal inflammation. Biochem. Biophys. Res. Commun. 2018, 501, 791–799. [Google Scholar] [CrossRef]
- Xue, X.; Feng, T.; Yao, S.; Wolf, K.J.; Liu, C.-G.; Liu, X. Microbiota Downregulates Dendritic Cell Expression of miR-10a, Which Targets IL-12/IL-23p40. J. Immunol. 2011, 187, 5879–5886. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Cao, A.T.; Cao, X.; Yao, S.; Carlsen, E.D.; Soong, L. Downregulation of microRNA-107 in intestinal CD11c+ myeloid cells in response to microbiota and proinflammatory cytokines increases IL-23p19 expression. Eur. J. Immunol. 2014, 44, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; He, C.; Liu, C.; Cao, A.T.; Xue, X.; Evans-Marin, H.L. miR-10a inhibits dendritic cell activation and Th1/Th17 cell immune responses in IBD. Gut 2015, 64, 1755–1764. [Google Scholar] [CrossRef]
- Nakata, K.; Sugi, Y.; Narabayashi, H.; Kobayakawa, T.; Nakanishi, Y.; Tsuda, M. Commensal Microbiota-induced microRNA modulates intestinal epithelial permeability through the small GTPase ARF4. J. Biol. Chem. 2017, 292, 15426–15433. [Google Scholar] [CrossRef] [Green Version]
- Johnston, D.G.W.; Williams, M.A.; Thaiss, C.A.; Cabrera-Rubio, R.; Raverdeau, M.; McEntee, C. Loss of microRNA-21 influences the gut microbiota, causing reduced susceptibility in a murine model of colitis. J. Crohn’s Colitis 2018, 12, 835–848. [Google Scholar] [CrossRef]
- Ryan, F.J.; Ahern, A.M.; Fitzgerald, R.S.; Laserna-Mendieta, E.J.; Power, E.M.; Clooney, A.G. Colonic microbiota is associated with inflammation and host epigenomic alterations in inflammatory bowel disease. Nat. Commun. 2020, 11, 1512. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.K.; Han, D.H.; Jang, Y.J.; Park, S.; Jang, S.J.; Lee, G. Alleviation of DSS-induced colitis: Via Lactobacillus acidophilus treatment in mice. Food Funct. 2021, 12, 340–350. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhou, L.; Wang, Y.; Dorfman, R.G.; Tang, D.; Xu, L. Faecalibacterium prausnitzii produces butyrate to decrease c-Myc-related metabolism and Th17 differentiation by inhibiting histone deacetylase 3. Int. Immunol. 2019, 31, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, L.; Liu, X.; Zhang, Y.Y.; Shi, H.; Jia, W. Extracellular Vesicles Derived from Trichinella spiralis Muscle Larvae Ameliorate TNBS-Induced Colitis in Mice. Front. Immunol. 2020, 11, 1174. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Z.; Yan, Y.; Ji, L.; He, J.; Xuan, B. Enterotoxigenic Bacteroides fragilis Promotes Intestinal Inflammation and Malignancy by Inhibiting Exosome-Packaged miR-149-3p. Gastroenterology 2021, 161, 1552–1566.e12. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.M.; Davis, J.P.; Lee, S.E.; Schaner, M.R.; Gipson, G.R.; Weiser, M. Alterations to chromatin in intestinal macrophages link IL-10 deficiency to inappropriate inflammatory responses. Eur. J. Immunol. 2016, 46, 1912–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzola, A.; González, R.; Gámez-Belmonte, R.; Ocón, B.; Aranda, C.J.; Martínez-Moya, P. miR-146a regulates the crosstalk between intestinal epithelial cells, microbial components and inflammatory stimuli. Sci. Rep. 2018, 8, 17350. [Google Scholar] [CrossRef]
- Geng, H.; Bu, H.F.; Liu, F.; Wu, L.; Pfeifer, K.; Chou, P.M. In Inflamed Intestinal Tissues and Epithelial Cells, Interleukin 22 Signaling Increases Expression of H19 Long Noncoding RNA, Which Promotes Mucosal Regeneration. Gastroenterology 2018, 155, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Lucafò, M.; Pugnetti, L.; Bramuzzo, M.; Curci, D.; Di Silvestre, A.; Marcuzzi, A. Long non-coding RNA gas5 and intestinal mmp2 and mmp9 expression: A translational study in pediatric patients with IBD. Int. J. Mol. Sci. 2019, 20, 5280. [Google Scholar] [CrossRef] [Green Version]
- Kempinska-Podhorodecka, A.; Adamowicz, M.; Ostrycharz, E.; Chmielarz, M.; Wójcicki, M.; Milkiewicz, P. Role of miR-506 in ulcerative colitis associated with primary sclerosing cholangitis. Sci. Rep. 2021, 11, 10134. [Google Scholar] [CrossRef]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilotta, A.J.; Ma, C.; Yang, W.; Yu, Y.; Yu, Y.; Zhao, X.; Zhou, Z.; Yao, S.; Dann, S.M.; Cong, Y. Propionate Enhances Cell Speed and Persistence to Promote Intestinal Epithelial Turnover and Repair. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1023–1044. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; Van Der Veeken, J.; DeRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.R.; Gonçalves, P.; Magro, F.; Martel, F. Microbiota-derived butyrate regulates intestinal inflammation: Focus on inflammatory bowel disease. Pharmacol. Res. 2020, 159, 104947. [Google Scholar] [CrossRef] [PubMed]
- Plöger, S.; Stumpff, F.; Penner, G.B.; Schulzke, J.-D.; Gäbel, G.; Martens, H.; Shen, Z.; Günzel, D.; Aschenbach, J.R. Microbial butyrate and its role for barrier function in the gastrointestinal tract. Ann. N. Y. Acad. Sci. 2012, 1258, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, C.H.; Lam, W.; Ma, D.L.; Gullen, E.A.; Cheng, Y.C. Butyrate mediates nucleotide-binding and oligomerisation domain (NOD) 2-dependent mucosal immune responses against peptidoglycan. Eur. J. Immunol. 2009, 39, 3529–3537. [Google Scholar] [CrossRef]
- Schulthess, J.; Pandey, S.; Capitani, M.; Rue-Albrecht, K.C.; Arnold, I.; Franchini, F.; Chomka, A.; Ilott, N.E.; Johnston, D.G.W.; Pires, E.; et al. The Short Chain Fatty Acid Butyrate Imprints an Antimicrobial Program in Macrophages. Immunity 2019, 50, 432–445.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef]
- Chen, L.; Sun, M.; Wu, W.; Yang, W.; Huang, X.; Xiao, Y.; Ma, C.; Xu, L.; Yao, S.; Liu, Z.; et al. Microbiota Metabolite Butyrate Differentially Regulates Th1 and Th17 Cells’ Differentiation and Function in Induction of Colitis. Inflamm. Bowel Dis. 2019, 25, 1450–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota metabolite butyrate constrains neutrophil functions and ameliorates mucosal inflammation in inflammatory bowel disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef] [PubMed]
- Berndt, B.E.; Zhang, M.; Owyang, S.Y.; Cole, T.S.; Wang, T.W.; Luther, J.; Veniaminova, N.A.; Merchant, J.L.; Chen, C.-C.; Huffnagle, G.B.; et al. Butyrate increases IL-23 production by stimulated dendritic cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Zou, F.; Qiu, Y.; Huang, Y.; Zou, H.; Cheng, X.; Niu, Q. Effects of short-chain fatty acids in inhibiting HDAC and activating p38 MAPK are critical for promoting B10 cell generation and function. Cell Death Dis. 2021, 12, 582. [Google Scholar] [CrossRef]
- Liu, Q.; Peng, Z.; Zhou, L.; Peng, R.; Li, X.; Zuo, W. Short-Chain Fatty Acid Decreases the Expression of CEBPB to Inhibit miR-145-Mediated DUSP6 and Thus Further Suppresses Intestinal Inflammation. Inflammation 2022, 45, 372–386. [Google Scholar] [CrossRef]
- Zimmerman, M.A.; Singh, N.; Martin, P.M.; Thangaraju, M.; Ganapathy, V.; Waller, J.L. Butyrate suppresses colonic inflammation through HDAC1-dependent fas upregulation and fas-mediated apoptosis of T cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, 1405–1415. [Google Scholar] [CrossRef]
- Lee, C.; Kim, B.G.; Kim, J.H.; Chun, J.; Im, J.P.; Kim, J.S. Sodium butyrate inhibits the NF-kappa B signaling pathway and histone deacetylation, and attenuates experimental colitis in an IL-10 independent manner. Int. Immunopharmacol. 2017, 51, 47–56. [Google Scholar] [CrossRef]
- Kespohl, M.; Vachharajani, N.; Luu, M.; Harb, H.; Pautz, S.; Wolff, S. The microbial metabolite butyrate induces expression of Th1-associated factors in CD4+ T cells. Front. Immunol. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, W.; Niu, J.; Yang, G.; Luo, J.; Lan, D. Heat-shock transcription factor 2 promotes sodium butyrate-induced autophagy by inhibiting mTOR in ulcerative colitis. Exp. Cell Res. 2020, 388, 111820. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-derived butyrate dynamically regulates intestinal homeostasis through regulation of actin-associated protein synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, N.; Xiong, J.; Wei, H.; Jiang, S.; Peng, J. Caprylic acid and nonanoic acid upregulate endogenous host defense peptides to enhance intestinal epithelial immunological barrier function via histone deacetylase inhibition. Int. Immunopharmacol. 2018, 65, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Krautkramer, K.A.; Kreznar, J.H.; Romano, K.A.; Vivas, E.I.; Barrett-Wilt, G.A.; Rabaglia, M.E.; Keller, M.P.; Attie, A.D.; Rey, F.E.; Denu, J.M. Diet-Microbiota Interactions Mediate Global Epigenetic Programming in Multiple Host Tissues. Mol. Cell 2016, 64, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, U.; Mustafi, R.; Zhu, H.; Zhu, X.; Deb, D.; Meredith, S.C.; Ayaloglu-Butun, F.; Fletcher, M.; Sanchez, A.; Pekow, J.; et al. Upregulation of polycistronic microRNA-143 and microRNA-145 in colonocytes suppresses colitis and inflammation-associated colon cancer. Epigenetics 2021, 16, 1317–1334. [Google Scholar] [CrossRef] [PubMed]
- Melhem, H.; Hansmannel, F.; Bressenot, A.; Battaglia-Hsu, S.-F.; Billioud, V.; Alberto, J.M.; Gueant, J.L.; Peyrin-Biroulet, L. Methyl-deficient diet promotes colitis and SIRT1-mediated endoplasmic reticulum stress. Gut 2016, 65, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Denizot, J.; Desrichard, A.; Agus, A.; Uhrhammer, N.; Dreux, N.; Vouret-Craviari, V.; Hofman, P.; Darfeuille-Michaud, A.; Barnich, N. Diet-induced hypoxia responsive element demethylation increases CEACAM6 expression, favouring Crohn’s disease-associatedEscherichia colicolonisation. Gut 2014, 64, 428–437. [Google Scholar] [CrossRef]
- Wei, M.; Gao, X.; Liu, L.; Li, Z.; Wan, Z.; Dong, Y.; Chen, X.; Niu, Y.; Zhang, J.; Yang, G. Visceral Adipose Tissue Derived Exosomes Exacerbate Colitis Severity via Pro-inflammatory MiRNAs in High Fat Diet Fed Mice. ACS Nano 2020, 14, 5099–5110. [Google Scholar] [CrossRef]
- Lv, Q.; Shi, C.; Qiao, S.; Cao, N.; Guan, C.; Dai, Y.; Wei, Z. Alpinetin exerts anti-colitis efficacy by activating AhR, regulating miR-302/DNMT-1/CREB signals, and therefore promoting Treg differentiation. Cell Death Dis. 2018, 9, 1–25. [Google Scholar] [CrossRef]
- Xiong, Y.; Qiu, J.; Li, C.; Qiu, Y.; Guo, L.; Liu, Y.; Wan, J.; Li, Y.; Wu, G.; Wang, L.; et al. Fortunellin-induced modulation of phosphatase and tensin homolog by MicroRNA-374a decreases inflammation and maintains intestinal barrier function in colitis. Front. Immunol. 2018, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, R.; Chen, J.; Wu, Q.; Kuang, Z. Baicalin protects against TNF-α-induced injury by down-regulating mir-191a that targets the tight junction protein ZO-1 in IEC-6 cells. Biol. Pharm. Bull. 2017, 40, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Galleggiante, V.; De Santis, S.; Liso, M.; Verna, G.; Sommella, E.; Mastronardi, M.; Campiglia, P.; Chieppa, M.; Serino, G. Quercetin-Induced miR-369-3p Suppresses Chronic Inflammatory Response Targeting C/EBP-β. Mol. Nutr. Food Res. 2019, 63, 1801390. [Google Scholar] [CrossRef] [PubMed]
- James, S.; Aparna, J.; Babu, A.; Paul, A.; Lankadasari, M.; Athira, S.; Kumar, S.; Vijayan, Y.; Namitha, N.; Mohammed, S.; et al. Cardamonin attenuates experimental colitis and associated colorectal cancer. Biomolecules 2021, 11, 661. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, D.J.; Yuan, W.Q.; Chen, C.; Liu, Q. Berberine represses Wnt/β-catenin pathway activation via modulating the microRNA-103a-3p/Bromodomain-containing protein 4 axis, thereby refraining pyroptosis and reducing the intestinal mucosal barrier defect induced via colitis. Bioengineered 2022, 13, 7392–7409. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Xing, Y.; Liu, J.; Dong, D.; Liu, Y.; Qiao, H.; Zhang, Y.; Hu, L. Lonicerin targets EZH2 to alleviate ulcerative colitis by autophagy-mediated NLRP3 inflammasome inactivation. Acta. Pharm. Sin. B 2021, 11, 2880–2899. [Google Scholar] [CrossRef] [PubMed]
- Alrafas, H.R.; Busbee, P.B.; Chitrala, K.N.; Nagarkatti, M.; Nagarkatti, P. Alterations in the gut microbiome and suppression of histone deacetylases by resveratrol are associated with attenuation of colonic inflammation and protection against colorectal cancer. J. Clin. Med. 2020, 9, 1796. [Google Scholar] [CrossRef]
- Alrafas, H.R.; Busbee, P.B.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol Downregulates miR-31 to Promote T Regulatory Cells during Prevention of TNBS-Induced Colitis. Mol. Nutr. Food Res. 2020, 64, 1900633. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, D.; Wan, X.; Bai, Y.; Yuan, C.; Wang, T.; Yuan, D.; Zhang, C.; Liu, C. Chlorogenic Acid Suppresses miR-155 and Ameliorates Ulcerative Colitis through the NF-κB/NLRP3 Inflammasome Pathway. Mol. Nutr. Food Res. 2020, 64, e2000452. [Google Scholar] [CrossRef]
- Benmoussa, A.; Diallo, I.; Salem, M.; Michel, S.; Gilbert, C.; Sévigny, J.; Provost, P. Concentrates of two subsets of extracellular vesicles from cow’s milk modulate symptoms and inflammation in experimental colitis. Sci. Rep. 2019, 9, 14661. [Google Scholar] [CrossRef] [Green Version]
- Reif, S.; Elbaum-Shiff, Y.; Koroukhov, N.; Shilo, I.; Musseri, M.; Golan-Gerstl, R. Cow and human milk-derived exosomes ameliorate colitis in DSS murine model. Nutrients 2020, 12, 2589. [Google Scholar] [CrossRef]
- Wu, D.; Kittana, H.; Shu, J.; Kachman, S.D.; Cui, J.; Ramer-Tait, A.E.; Zempleni, J. Dietary Depletion of Milk Exosomes and their MicroRNA Cargos Elicits a Depletion of miR-200a-3p and Elevated Intestinal Inflammation and CXCL9 Expression in Mdr1a−/− Mice. Curr. Dev. Nutr. 2019, 3, nzz122. [Google Scholar] [CrossRef]
- Zhang, X.; Gu, J.; Zhao, C.; Hu, Y.; Zhang, B.; Wang, J.; Lv, H.; Ji, X.; Wang, S. Sweeteners Maintain Epithelial Barrier Function Through the miR-15b/RECK/MMP-9 Axis, Remodel Microbial Homeostasis, and Attenuate Dextran Sodium Sulfate-Induced Colitis in Mice. J. Agric. Food Chem. 2022, 70, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liang, W.; Yang, X.; Li, Q.; Zhang, G. Cytoprotective effects of galacto-oligosaccharides on colon epithelial cells via up-regulating miR-19b. Life Sci. 2019, 231, 116589. [Google Scholar] [CrossRef]
- Zha, Z.; Lv, Y.; Tang, H.; Li, T.; Miao, Y.; Cheng, J. An orally administered butyrate-releasing xylan derivative reduces inflammation in dextran sulphate sodium-induced murine colitis. Int. J. Biol. Macromol. 2020, 156, 1217–1233. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.-L.; Chen, L.; Wen, X.-S.; Zuo, J.-P.; Wang, X.-Y.; Lu, Z.-J.; Yang, Y.-F. Suppression of Th17 cell differentiation via sphingosine-1-phosphate receptor 2 by cinnamaldehyde can ameliorate ulcerative colitis. Biomed. Pharmacother. 2021, 134, 111116. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Shen, Y.; Wang, M.; Wang, X.; Yang, Y. Suppression of miR-21 and miR-155 of macrophage by cinnamaldehyde ameliorates ulcerative colitis. Int. Immunopharmacol. 2019, 67, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, S.; Lv, X.; Lu, J.; Ren, C.; Zeng, Z. Limonin ameliorates ulcerative colitis by regulating STAT3/miR-214 signaling pathway. Int. Immunopharmacol. 2019, 75, 105768. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z. Edible ginger-derived nanoparticles: A novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, T.; Lv, X.; Zhang, J.; Liu, S. Ginsenoside Rh2 alleviates ulcerative colitis by regulating the STAT3/miR-214 signaling pathway. J. Ethnopharmacol. 2021, 274, 113997. [Google Scholar] [CrossRef]
- Wang, L.S.; Kuo, C.T.; Stoner, K.; Yearsley, M.; Oshima, K.; Yu, J. Dietary black raspberries modulate DNA methylation in dextran sodium sulfate (DSS)-induced ulcerative colitis. Carcinogenesis 2013, 34, 2842–2850. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.S.; Kuo, C.T.; Huang, T.H.M.; Yearsley, M.; Oshima, K.; Stoner, G.D. Black raspberries protectively regulate methylation of Wnt pathway genes in precancerous colon tissue. Cancer Prev. Res. 2013, 6, 1317–1327. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, C.; Liu, Y.; Tang, L.; Zheng, M.; Xu, C. MiR-122 targets NOD2 to decrease intestinal epithelial cell injury in Crohn’s disease. Biochem. Biophys. Res. Commun. 2013, 438, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Lippai, D.; Bala, S.; Catalano, D.; Kodys, K.; Szabo, G. Micro-RNA-155 Deficiency Prevents Alcohol-Induced Serum Endotoxin Increase and Small Bowel Inflammation in Mice. Alcohol. Clin. Exp. Res. 2014, 38, 2217–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Wei, Y. Modulators of microrna function in the immune system. Int. J. Mol. Sci. 2020, 21, 2357. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, D.F.; Donovan, M.G.; Doetschman, T.C.; Selmin, O.I. N-6 linoleic acid induces epigenetics alterations associated with colonic inflammation and cancer. Nutrients 2019, 11, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimier, E.; Chervy, M.; Agus, A.; Sivignon, A.; Billard, E.; Privat, M. Methyl-donor supplementation prevents intestinal colonization by Adherent-Invasive, E. coli in a mouse model of Crohn’s disease. Sci. Rep. 2020, 10, 12922. [Google Scholar] [CrossRef]
- Amerikanou, C.; Papada, E.; Gioxari, A.; Smyrnioudis, I.; Kleftaki, S.A.; Valsamidou, E. Mastiha has efficacy in immune-mediated inflammatory diseases through a microRNA-155 Th17 dependent action. Pharmacol. Res. 2021, 171, 105753. [Google Scholar] [CrossRef]
- Chi, J.H.; Seo, G.S.; Cheon, J.H.; Lee, S.H. Isoliquiritigenin inhibits TNF-α-induced release of high-mobility group box 1 through activation of HDAC in human intestinal epithelial HT-29 cells. Eur. J. Pharmacol. 2017, 796, 101–109. [Google Scholar] [CrossRef]
- Angel-Morales, G.; Noratto, G.; Mertens-Talcott, S. Red wine polyphenolics reduce the expression of inflammation markers in human colon-derived CCD-18Co myofibroblast cells: Potential role of microRNA-126. Food Funct. 2012, 3, 745–752. [Google Scholar] [CrossRef]
- Ojwang, L.O.; Banerjee, N.; Noratto, G.D.; Angel-Morales, G.; Hachibamba, T.; Awika, J.M. Polyphenolic extracts from cowpea (Vigna unguiculata) protect colonic myofibroblasts (CCD18Co cells) from lipopolysaccharide (LPS)-induced inflammation–modulation of microRNA 126. Food Funct. 2015, 6, 145–153. [Google Scholar] [CrossRef]
- Kim, H.; Banerjee, N.; Barnes, R.C.; Pfent, C.M.; Talcott, S.T.; Dashwood, R.H. Mango polyphenolics reduce inflammation in intestinal colitis—involvement of the miR-126/PI3K/AKT/mTOR axis in vitro and in vivo. Mol. Carcinog. 2017, 56, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Banerjee, N.; Sirven, M.A.; Minamoto, Y.; Markel, M.E.; Suchodolski, J.S. Pomegranate polyphenolics reduce inflammation and ulceration in intestinal colitis—involvement of the miR-145/p70S6K1/HIF1α axis in vivo and in vitro. J. Nutr. Biochem. 2017, 43, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Peng, S.; Wang, S.; Li, X.; Li, H.; Shen, L. Pristimerin reduces dextran sulfate sodium-induced colitis in mice by inhibiting microRNA-155. Int. Immunopharmacol. 2021, 94, 107491. [Google Scholar] [CrossRef] [PubMed]
- Papoutsopoulou, S.; Satsangi, J.; Campbell, B.J.; Probert, C.S. Review article: Impact of cigarette smoking on intestinal inflammation—Direct and indirect mechanisms. Aliment. Pharmacol. Ther. 2020, 51, 1268–1285. [Google Scholar] [CrossRef] [PubMed]
- Ambatipudi, S.; Cuenin, C.; Hernandez-Vargas, H.; Ghantous, A.; Le Calvez-Kelm, F.; Kaaks, R. Tobacco smoking-associated genome-wide DNA methylation changes in the EPIC study. Epigenomics 2016, 8, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitling, L.P.; Yang, R.; Korn, B.; Burwinkel, B.; Brenner, H. Tobacco-smoking-related differential DNA methylation: 27K discovery and replication. Am. J. Hum. Genet. 2011, 88, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wong, E.M.; Bui, M.; Nguyen, T.L.; Joo, J.H.E.; Stone, J. Causal effect of smoking on DNA methylation in peripheral blood: A twin and family study. Clin. Epigenetics 2018, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Wan, E.S.; Qiu, W.; Baccarelli, A.; Carey, V.J.; Bacherman, H.; Rennard, S.I. Cigarette smoking behaviors and time since quitting are associated with differential DNA methylation across the human genome. Hum. Mol. Genet. 2012, 21, 3073–3082. [Google Scholar] [CrossRef]
- McCartney, D.L.; Stevenson, A.J.; Hillary, R.F.; Walker, R.M.; Bermingham, M.L.; Morris, S.W. Epigenetic signatures of starting and stopping smoking. EBioMedicine 2018, 37, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Wahl, S.; Pfeiffer, L.; Ward-Caviness, C.K.; Kunze, S.; Kretschmer, A. The dynamics of smoking-related disturbed methylation: A two time-point study of methylation change in smokers, non-smokers and former smokers. BMC Genom. 2017, 18, 805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joehanes, R.; Just, A.C.; Marioni, R.E.; Pilling, L.C.; Reynolds, L.M.; Mandaviya, P.R. Epigenetic Signatures of Cigarette Smoking. Circ. Cardiovasc. Genet. 2016, 9, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Wang, R.; Strulovici-Barel, Y.; Salit, J.; Staudt, M.R.; Ahmed, J. Persistence of smoking-induced dysregulation of MiRNA expression in the small airway epithelium despite smoking cessation. PLoS ONE 2015, 10, e0120824. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.M.; Chase, R.P.; Lamb, A.; Reyes, A.; Saferali, A.; Yun, J.H. RNA sequencing identifies novel non-coding RNA and exon-specific effects associated with cigarette smoking. BMC Med. Genom. 2017, 10, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrijens, K.; Bollati, V.; Nawrot, T.S. MicroRNAs as potential signatures of environmental exposure or effect: A systematic review. Environ. Health Perspect. 2015, 123, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Luettich, K. MicroRNAs as potential biomarkers of smoking-related diseases. Biomark. Med. 2012, 6, 671–684. [Google Scholar] [CrossRef] [Green Version]
- National Center for Chronic Disease Prevention and Health Promotion (US) Office on Smoking and Health. The Health Consequences of Smoking—50 Years of Progress; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2014; pp. 1–36.
- Hindorff, L.A.; Sethupathy, P.; Junkins, H.A.; Ramos, E.M.; Mehta, J.P.; Collins, F.S. Potential etiologic and functional implications of genome-wide association loci for human diseases and traits. Proc. Natl. Acad. Sci. USA 2009, 106, 9362–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Xia, P.; Su, P. High-Dimensional DNA Methylation Mediates the Effect of Smoking on Crohn’s Disease. Front. Genet. 2022, 13, 302. [Google Scholar] [CrossRef]
- Andersen, A.M.; Lei, M.K.; Beach, S.R.H.; Philibert, R.A.; Sinha, S.; Colgan, J.D. Cigarette and cannabis smoking effects on GPR15+ helper T cell levels in peripheral blood: Relationships with epigenetic biomarkers. Genes 2020, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Szulakowski, P.; Crowther, A.J.L.; Jiménez, L.A.; Donaldson, K.; Mayer, R.; Leonard, T.B. The effect of smoking on the transcriptional regulation of lung inflammation in patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2006, 174, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Ito, M.; Elliott, W.M.; Cosio, B.; Caramori, G.; Kon, O.M. Decreased Histone Deacetylase Activity in Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2005, 352, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Guan, X.-J.; Peng, X.-H.; Cui, Z.-L.; Luan, C.-Y.; Guo, X.-J. Acetylation of lysine 9 on histone H3 is associated with increased pro-inflammatory cytokine release in a cigarette smoke-induced rat model through HDAC1 depression. Inflamm. Res. 2015, 64, 513–526. [Google Scholar] [CrossRef]
- Sundar, I.K.; Rahman, I. Gene expression profiling of epigenetic chromatin modification enzymes and histone marks by cigarette smoke: Implications for COPD and lung cancer. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L1245–L1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraro, M.; Di Vincenzo, S.; Dino, P.; Bucchieri, S.; Cipollina, C.; Gjomarkaj, M. Budesonide, Aclidinium and Formoterol in combination limit inflammaging processes in bronchial epithelial cells exposed to cigarette smoke. Exp. Gerontol. 2019, 118, 78–87. [Google Scholar] [CrossRef]
- To, M.; Yamamura, S.; Akashi, K.; Charron, C.E.; Haruki, K.; Barnes, P.J. Defect of adaptation to hypoxia in patients with COPD due to reduction of histone deacetylase 7. Chest 2012, 141, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, I.; Lim, J.H.; Oh, D.K.; Kim, W.J.; Oh, Y.M. Gene expression profile of human lung in a relatively early stage of COPD with emphysema. Int. J. Chronic Obstr. Pulm. Dis. 2018, 13, 2643–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, I.K.; Nevid, M.Z.; Friedman, A.E.; Rahman, I. Cigarette smoke induces distinct histone modifications in lung cells: Implications for the pathogenesis of COPD and lung cancer. J. Proteome Res. 2014, 13, 982–996. [Google Scholar] [CrossRef] [Green Version]
- Marwick, J.A.; Kirkham, P.A.; Stevenson, C.S.; Danahay, H.; Giddings, J.; Butler, K. Cigarette smoke alters chromatin remodeling and induces proinflammatory genes in rat lungs. Am. J. Respir. Cell Mol. Biol. 2004, 31, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Schug, T.T.; Xu, Q.; Gao, H.; Peres-da-Silva, A.; Draper, D.W.; Fessler, M.B. Myeloid Deletion of SIRT1 Induces Inflammatory Signaling in Response to Environmental Stress. Mol. Cell. Biol. 2010, 30, 4712–4721. [Google Scholar] [CrossRef] [Green Version]
- Lo Sasso, G.; Menzies, K.J.; Mottis, A.; Piersigilli, A.; Perino, A.; Yamamoto, H. SIRT2 deficiency modulates macrophage polarization and susceptibility to experimental colitis. PLoS ONE 2014, 9, e103573. [Google Scholar] [CrossRef]
- Winkler, A.R.; Nocka, K.N.; Williams, C.M.M. Smoke exposure of human macrophages reduces HDAC3 activity, resulting in enhanced inflammatory cytokine production. Pulm. Pharmacol. Ther. 2012, 25, 286–292. [Google Scholar] [CrossRef]
- Poralla, L.; Stroh, T.; Erben, U.; Sittig, M.; Liebig, S.; Siegmund, B. Histone deacetylase 5 regulates the inflammatory response of macrophages. J. Cell. Mol. Med. 2015, 19, 2162–2171. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, G.; Yang, X. HDAC5 promotes Mycoplasma pneumoniae-induced inflammation in macrophages through NF-κB activation. Life Sci. 2019, 221, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M. Anti-inflammatory properties of sirtuin 6 in human umbilical vein endothelial cells. Mediat. Inflamm. 2012, 2012, 597514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, H.; Luo, G.; Dai, X. SIRT4 inhibits cigarette smoke extracts-induced mononuclear cell adhesion to human pulmonary microvascular endothelial cells via regulating NF-κB activity. Toxicol. Lett. 2014, 226, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Huang, G.; Wei, T.; Gao, J.; Huang, C.; Sun, M. Sirtuin 3-induced macrophage autophagy in regulating NLRP3 inflammasome activation. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Huang, Q.; Liao, M.-J.; Xu, F.; Zhang, T.; He, J.; Zhang, L.; Liu, H.-Z. EZH2 plays a crucial role in ischemia/reperfusion-induced acute kidney injury by regulating p38 signaling. Inflamm. Res. 2019, 68, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Larghero, P.; Longobardi, M.; Cartiglia, C.; Camoirano, A.; Steele, V.E. Dose-responsiveness and persistence of microRNA expression alterations induced by cigarette smoke in mouse lung. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2011, 717, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Izzotti, A.; Larghero, P.; Balansky, R.; Pfeffer, U.; Steele, V.E.; de Flora, S. Interplay between histopathological alterations, cigarette smoke and chemopreventive agents in defining microRNA profiles in mouse lung. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2011, 717, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Wang, P.Y.; Wan, J.J.; Zhang, Y.; Wei, J.; Sun, Y. MicroRNA124-IL6R Mediates the Effect of Nicotine in Inflammatory Bowel Disease by Shifting Th1/Th2 Balance toward Th1. Front. Immunol. 2020, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, Q.; Gui, H.; Xu, D.P.; Yang, Y.L.; Su, D.F. MicroRNA-124 mediates the cholinergic anti-inflammatory action through inhibiting the production of pro-inflammatory cytokines. Cell Res. 2013, 23, 1270–1283. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Wan, J.J.; Sun, Y.; Wu, T.; Wang, P.Y.; Du, P. Nicotine protects against DSS colitis through regulating microRNA-124 and STAT3. J. Mol. Med. 2017, 95, 221–233. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, X.; Xu, Y. Aberrant expression of miR-21 in patients with inflammatory bowel disease A protocol for systematic review and meta analysis. Medicine 2020, 99, e19693. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Liang, Y.; Yang, J.; Xia, Y.; Chen, H.; Han, H. MicroRNA-21 Knockout Improve the Survival Rate in DSS Induced Fatal Colitis through Protecting against Inflammation and Tissue Injury. PLoS ONE 2013, 8, e66814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Shen, J.; Cheng, J.; Fan, X. MicroRNA-21 regulates intestinal epithelial tight junction permeability. Cell Biochem. Funct. 2015, 33, 235–240. [Google Scholar] [CrossRef]
- Smigielska-Czepiel, K.; van den Berg, A.; Jellema, P.; Slezak-Prochazka, I.; Maat, H.; van den Bos, H. Dual Role of miR-21 in CD4+ T-Cells: Activation-Induced miR-21 Supports Survival of Memory T-Cells and Regulates CCR7 Expression in Naive T-Cells. PLoS ONE 2013, 8, e76217. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Q.; Wang, P.; Wang, T.; Qi, J.; Wei, M.; Wang, S. MicroRNA-21 regulates T-cell apoptosis by directly targeting the tumor suppressor gene Tipe2. Cell Death Dis. 2014, 5, e1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, Y.; Mazzurana, L.; Forkel, M.; Okazaki, K.; Aoi, M.; Schmidt, P.T. Downregulation of MicroRNA-21 in colonic CD3+ T cells in UC remission. Inflamm. Bowel Dis. 2016, 22, 2788–2793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.X.; Munitz, A.; Rothenberg, M.E. MicroRNA-21 Is Up-Regulated in Allergic Airway Inflammation and Regulates IL-12p35 Expression. J. Immunol. 2009, 182, 4994–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimi, S.T.; Fulcher, J.A.; Chang, M.H.; Gov, L.; Wang, S.; Lee, B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 2009, 114, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheedy, F.J. Turning 21, Induction of miR-21 as a key switch in the inflammatory response. Front. Immunol. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Cao, S.; Liu, F.; Liu, Y. miR-195 plays a role in steroid resistance of ulcerative colitis by targeting Smad7. Biochem. J. 2015, 471, 357–367. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, Y.; Li, Y.; Xu, W.; Luo, F.; Wang, B. NF-κB-Mediated Inflammation Leading to EMT via miR-200c is involved in cell transformation induced by cigarette smoke extract. Toxicol. Sci. 2013, 135, 265–276. [Google Scholar] [CrossRef]
- Louis, E.; Michel, V.; Hugot, J.P.; Reenaers, C.; Fontaine, F.; Delforge, M. Early development of stricturing or penetrating pattern in Crohn’s disease is influenced by disease location, number of flares, and smoking but not by NOD2/CARD15 genotype. Gut 2003, 52, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Flier, S.N.; Tanjore, H.; Kokkotou, E.G.; Sugimoto, H.; Zeisberg, M.; Kalluri, R. Identification of epithelial to mesenchymal transition as a novel source of fibroblasts in intestinal fibrosis. J. Biol. Chem. 2010, 285, 20202–20212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharl, M.; Huber, N.; Lang, S.; Fürst, A.; Jehle, E.; Rogler, G. Hallmarks of epithelial to mesenchymal transition are detectable in Crohn’s disease associated intestinal fibrosis. Clin. Transl. Med. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthakrishnan, A.N.; Higuchi, L.M.; Huang, E.S.; Khalili, H.; Richter, J.M.; Fuchs, C.S. Aspirin, nonsteroidal anti-inflammatory drug use, and risk for crohn disease and ulcerative colitis. Ann. Intern. Med. 2012, 156, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Cornish, J.A.; Tan, E.; Simillis, C.; Clark, S.K.; Teare, J.; Tekkis, P.P. The risk of oral contraceptives in the etiology of inflammatory bowel disease: A meta-analysis. Am. J. Gastroenterol. 2008, 103, 2394–2400. [Google Scholar] [CrossRef] [PubMed]
- Minacapelli, C.D.; Bajpai, M.; Geng, X.; Van Gurp, J.; Poplin, E.; Amenta, P.S. MiR-206 as a biomarker for response to mesalamine treatment in ulcerative colitis. Inflamm. Bowel Dis. 2019, 25, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Adamowicz, M.; Milkiewicz, P.; Kempinska-Podhorodecka, A. 5-aminosalicylic acid inhibits the expression of oncomirs and pro-inflammatory microRNAS: An in vitro study. J. Physiol. Pharmacol. 2021, 72, 529–535. [Google Scholar] [CrossRef]
- He, C.; Shi, Y.; Wu, R.; Sun, M.; Fang, L.; Wu, W. MIR-301a promotes intestinal mucosal inflammation through induction of IL-17A and TNF-α in IBD. Gut 2016, 65, 1938–1950. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Zhao, Q. Lnc-ITSN1-2, Derived from RNA Sequencing, Correlates with Increased Disease Risk, Activity and Promotes CD4+ T Cell Activation, Proliferation and Th1/Th17 Cell Differentiation by Serving as a ceRNA for IL-23R via Sponging miR-125a in Inflammatory Bowel Di. Front. Immunol. 2020, 11, 852. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, L.; Hu, T.; Yin, J.; Xu, L.; Pang, Z. CircRNA_103765 acts as a proinflammatory factor via sponging miR-30 family in Crohn’s disease. Sci Rep 2021, 11, 565. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.K.; Heier, C.R.; Calderon, L.D.; Tully, C.B.; Fiorillo, A.A.; Van Der Anker, J. Serum mirnas are pharmacodynamic biomarkers associated with therapeutic response in pediatric inflammatory bowel disease. Inflamm. Bowel Dis. 2020, 26, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Dubois-Camacho, K.; Diaz-Jimenez, D.; De la Fuente, M.; Quera, R.; Simian, D.; Martínez, M. Inhibition of miR-378a-3p by Inflammation Enhances IL-33 Levels: A Novel Mechanism of Alarmin Modulation in Ulcerative Colitis. Front. Immunol. 2019, 10, 2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Gong, J.; Li, Y.; Gu, L.; Cao, L.; Wang, Z. Mucosal microRNAs expression profiles before and after exclusive enteral nutrition therapy in adult patients with Crohn’s disease. Nutrients 2016, 8, 519. [Google Scholar] [CrossRef]
- Gu, L.; Ren, F.; Fang, X.; Yuan, L.; Liu, G.; Wang, S. Exosomal MicroRNA-181a Derived from Mesenchymal Stem Cells Improves Gut Microbiota Composition, Barrier Function, and Inflammatory Status in an Experimental Colitis Model. Front. Med. 2021, 8, 898. [Google Scholar] [CrossRef]
- Yang, R.; Huang, H.; Cui, S.; Zhou, Y.; Zhang, T.; Zhou, Y. IFN-γ promoted exosomes from mesenchymal stem cells to attenuate colitis via miR-125a and miR-125b. Cell Death Dis. 2020, 11, 603. [Google Scholar] [CrossRef]
- Becker, E.; Bengs, S.; Aluri, S.; Opitz, L.; Atrott, K.; Stanzel, C. Doxycycline, metronidazole and isotretinoin: Do they modify microRNA/mRNA expression profiles and function in murine T-cells? Sci. Rep. 2016, 6, 37082. [Google Scholar] [CrossRef]
- Garrido-Mesa, J.; Rodríguez-Nogales, A.; Algieri, F.; Vezza, T.; Hidalgo-Garcia, L.; Garrido-Barros, M. Immunomodulatory tetracyclines shape the intestinal inflammatory response inducing mucosal healing and resolution. Br. J. Pharmacol. 2018, 175, 4353–4370. [Google Scholar] [CrossRef]
- Hattori, N.; Niwa, T.; Ishida, T.; Kobayashi, K.; Imai, T.; Mori, A. Antibiotics suppress colon tumorigenesis through inhibition of aberrant DNA methylation in an azoxymethane and dextran sulfate sodium colitis model. Cancer Sci. 2019, 110, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Veltman, K.; Hummel, S.; Cichon, C.; Sonnenborn, U.; Schmidt, M.A. Identification of specific miRNAs targeting proteins of the apical junctional complex that simulate the probiotic effect of E. coli Nissle 1917 on T84 epithelial cells. Int. J. Biochem. Cell Biol. 2012, 44, 341–349. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, L.; Zhang, S.; Yang, Y.; Xu, L.; Hua, Z. Bifidobacterium longum affects the methylation level of forkhead box P3 promoter in 2, 4, 6-trinitrobenzenesulphonic acid induced colitis in rats. Microb. Pathog. 2017, 110, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N. Differential intestinal anti-inflammatory effects of Lactobacillus fermentum and Lactobacillus salivarius in DSS mouse colitis: Impact on microRNAs expression and microbiota composition. Mol. Nutr. Food Res. 2017, 61, 1700144. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N. Intestinal anti-inflammatory effect of the probiotic Saccharomyces boulardii in DSS-induced colitis in mice: Impact on microRNAs expression and gut microbiota composition. J. Nutr. Biochem. 2018, 61, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Din, A.U.; Hassan, A.; Zhu, Y.; Zhang, K.; Wang, Y.; Li, T. Inhibitory effect of Bifidobacterium bifidum ATCC 29521 on colitis and its mechanism. J. Nutr. Biochem. 2020, 79, 108353. [Google Scholar] [CrossRef]
- Sun, S.; Xu, X.; Liang, L.; Wang, X.; Bai, X.; Zhu, L. Lactic Acid-Producing Probiotic Saccharomyces cerevisiae Attenuates Ulcerative Colitis via Suppressing Macrophage Pyroptosis and Modulating Gut Microbiota. Front. Immunol. 2021, 12, 4847. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhu, S.; Zhou, R.; Yi, F.; Bing, Y.; Huang, S. Effects of Sinomenine on the Expression of microRNA-155 in 2,4,6-Trinitrobenzenesulfonic Acid-Induced Colitis in Mice. PLoS ONE 2013, 8, e73757. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Li, Y.; Guo, Z.; Gong, J.; Zhu, W.; Li, N. Triptolide ameliorates ileocolonic anastomosis inflammation in IL-10 deficient mice by mechanism involving suppression of miR-155/SHIP-1 signaling pathway. Mol. Immunol. 2013, 56, 340–346. [Google Scholar] [CrossRef]
- Changlin, Z.; Ying, Z.; Shuhua, Z.; Zeng, J.; Chi, H. Research on the Protective Effect of MiR-185-3p Mediated by Huangqin-Tang Decoction (HQT) on the Epithelial Barrier Function of Ulcerative Colitis. Evid.-Based Complement. Altern. Med. 2021, 2021, 4775606. [Google Scholar] [CrossRef]
- Wei, K.; Zhang, D.; Hong, J.; Zhang, C.; Feng, X.; Huang, Y. Herb-partitioned moxibustion and the miRNAs related to Crohn’s disease: A study based on rat models. Evid.-Based Complement. Altern. Med. 2015, 2015, 265238. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, J.; Chu, H.; Chen, D.; Guo, H. Salvianolic acid B restored impaired barrier function via downregulation of MLCK by microRNA-1 in rat colitis model. Front. Pharmacol. 2016, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Ma, Z.; Cui, Y.H.; Dong, H.S.; Zhao, J.M.; Dou, C.Z. Effects of Herb-Partitioned Moxibustion on the miRNA Expression Profiles in Colon from Rats with DSS-Induced Ulcerative Colitis. Evid.-Based Complement. Altern. Med. 2017, 2017, 1767301. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.W.; Wang, J.M.; Wang, D.; Wu, R.; Ji, Z.L. Triptolide exerts protective effects against fibrosis following ileocolonic anastomosis by mechanisms involving the MIR-16-1/HSP70 pathway in IL-10-deficient mice. Int. J. Mol. Med. 2017, 40, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Q.; Wang, K.; Qiao, S.; Yang, L.; Xin, Y.; Dai, Y. Norisoboldine, a natural AhR agonist, promotes Treg differentiation and attenuates colitis via targeting glycolysis and subsequent NAD+/SIRT1/SUV39H1/H3K9me3 signaling pathway article. Cell Death Dis. 2018, 9, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Wang, J.M.; Wang, D.; Wu, R.; Hou, H.W. Triptolide inhibits migration and proliferation of fibroblasts from ileocolonic anastomosis of patients with Crohn’s disease via regulating the miR-16-1/HSP70 pathway. Mol. Med. Rep. 2019, 19, 4841–4851. [Google Scholar] [CrossRef]
- Zong, X.; Xiao, X.; Kai, L.; Cheng, Y.; Fu, J.; Xu, W. Atractylodis macrocephalae polysaccharides protect against DSS-induced intestinal injury through a novel lncRNA ITSN1-OT1. Int. J. Biol. Macromol. 2021, 167, 76–84. [Google Scholar] [CrossRef]
- Liu, H.; Liang, J.; Zhong, Y.; Xiao, G.; Efferth, T.; Georgiev, M.I. Dendrobium officinale Polysaccharide Alleviates Intestinal Inflammation by Promoting Small Extracellular Vesicle Packaging of miR-433-3p. J. Agric. Food Chem. 2021, 69, 13510–13523. [Google Scholar] [CrossRef]
- Huang, N.; Katz, J.P.; Martin, D.R.; Wu, G.D. Inhibition of IL-8 gene expression in Caco-2 cells by compounds which induce histone hyperacetylation. Cytokine 1997, 9, 27–36. [Google Scholar] [CrossRef]
- Simeoli, R.; Mattace Raso, G.; Pirozzi, C.; Lama, A.; Santoro, A.; Russo, R. An orally administered butyrate-releasing derivative reduces neutrophil recruitment and inflammation in dextran sulphate sodium-induced murine colitis. Br. J. Pharmacol. 2017, 174, 1484–1496. [Google Scholar] [CrossRef] [Green Version]
- Tazi, J.; Begon-Pescia, C.; Campos, N.; Apolit, C.; Garcel, A.; Scherrer, D. Specific and selective induction of miR-124 in immune cells by the quinoline ABX464, A transformative therapy for inflammatory diseases. Drug Discov. Today 2021, 26, 1030–1039. [Google Scholar] [CrossRef]
- Chen, Q.; Duan, X.; Xu, M.; Fan, H.; Dong, Y.; Wu, H. BMSC-EVs regulate Th17 cell differentiation in UC via H3K27me3. Mol. Immunol. 2020, 118, 191–200. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Cao, J.; Ye, C. Exosomes from Adipose-Derived Stem Cells Promotes VEGF-C-Dependent Lymphangiogenesis by Regulating miRNA-132/TGF-β Pathway. Cell Physiol. Biochem. 2018, 49, 160–171. [Google Scholar] [CrossRef]
- Horniblow, R.D.; Pathak, P.; Balacco, D.L.; Acharjee, A.; Lles, E.; Gkoutos, G. Iron-mediated epigenetic activation of NRF2 targets. J. Nutr. Biochem. 2022, 101, 108929. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ding, J.; Zhang, H.; Shen, J.; Hao, Y.; Zhang, X. Lactobacillus casei LH23 modulates the immune response and ameliorates DSS-induced colitis via suppressing JNK/p-38 signal pathways and enhancing histone H3K9 acetylation. Food Funct. 2020, 11, 5473–5485. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zuo, L.; Zhu, W.; Gong, J.; Zhang, W.; Guo, Z. Telmisartan attenuates the inflamed mesenteric adipose tissue in spontaneous colitis by mechanisms involving regulation of neurotensin/microRNA-155 pathway. Biochem. Pharmacol. 2015, 93, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Caradonna, F.; Cruciata, I.; Lauria, A.; Perrone, A.; Gentile, C. Melatonin reduces inflammatory response in human intestinal epithelial cells stimulated by interleukin-1β. J. Pineal. Res. 2019, 67, e12598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, Y.; Meng, J.; Girotra, M.; Ramakrishnan, S.; Roy, S. Immune modulation mediated by extracellular vesicles of intestinal organoids is disrupted by opioids. Mucosal Immunol. 2021, 14, 887–898. [Google Scholar] [CrossRef]
- Yang, Z.B.; Qiu, L.Z.; Chen, Q.; Lin, J.D. Artesunate alleviates the inflammatory response of ulcerative colitis by regulating the expression of miR-155. Pharm. Biol. 2021, 59, 97–105. [Google Scholar] [CrossRef]
- Felice, C.; Lewis, A.; Iqbal, S.; Gordon, H.; Rigoni, A.; Colombo, M.P. Intestinal Inflammation is Linked to Hypoacetylation of Histone 3 Lysine 27 and can be Reversed by Valproic Acid Treatment in Inflammatory Bowel Disease Patients. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 889–891.e6. [Google Scholar] [CrossRef]
- Chu, Y.; Zhu, Y.; Zhang, Y.; Liu, X.; Guo, Y.; Chang, L. Tetrandrine attenuates intestinal epithelial barrier defects caused by colitis through promoting the expression of Occludin via the AhR-miR-429 pathway. FASEB J. 2021, 35, e21502. [Google Scholar] [CrossRef]
- Gubatan, J.; Chou, N.D.; Nielsen, O.H.; Moss, A.C. Systematic review with meta-analysis: Association of vitamin D status with clinical outcomes in adult patients with inflammatory bowel disease. Aliment. Pharmacol. Ther. 2019, 50, 1146–1158. [Google Scholar] [CrossRef]
- McGillis, L.; Bronte-Tinkew, D.M.; Dang, F.; Capurro, M.; Prashar, A.; Ricciuto, A. Vitamin D deficiency enhances expression of autophagy-regulating miR-142-3p in mouse and “involved” IBD patient intestinal tissues. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G171–G184. [Google Scholar] [CrossRef]
- Liu, F.-H.; Li, S.-S.; Li, X.-X.; Wang, S.; Li, M.-G.; Guan, L.; Luan, T.-G.; Liu, Z.-G.; Liu, Z.-J.; Yang, P.-C. Vitamin D3 induces vitamin D receptor and HDAC11 binding to relieve the promoter of the tight junction proteins. Oncotarget 2017, 8, 58781–58789. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhu, Y.; Li, C.; Yu, J.; Ren, D.D.; Qiu, S. 1,25-Dihydroxyvitamin D regulates macrophage polarization and ameliorates experimental inflammatory bowel disease by suppressing miR-125b. Int. Immunopharmacol. 2019, 67, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Barrón-Cabrera, E.; Ramos-Lopez, O.; González-Becerra, K.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez-López, E. Epigenetic Modifications as Outcomes of Exercise Interventions Related to Specific Metabolic Alterations: A Systematic Review. Lifestyle Genom. 2019, 12, 25–44. [Google Scholar] [CrossRef]
- White, A.J.; Sandler, D.P.; Bolick, S.C.E.; Xu, Z.; Taylor, J.A.; Deroo, L.A. Recreational and household physical activity at different time points and DNA global methylation. Eur. J. Cancer 2013, 49, 2199–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchis-Gomar, F.; Garcia-Gimenez, J.L.; Perez-Quilis, C.; Gomez-Cabrera, M.C.; Pallardo, F.V.; Lippi, G. Physical exercise as an epigenetic modulator: Eustress, the “positive stress” as an effector of gene expression. J. Strength Cond. Res. 2012, 26, 3469–3472. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Åkerström, T.C.A.; Nielsen, A.R.; Fischer, C.P. Role of myokines in exercise and metabolism. J. Appl. Physiol. 2007, 103, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Tarnowski, M.; Kopytko, P.; Piotrowska, K. Epigenetic regulation of inflammatory responses in the context of physical activity. Genes 2021, 12, 1313. [Google Scholar] [CrossRef]
- Wang, J.; Liu, S.; Li, G.; Xiao, J. Exercise Regulates the Immune System. Adv. Exp. Med. Biol. 2020, 1228, 395–408. [Google Scholar] [CrossRef]
- Machado, O.A.S.; Diniz, V.L.S.; Passos, M.E.P.; de Oliveira, H.H.; Santos-Oliveira, L.C.; Alecrim, A.L. Physical exercise increases global and gene-specific (interleukin-17 and interferon-γ) DNA methylation in lymphocytes from aged women. Exp. Physiol. 2021, 106, 1878–1885. [Google Scholar] [CrossRef]
- Dorneles, G.P.; Boeira, M.C.R.; Schipper, L.L.; Silva, I.R.V.; Elsner, V.R.; Dal Lago, P. Acute Strenuous Exercise Induces an Imbalance on Histone H4 Acetylation/Histone Deacetylase 2 and Increases the Proinflammatory Profile of PBMC of Obese Individuals. Oxid. Med. Cell Longev. 2017, 2017, 1530230. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.P.; Bhalla, D.K.; Whayne, T.F.; Gairola, C.G. Cigarette smoke and adverse health effects: An overview of research trends and future needs. Int. J. Angiol. 2007, 16, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Terzikhan, N.; Verhamme, K.M.C.; Hofman, A.; Stricker, B.H.; Brusselle, G.G.; Lahousse, L. Prevalence and incidence of COPD in smokers and non-smokers: The Rotterdam Study. Eur. J. Epidemiol. 2016, 31, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Marcin, W.; Michael, S. Genetics and epigenetics of inflammatory bowel disease. Swiss Med. Wkly. 2018, 148, w14671. [Google Scholar] [CrossRef]
- Wallace, K.; Grau, M.V.; Levine, A.J.; Shen, L.; Hamdan, R.; Chen, X. Association between folate levels and CpG island hypermethylation in normal colorectal mucosa. Cancer Prev. Res. 2010, 3, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Ananthakrishnan, A.N.; Nguyen, D.D.; Sauk, J.; Yajnik, V.; Xavier, R.J. Genetic polymorphisms in metabolizing enzymes modifying the association between smoking and inflammatory bowel diseases. Inflamm. Bowel Dis. 2014, 20, 783–789. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Szakal, D.; Ross, M.C.; Dowd, S.E.; Mir, S.A.V.; Schaible, T.D.; Petrosino, J.F. Maternal micronutrients can modify colonic mucosal microbiota maturation in murine offspring. Gut Microbes 2012, 3, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Nakahigashi, M.; Saniabadi, A.R. Review article: Diet and inflammatory bowel disease-Epidemiology and treatment. Aliment. Pharmacol. Ther. 2009, 30, 99–112. [Google Scholar] [CrossRef]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Ananthakrishnan, A.N. The exposome in inflammatory bowel disease. Trop. Gastroenterol. 2014, 35, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawan, C.; Vaissière, T.; Murr, R.; Herceg, Z. Epigenetic drivers and genetic passengers on the road to cancer. Mutat. Res. 2008, 642, 1–13. [Google Scholar] [CrossRef] [PubMed]

| Metabolite | Activity | Epigenetic Mechanism | Tissue/Cells | Mechanism | Model | Author |
|---|---|---|---|---|---|---|
| SCFAs | ||||||
| SCFAs | Anti-inflammatory activity | HDACs inhibition | T cells | Inhibits HDACs in T cells and increases the acetylation of p70 S6 kinase and phosphorylation rS6, regulating the mTOR pathway required for generation of Th17 (T helper type 17), Th1, and IL-10(+) T cells | C57BL/6 mice; CD4+ T cells isolated from the spleen and lymph nodes | Park J, et al. (2015) [154] |
| SCFAs | Anti-inflammatory activity | HDACs inhibition | B cells | Upregulates regulatory B cells capable of producing IL-10 in a manner dependent on their HDAC inhibitory activity | DSS-induced colitis in mice | Zou F, et al. (2021) [155] |
| SCFAs | Anti-inflammatory activity | miR-145 | IECs | Decreases the CEBPB expression, which could bind to the miR-145 promoter to inhibit its expression, thereby promoting the expression of DUSP6 (dual-specificity phosphatase 6) and thus prevents the development of intestinal inflammation | LPS-treated intestinal epithelial cells | Liu Q, et al. (2022) [156] |
| Butyrate | ||||||
| Butyrate | Anti-inflammatory activity | Histone acetylation | IECs | Butyrate, by inducing an increase in histone acetylation in the NOD2 promoter region, induces NOD2 upregulation, and impact the defence mechanism against the bacterial membrane component peptidoglycan by inducing IL-8 and GRO-alpha secretion | Caco-2 cell line | Leung CH, et al. (2009) [147] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | Dendritic cells | Butyrate has a role of HDACi on the epigenetic modification of gene expression, inhibits IL-12 and upregulates subunit IL-23p19 | DSS-induced colitis in mice | Berndt BE, et al. (2012) [153] |
| Butyrate | Anti-inflammatory activity | HDAC1 inhibition | T cells | Butyrate inhibits HDAC1 activity to induce Fas promoter hyperacetylation and Fas upregulation in T cells and promote Fas-mediated apoptosis of T cells to eliminate the source of inflammation | BALB/c mice | Zimmerman MA, et al. (2012) [157] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | IECs | Butyrate may contribute to the restoration of the tight junction barrier in IBD by affecting the expression of claudin-2, occludin, cingulin, and zonula occludens proteins (ZO-1, ZO-2) via inhibition of histone deacetylase | DSS-induced colitis in mice | Plöger S, et al. (2012) [146] |
| Butyrate | Anti-inflammatory activity | Histone H3 acetylation | T cells | Butyrate enhances histone H3 acetylation in the promoter and conserves non-coding sequence regions of the Foxp3 locus, regulating the differentiation of Treg cells, ameliorating colitis | Germ-free and CRB-associated mice; OT-II (Ly5.2) transgenic CD4+ T cells | Furusawa Y, et al. (2013) [144] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | Macrophages | Butyrate reduces de production of proinflammatory mediators by macrophages including nitric oxide, IL-6, and IL-12, but did not affect levels of TNF-α or MCP-1 | DSS-induced colitis in mice | Chang PV, et al. (2014) [149] |
| Butyrate | Anti-inflammatory activity | H3K9 acetylation | Macrophages | Butyrate activates STAT6-mediated transcription through H3K9 acetylation driving M2 macrophage polarization | DSS-induced colitis in mice | Ji J, et al. (2016) [150] |
| Butyrate | Anti-inflammatory activity | Histone H3 acetylation | Macrophages | Oral supplementation with butyrate attenuates experimental murine colitis by blocking NF-κB signaling and reverses histone acetylation | DSS-induced colitis in mice, IL-10−/− mice and RAW264.7 cells | Lee C, et al. (2017) [158] |
| Butyrate | Anti or proinflammatory activity depending on its concentration and immunological milieu | HDACs inhibition | T cells | Lower butyrate concentrations facilitates differentiation of Tregs in vitro and in vivo under steady-state conditions. In contrast, higher concentrations of butyrate induces expression of the transcription factor T-bet in all investigated T cell subsets resulting in IFN-γ-producing Tregs or conventional T cells. This effect was mediated by the inhibition of histone deacetylase activity | DSS-induced colitis in mice; CD4+ T cells | Kespohl M, et al. (2017) [159] |
| Butyrate | Anti-inflammatory activity | HDAC3 inhibition | MonocyteMacrophage | Butyrate induces the monocyte to macrophage differentiation and promotes its antimicrobial activity and restricts bacterial translocation, through HDAC3 inhibition | Human monocytes isolated from leukocyte cones of healthy blood donors | Schulthess J, et al. (2019) [148] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | T cells | Butyrate promotes Th1 cell development by promoting IFN-γ and T-bet expression and inhibits Th17 cell development by suppressing IL-17, Rorα, and Rorγt expression and upregulate IL-10 production in Th1 and Th17 | CBir1 transgenic T cells; Rag1−/− mice | Chen L, et al. (2019) [151] |
| Butyrate | Anti-inflammatory activity | Increase of histone acetylation | IECs | Butyrate induces HSF2 (Heat-shock transcription factor 2) expression epigenetically via increasing histone acetylation levels at the promoter region, enhancing autophagy in IECs | UC (n = 50) and healthy (n = 30) patients; DSS-induced colitis in mice; HT-29 cells | Zhang F, et al. (2020) [160] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | IECs | Butyrate induces SYNPO (Synaptopodin) in epithelial cell lines through mechanisms possibly involving histone deacetylase inhibition. SYNPO contributes by intestinal homeostasis by controlling intestinal permeability | Epithelial cell lines; DSS-induced colitis in mice | Wang RX, et al. (2020) [161] |
| Butyrate | Anti-inflammatory activity | HDAC inhibition | Neutrophils | Butyrate significantly inhibits IBD neutrophils to produce proinflammatory cytokines, chemokines, and calprotectins through HDAC inhibition | Peripheral neutrophils isolated from IBD patients and healthy donors; DSS-induced colitis in mice | Li G, et al. (2021) [152] |
| Propionate | Anti-inflammatory activity | HDAC1 inhibition | IECs | Propionate promotes intestinal epithelial cell migration by enhancing cell spreading and polarization, a function dependant of the inhibition of class I HDAC | Mouse small intestinal epithelial cells (MSIE) and human Caco-2 cells; DSS-induced colitis in mice | Bilotta AJ, et al. (2021) [142] |
| Caprylic acid (C8) and nonanoic acid (C9) (medium chain fatty acids) | Anti-inflammatory activity | Acetylation of histone 3 lysine 9 (H3K9) | IECs | Reduces bacterial translocation, enhances antibacterial activity, and attenuates the activity of the classical histone deacetylase pathway to facilitate the acetylation of histone 3 lysine 9 (H3K9) at the promoters pBD-1 and pBD-2, remarkably increases the secretion of porcine β-defensins 1 (pBD-1) and pBD-2 | Porcine jejunal epithelial cell line-J2 | Wang J, et al. (2018) [162] |
| Drug | Activity | Epigenetic Mechanism | Tissue/Cells | Mechanism | Model | Author |
|---|---|---|---|---|---|---|
| IBD medication | ||||||
| Mesalamine | Anti-inflammatory activity | miR-206 | IECs and colonic tissues | Long-term treatment donw-regulates miR-206 which confer a protective effect in inducing and maintaining histologic remission | HT29 colon cells; UC patients (n = 10) | Minacapelli CD, et al. (2019) [258] |
| 5-ASA | Anti-inflammatory activity | miR-125b, miR-150, miR-155, miR-346 and miR-506 | IECs | 5-ASA suppressed the levels of miR-125b, miR-150, miR-155, miR-346 and miR-506 in IECs and inhibition of these miR were associated with significant inductions of their target genes such as vitamin D receptor (VDR), suppressor of cytokine signaling (SOCS1), Forkhead box O (FOXO3a) and DNA methyltransferase 1 (DNMT1) | Caco-2 cells | Adamowicz M, et al. (2021) [259] |
| Infliximab | Anti-inflammatory activity | miR-10a | DCs | Anti-TNF mAb treatment significantly promote miR-10a expression, whereas it markedly inhibited NOD2 and IL-12/IL-23p40 in the inflamed mucosa | Human monocyte-derived dendritic cells (DC); IBD patients | Wu W, et al. (2015) [127] |
| Infliximab | Anti-inflammatory activity | miR-301a | T cells | Decreases miR-301a expression in IBD CD4+ T cells by decreasing Th17 cell differentiation through upregulation of SNIP1 | Peripheral blood mononuclear cells (PBMC); inflamed mucosa of patients with IBD | He C, et al. (2016) [260] |
| Infliximab | Anti-inflammatory activity | lnc-ITSN1-2 | T cells | Lnc-ITSN1-2 promotes IBD CD4+ T cell activation, proliferation, and Th1/Th17 cell differentiation by serving as a competing endogenous RNA for IL-23R via sponging miR-125a | Intestinal mucosa from IBD patients (n = 6) and healthy controls (n = 6) | Nie J, et al. (2020) [261] |
| Infliximab | Anti-inflammatory activity | miR-30 family | IECs | Decreases circRNA_103765 expression, which act as a molecular sponge to adsorb the miR-30 family and impair the negative regulation of Delta-like ligand 4 (DLL4) and protect human IECs from TNF-α-induced apoptosis | IBD patients; PBMCs | Ye Y, et al. (2021) [262] |
| Infliximab | Anti-inflammatory activity | miR-146a and miR-146b | Serum and intestinal mucosae | Decreases miR-146a and miR-146b levels in serum. miR-146a probably promotes colitis through TLR4/MyD88/NF-κB signaling pathway | Serum of 19 IBD patients | Batra SK, et al. (2020) [263] |
| Infliximab (IFX) therapy and longer-term steroids (weeks) | Anti-inflammatory activity | miR-320a | Decreases miR-320a serum level. miR-320a could play a role in sensitization of the quiescent mucosa to environmental factors | Serum of 19 IBD patients | ||
| Anti-TNF and glucocorticoids | Anti-inflammatory activity | let-7c | let-7c serum level decreases, thus reduces M2 macrophage polarization (anti-inflammatory) and promote M1 (proinflammatory) polarization | Serum of 19 IBD patients | ||
| Anti-TNF | Anti-inflammatory activity | miR-10a | DCs | Blockade TNF with anti-TNF mAb markedly enhances miR10a expression in the intestinal mucosa. miR-10a could block intestinal inflammation and reduce the differentiation Th1 and Th17 | C57BL/6 (B6) mice | Xue X, et al. (2011) [125] |
| Anti-TNF | Anti-inflammatory activity | miR-378a-3p, miR-378c | Colonic mucosae | Increases levels of miR-378a-3p and miR-378c. Over-expression of miR-378a-3p decreased the levels of an IL-33 target sequence β-gal-reporter gene | Active UC patients (n = 24); inactive UC (n = 10); controls (n = 6); HEK293 cells | Dubois-Camacho K, et al. (2019) [264] |
| Enemas containing short chain fatty acids (SCFA) such as butyrate, propionate, and acetate | Anti-inflammatory activity | Histone acetylation | IECs | SCFAs increase histone acetylation states and inhibit the production of proinflammatory substances, such as IL-8, by the intestinal epithelium | Caco-2 cells | Huang N, et al. (1997) [288] |
| N-(1-carbamoyl-2-phenylethyl) butyramide (FBA), a butyrate-releasing derivative | Anti-inflammatory activity | Histone deacetylase-9 and H3 histone acetylation | Colonic mucosae | FBA, similar to its parental compound sodium butyrate, inhibited histone deacetylase-9 and restored H3 histone acetylation, exerting an anti-inflammatory effect through NF-κB inhibition and the upregulation of PPARγ | DSS-induced colitis in mice | Simeoli R, et al. (2017) [289] |
| Exclusive enteral nutrition (EEN) | Anti-inflammatory activity | hsa-miR-192-5p, hsa-miR-423-3p, hsa-miR-99a-5p, hsa-miR-124-3p, hsa-miR-301a-5p, hsa-miR-495-5p, and hsa-let-7b-5p | Intestinal mucosae | EEN induces mucosal miRNAs expression profile (altered expressions of hsa-miR-192-5p, hsa-miR-423-3p, hsa-miR-99a-5p, hsa-miR-124-3p, hsa-miR-301a-5p, hsa-miR-495-5p, and hsa-let-7b-5p) after EEN therapy was significantly changed compared with inflamed mucosa before treatment | CD patients (n = 30) | Guo Z, et al. (2016) [265] |
| ABX464 | Anti-inflammatory activity | miR-124 | Immune cells | Upregulates miR-124 in human immune cells, which is a negative regulator of inflammation and was shown to target RNAs, such as STAT and TLR | Tazi J, et al. (2021) [290] | |
| MSCs | Anti-inflammatory activity | miR-181a | IECs | MSC-derived exosomal miR-181a could alleviate colitis by promoting intestinal barrier function decreased (increasing level of Claudin-1, ZO-1, and IκB) | DSS-induced colitis in mice and induced human colonic epithelial cell (HCOEPIC) | Gu L, et al. (2021) [266] |
| MSCs | Anti-inflammatory activity | H3K27me3 | T cells | Extracellular vesicles from MSCs could inhibit the differentiation of Th17 cells by regulating H3K27me3 | TNBS-induced colitis in mice | Chen Q, et al. (2020) [291] |
| IFN-γ pretreated bone marrow mesenchymal stem cells | Anti-inflammatory activity | miR-125a and miR-125b | T cells | Increases the level of miR-125a and miR-125b of exosomes, which directly targeted on Stat3, to repress Th17 cell differentiation | DSS-induced colitis in mice | Yang R, et al. (2020) [267] |
| Vascular endothelial growth factor-C-treated adipose-derived stem cells (ADSCs) | Anti-inflammatory activity | miR-132 | Lymphatic endothelial cells | VEGF-C-treated ADSCs have a higher level of miR-132, which promotes lymphangiogenic response by directly targeting Smad-7 and regulating TGF-β/Smad signaling | Lymphatic endothelial cells (LECs) | Wang X, et al. (2018) [292] |
| Supplementation | ||||||
| Iron | Proinflammatory activity | TET1 induction; NRF2, NQ01, GPX2 demethylation | IECs and intestinal mucosae | Chronic iron exposure leads to induction of TET1 expression leading to demethylation of NRF2 (nuclear factor erythroid 2-related factor 2) pathway targets (including NAD(P)H Quinone Dehydrogenase 1 (NQO1) and Glutathione peroxidase 2 (GPX2). NQO1 and GPX2 hypomethylation led to increased gene and protein expression, and could be a route by which cells overcome persistent and chronic oxidative stress | Caco-2 cells and wild-type C57BL/6 mice | Horniblow RD, et al. (2022) [293] |
| Antibiotics | ||||||
| Isotretinoin | Anti-inflammatory activity | miR | T cells | 3 miR overexpressed in naive T-cells and potentially downregulate 777 miR targets (cytoskeleton remodelling and the c-Jun N terminal kinase (JNK) signaling pathway) | Balb/c mice | Becker E, et al. (2016) [268] |
| Metronidazole | Anti-inflammatory activity | miR | 5 miR were significantly lower in naive T-cells resulting in the prediction of 340 potentially upregulated miR targets associated with IL-2 activation and signaling, cytoskeleton remodelling and epithelial-to-mesenchymal-transition (EMT). | |||
| Doxycycline | Anti-inflammatory activity | miR-144-3p | Overexpression of miR-144-3p that resulted in the prediction of 493 potentially downregulated miR targets involved in protein kinase A (PKA), protein kinase B and nuclear factor of activated T-cells (NFAT) signaling pathways | |||
| Tetracyclines | Anti-inflammatory activity | miR-150, miR-155, miR-375 and miR-146 | Colonic tissues | Reduce miR-150 and miR-155 expression, upregulate miR-375 and miR-142 | DSS-induced colitis in mice and bone marrow-derived macrophages | Garrido-Mesa J, et al. (2018) [269] |
| Antibiotics treatment | Anti-inflammatory activity | DNA demethylation | IECs | Suppresses aberrant DNA methylation of three marker CpG islands (Cbln4, Fosb, and Msx1) induced by chronic inflammation | AOM/DSS-induced colitis in mice | Hattori N, et al. (2019) [270] |
| Probiotics | ||||||
| Probiotic bacterium Escherichia coli Nissle 1917 (EcN) | Anti-inflammatory activity | miR-203, miR-483-3p, miR-595 | IECs | Increases miR-203, miR-483-3p, miR-595 targeting tight junction (TJ) proteins; these miRNAs are involved in the regulation of barrier function by modulating the expression of regulatory and structural components of tight junctional complexes. | T84 cells | Veltman K, et al. (2012) [271] |
| Bifidobacterium longum | Anti-inflammatory activity | DNA demethylation | Peripheral blood mononuclear cells | B. Longum treatment significantly demethylates several CpG sites in Foxp3 promoter | TNBS-induced colitis in rat; spleen peripheral blood mononuclear cells (PBMC) cells was extracted | Zhang M, et al. (2017) [272] |
| Lactobacillus fermentum and Lactobacillus salivarius | Anti-inflammatory activity | miR-155, miR-223, miR-150 and miR-143 | Colonic tissues | They increase the expression of miR-155 and miR-223, and miR-150 and miR-143 for L. fermentum, involved in the immune response (restoration of Treg cell population and the Th1/Th2 cytokine balance) and in the intestinal barrier function | C57BL/6J mice | Rodríguez-Nogales A, et al. (2017) [273] |
| Saccharomyces boulardii | Anti-inflammatory activity | miR-155 and miR-223; miR-143 and miR-375 | Colonic tissuess | Increasing the expression of miR-155 and miR-223, whereas decreasing the expression miR-143 and miR-375 | DSS-induced colitis in mice | Rodríguez-Nogales A, et al. (2018) [274] |
| Bifidobacterium bifidum ATCC 29521 | Anti-inflammatory activity | miR-150, miR-155, miR-223 | Colonic mucosae | Restorates miR-150, miR-155, miR-223, upregulates anti-inflammatory cytokines (IL-10, PPARγ, IL-6), tight junction proteins (such as ZO-1, MUC-2, Claudin-3, and E Cadherin-1) and downregulates inflammatory genes (TNF-α, IL-1β) | DSS-induced colitis in mice | Din AU, et al. (2020) [275] |
| Lactobacillus casei LH23 probiotic | Anti-inflammatory activity | Histone H3K9 acetylation | Colonic tissues | Modulates the immune response and ameliorates colitis via suppressing JNK/p-38 signal pathways and enhancing histone H3K9 acetylation | DSS-induced colitis in mice; LPS-induced RAW264.7 cells | Liu M, et al. (2020) [294] |
| Lactic Acid-Producing Probiotic Saccharomyces cerevisiae | Anti-inflammatory activity | Histone H3K9 acetylation and histone H3K18 lactylation | Macrophages | Promotes histone H3K9 acetylation and histone H3K18 lactylation and attenuates intestinal inflammation via suppressing macrophage pyroptosis | DSS-induced colitis in mice | Sun S, et al. (2021) [276] |
| Other medication | ||||||
| Telmisartan (angiotensin II type 1 receptor blocker and a peroxisome proliferator-activated receptor-receptor-γ agonist) | Anti-inflammatory activity | miR-155 | Mesenteric adipocytes | Restorates the mesenteric adipose tissue adipocyte morphology and the expression of adipokines by suppressing the neurotensin/miR-155 pathway | IL-10(−)/(−) mice; cultured mesenteric adipose tissue from Crohn’s disease patients | Li Y, et al. (2015) [295] |
| Melatonine | Anti-inflammatory activity | Prevent DNA methylation | IECs | Prevents DNA demethylation, reduces NF-κB activation, decreases the levels of inflammatory mediators (including IL-6, IL-8, COX-2, and NO), and reduces increase in paracellular permeability, attenuating the inflammatory response | Caco-2 cells | Mannino G, et al. (2019) [296] |
| Morphine | Proinflammatory activity | Let7c-5p | Macrophages, DCs | Opioid treatment can disrupt gut immune homeostasis by inhibiting packaging of miR into EVs secreted by intestinal crypt cells (with a decreased amount of Let7c-5p) | C57BL/6J mice; organoid culture | Zhang Y, et al. (2021) [297] |
| Artesunate | Anti-inflammatory activity | miR-155 | Macrophages | Inhibits the expression of miR-155 to inhibit the NF-κB pathway | LPS-induced RAW264.7 cells; BALB/c mice model | Yang ZB, et al. (2021) [298] |
| Valproic acid treatment | Anti-inflammatory activity | HDAC inhibition | Intestinal tissue | Inhibits HDAC activity and increases H3K27ac levels and reduced expression of IL6, IL10, IL1B, and IL23 | DSS-induced colitis in mice | Felice C, et al. (2021) [299] |
| Tetrandrine | Anti-inflammatory activity | miR-429 | IECs | Tetrandrine can attenuate the intestinal epithelial barrier defects in colitis through promoting occludin expression via the AhR/miR-429 pathway | DSS-induced colitis in mice | Chu Y, et al. (2021) [300] |
| Chinese medicine | ||||||
| Sinomenine, a pure alkaloid isolated in Chinese medicine | Anti-inflammatory activity | miR-155 | Colonic tissues | Downregulates the levels of miR-155 and several related inflammatory cytokines | TNBS-induced colitis in mice | Yu Q, et al. (2013) [277] |
| Tripterygium wilfordii Hook F (TWHF) | Anti-inflammatory activity | miR-155 | Ileocolonic anastomosis | Triptolide could suppress miR-155/SHIP-1 signaling pathway and attenuated expression of inflammatory cytokines after ileocaecal resection | IL-10(−/−) mice | Wu R, et al. (2013) [278] |
| Herb-partitioned moxibustion (HPM) | Anti-inflammatory activity | miR-147 and miR-205 | Colonic tissues | Upregulates the expression of miR-147 and miR-205 and then further regulate some of their target genes, thereby indirectly inhibiting the inflammatory signal pathways mediated by TLR, NF-κB, and so forth and decreasing the production of downstream inflammatory cytokines such as TNF-α and IL-1β, so as to alleviate intestinal inflammation in CD | Experimental CD rat models | Wei K, et al. (2015) [280] |
| Salvianolic acid B (Sal B) is isolated from the traditional Chinese medical herb Salvia miltiorrhiza | Anti-inflammatory activity | miR-1 | IECs | Sal B restores barrier function by miR-1 activation and subsequent myosin light chain kinase (MLCK) inactivation | TNBS-induced rat colitis model | Xiong Y, et al. (2016) [281] |
| Herb-partitioned moxibustion (HPM) | Anti-inflammatory activity | miR-184 and miR-490-5p | Colonic tissue | HPM regulates miR-184 and miR-490-5p expression, act on the transcription of their target genes to regulate inflammatory signaling pathways, and attenuate inflammation and tissue injury in the colons of rats with DSS-induced UC | DSS-induced colitis in mice | Huang Y, et al. (2017) [282] |
| Triptolide (TPL) | Anti-inflammatory activity | miR-16-1 | Ileocolonic anastomosis | TPL reduces miR-16-1 levels aggravating anastomotic inflammation and fibrosis | IL-10−/− mice | Hou HW, et al. (2017) [283] |
| Norisoboldine (NOR), a natural aryl hydrocarbon receptor (AhR) | Anti-inflammatory activity | H3K9me3 modification | T cells | NOR promoted Treg differentiation and then alleviated the development of colitis by regulating AhR (aryl hydrocarbon receptor)/glycolysis axis and decreases NAD+ and SIRT1 (sirtuin 1), facilitates the ubiquitin-proteasomal degradation of SUV39H1, which is a major member of histone KMTs and catalyses the H3K9me3 modification, which is associated with transcription repression of Foxp3 | Lv Q, et al. (2018) [284] | |
| Triptolide (TPL), the most potent bioactive substance in TWHF (Tripterygium wilfordii Hook F) extract | Anti-inflammatory activity | miR-16-1 | Fibroblasts | Inhibits migration and proliferation of fibroblasts from ileocolonic anastomosis of CD patients via regulating the miR-16-1/HSP70 pathway | Fibroblasts from strictured anastomosis tissue (SAT) samples and matched anastomosis-adjacent normal tissue (NT) samples of CD patients (n = 10) | Chen M, et al. (2019) [285] |
| Polysaccharide RAMPtp from Atractylodis macrocephalae Koidz | Anti-inflammatory activity | lncRNA ITSN1-OT1 | IECs | Induces lncRNA ITSN1-OT1, which blocks the nuclear import of phosphorylated STAT2 and prevents the decrease of expression and structural destroy of tight junction proteins | DSS-induced colitis in mice | Zong X, et al. (2021) [286] |
| Dendrobium officinale polysaccharide (DOP) | Anti-inflammatory activity | miR-433-3p | IECs, macrophages | DOP interfered with the secretion of small extracellular vesicles (DIEs) by IEC, with increased miR-433-3p expression. When delivered to macrophages, miR-433-3p targeted the MAPK8 gene, leading to inhibition of the MAPK signaling pathway and reduced production of inflammatory cytokines | IECs, macrophages | Liu H, et al. (2021) [287] |
| Huangqin-Tang decoction (HQT) | Anti-inflammatory activity | miR-185-3p | IECs | HQT could upregulate miR-185-3p, thereby affecting the myosin light chain kinase (MLCK)/myosin light chain phosphorylation (p-MLC) pathway and leading to increased expression of occludin protein, which ultimately protected the intestinal epithelial barrier function | Balb/c mice | Changlin Z, et al. (2021) [279] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieujean, S.; Caron, B.; Haghnejad, V.; Jouzeau, J.-Y.; Netter, P.; Heba, A.-C.; Ndiaye, N.C.; Moulin, D.; Barreto, G.; Danese, S.; et al. Impact of the Exposome on the Epigenome in Inflammatory Bowel Disease Patients and Animal Models. Int. J. Mol. Sci. 2022, 23, 7611. https://doi.org/10.3390/ijms23147611
Vieujean S, Caron B, Haghnejad V, Jouzeau J-Y, Netter P, Heba A-C, Ndiaye NC, Moulin D, Barreto G, Danese S, et al. Impact of the Exposome on the Epigenome in Inflammatory Bowel Disease Patients and Animal Models. International Journal of Molecular Sciences. 2022; 23(14):7611. https://doi.org/10.3390/ijms23147611
Chicago/Turabian StyleVieujean, Sophie, Bénédicte Caron, Vincent Haghnejad, Jean-Yves Jouzeau, Patrick Netter, Anne-Charlotte Heba, Ndeye Coumba Ndiaye, David Moulin, Guillermo Barreto, Silvio Danese, and et al. 2022. "Impact of the Exposome on the Epigenome in Inflammatory Bowel Disease Patients and Animal Models" International Journal of Molecular Sciences 23, no. 14: 7611. https://doi.org/10.3390/ijms23147611