
Regulation of VEGFA, KRAS, and NFE2L2 Oncogenes by MicroRNAs in Head and Neck Cancer

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
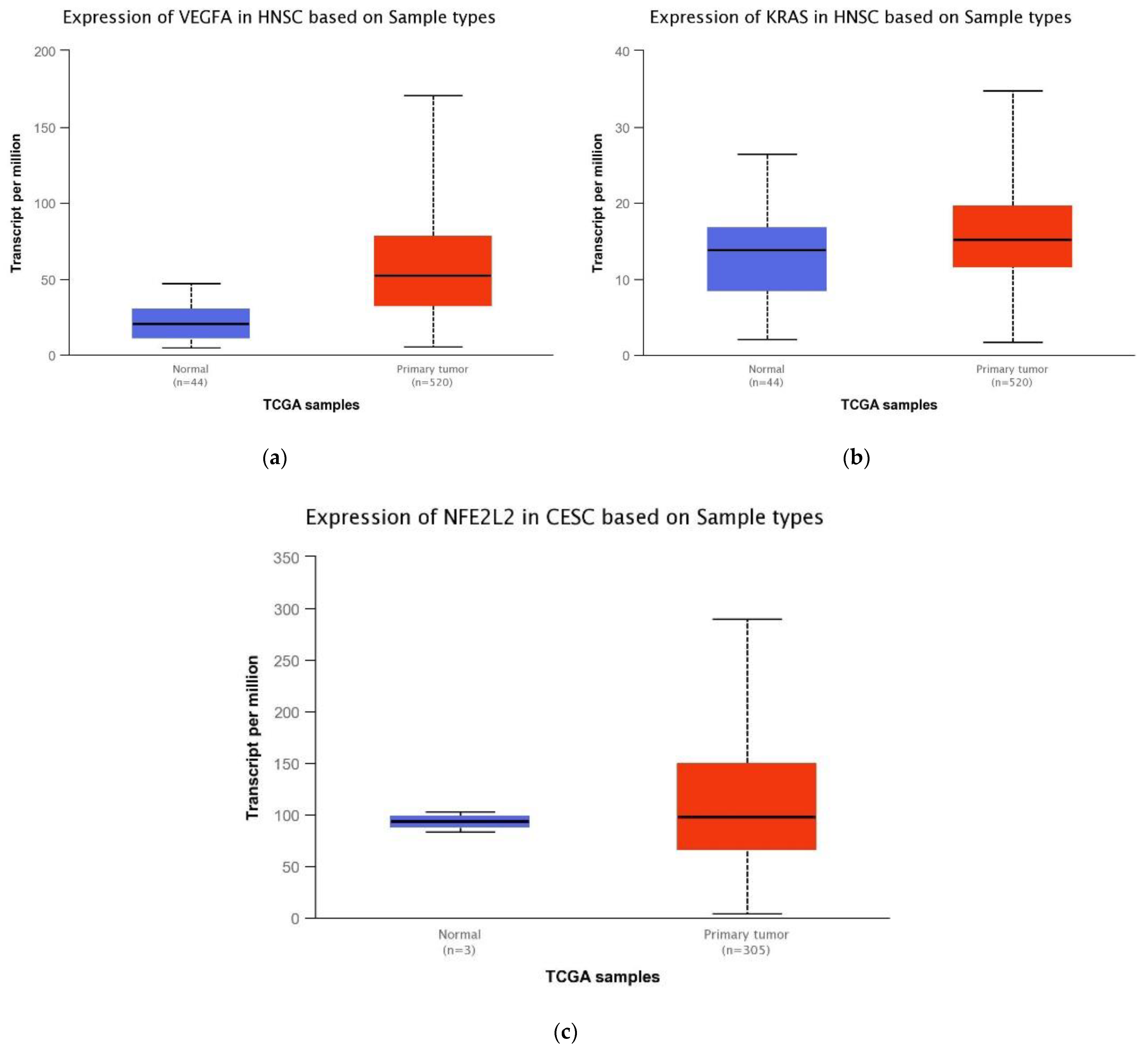
2.1. Bioinformatics-TCGA Analysis
2.2. Expression of miR-17-5p, miR-140-5p, and miR-874-3p in HNC Cell Lines
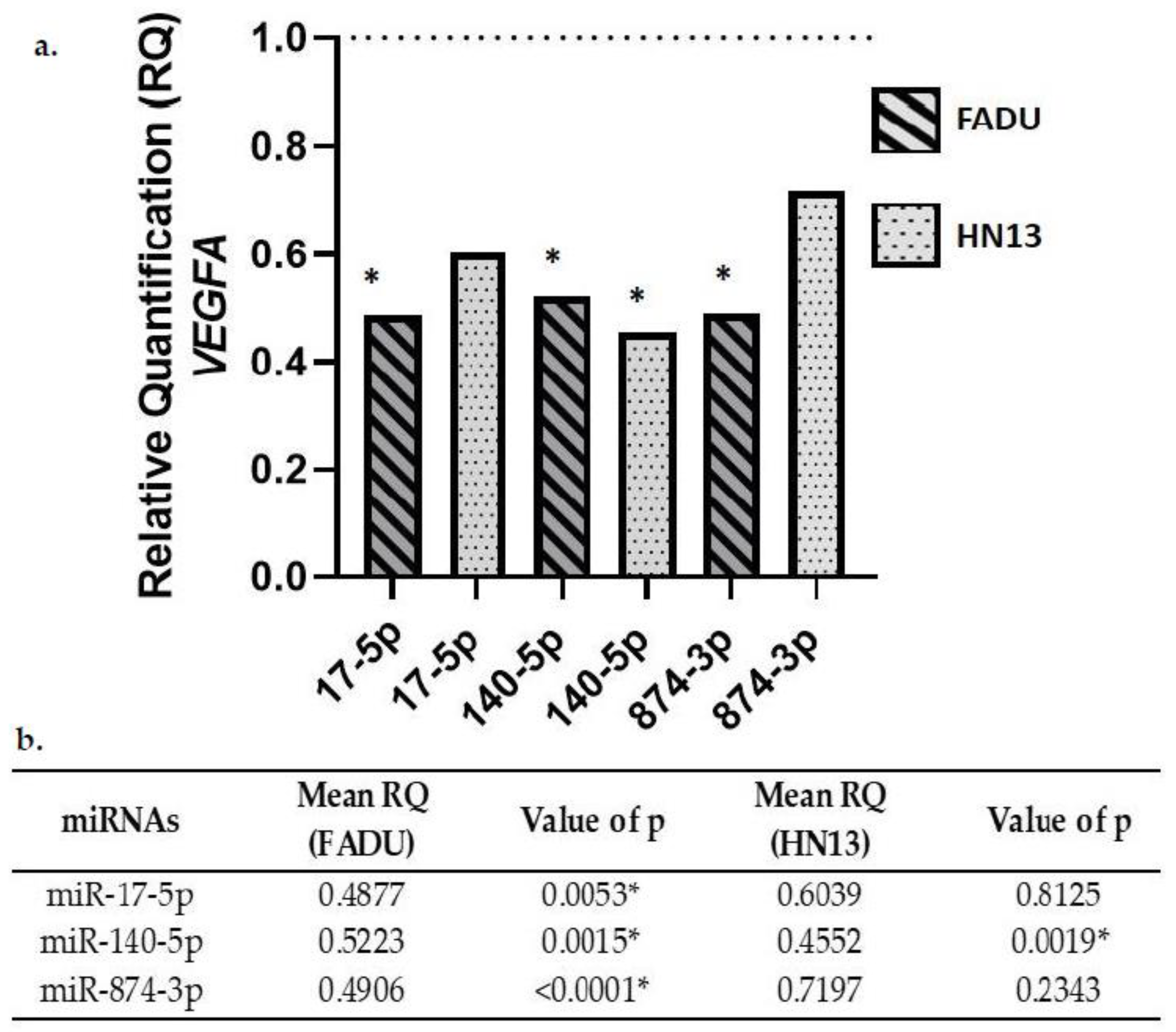
2.3. Gene Expression of VEGFA in Transfected Cells
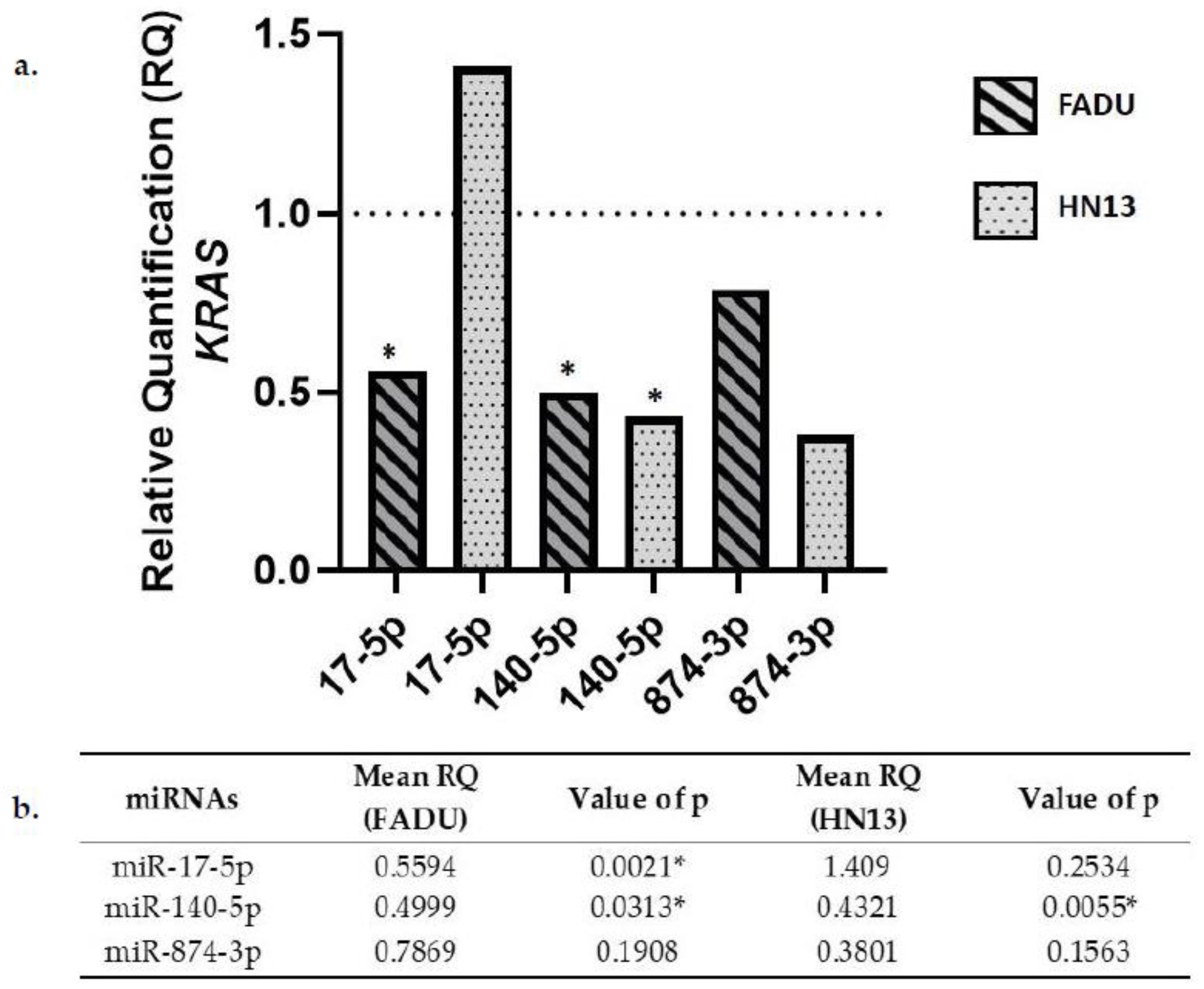
2.4. KRAS Gene Expression in Transfected Cells
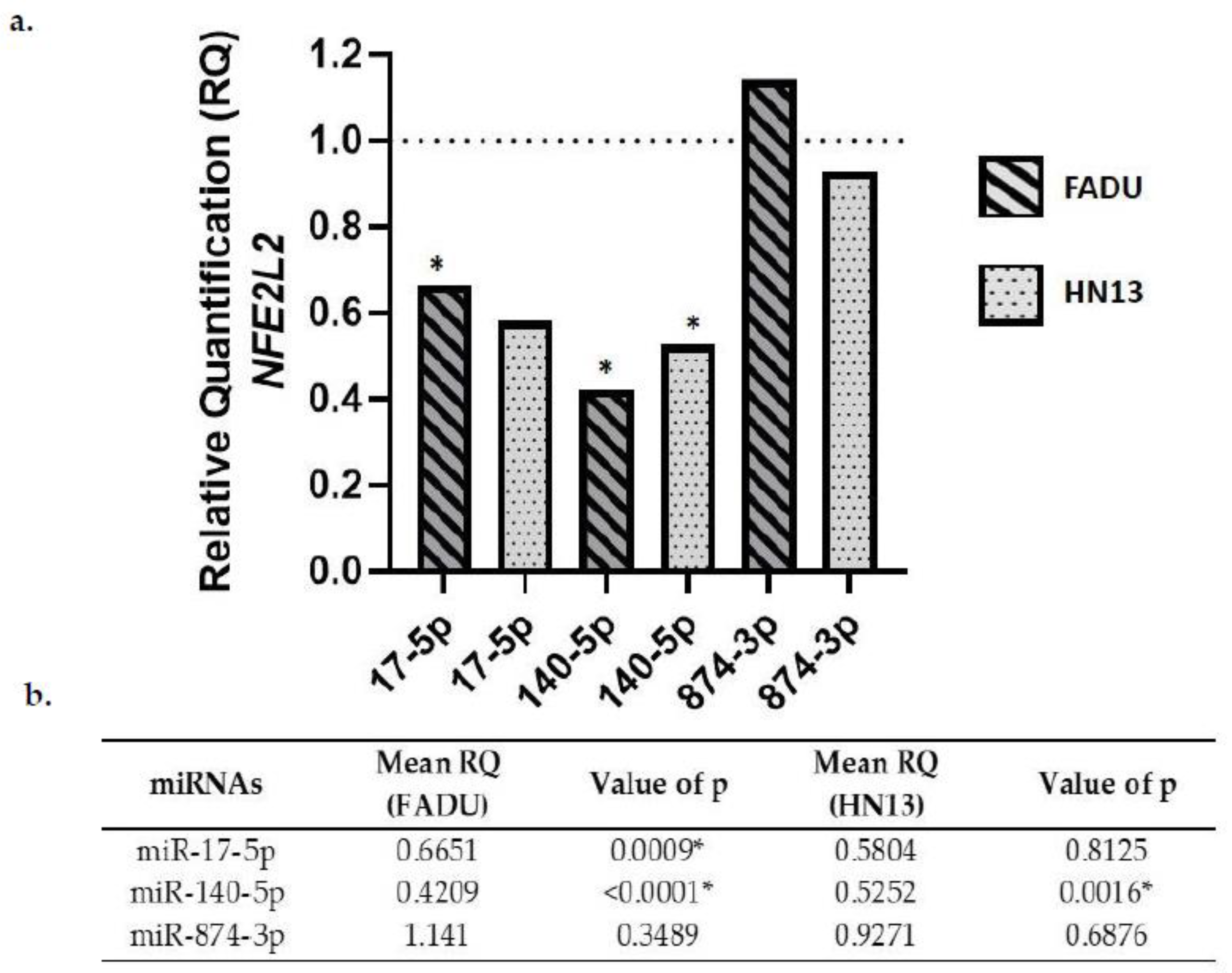
2.5. NFE2L2 Gene Expression in Transfected Cells
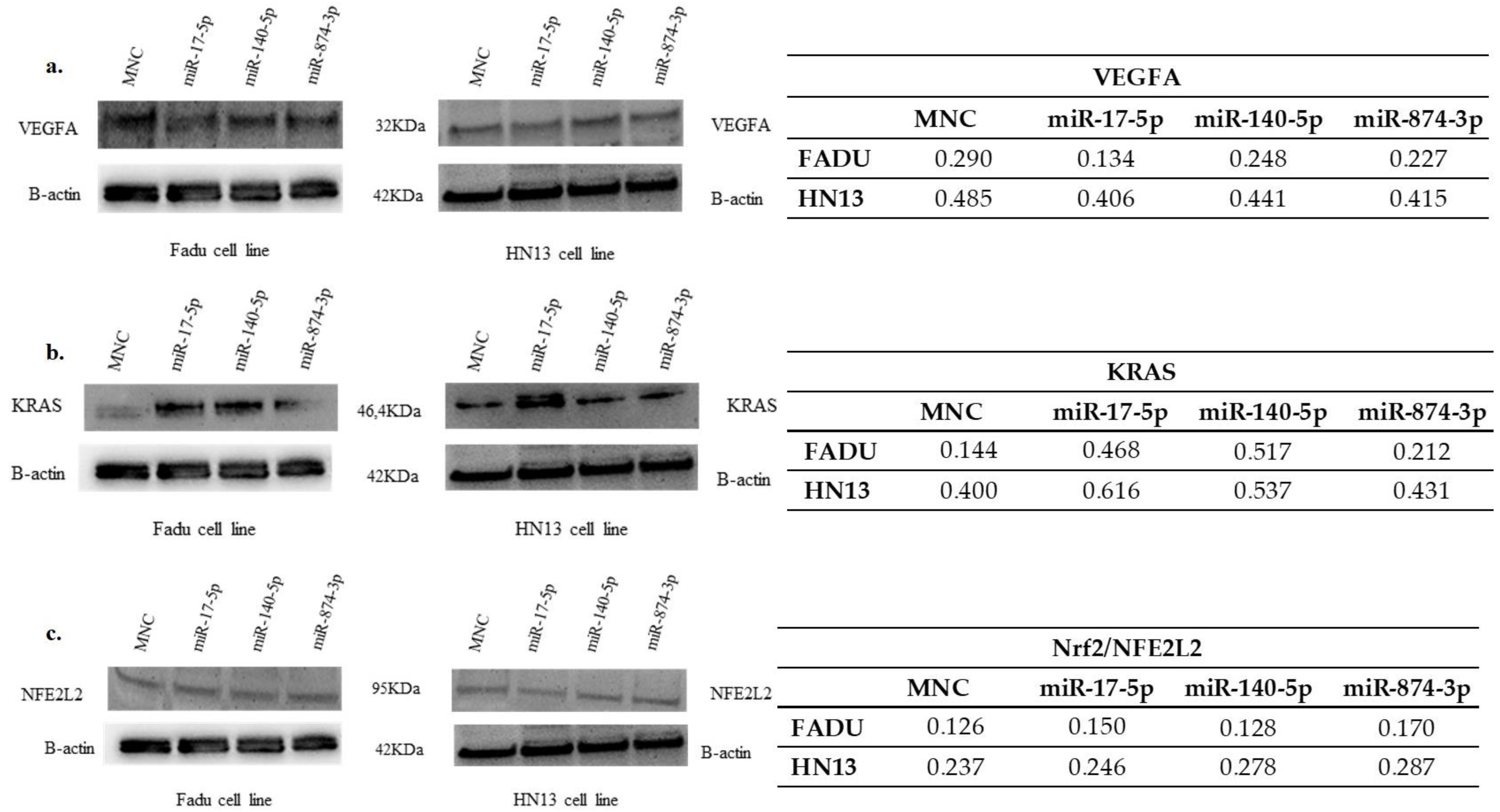
2.6. Expression of VEGFA Protein in Transfected Cells
2.7. Expression of KRAS Protein in Transfected Cells
2.8. Expression of NFE2L2/NRF2 Protein in Transfected Cells
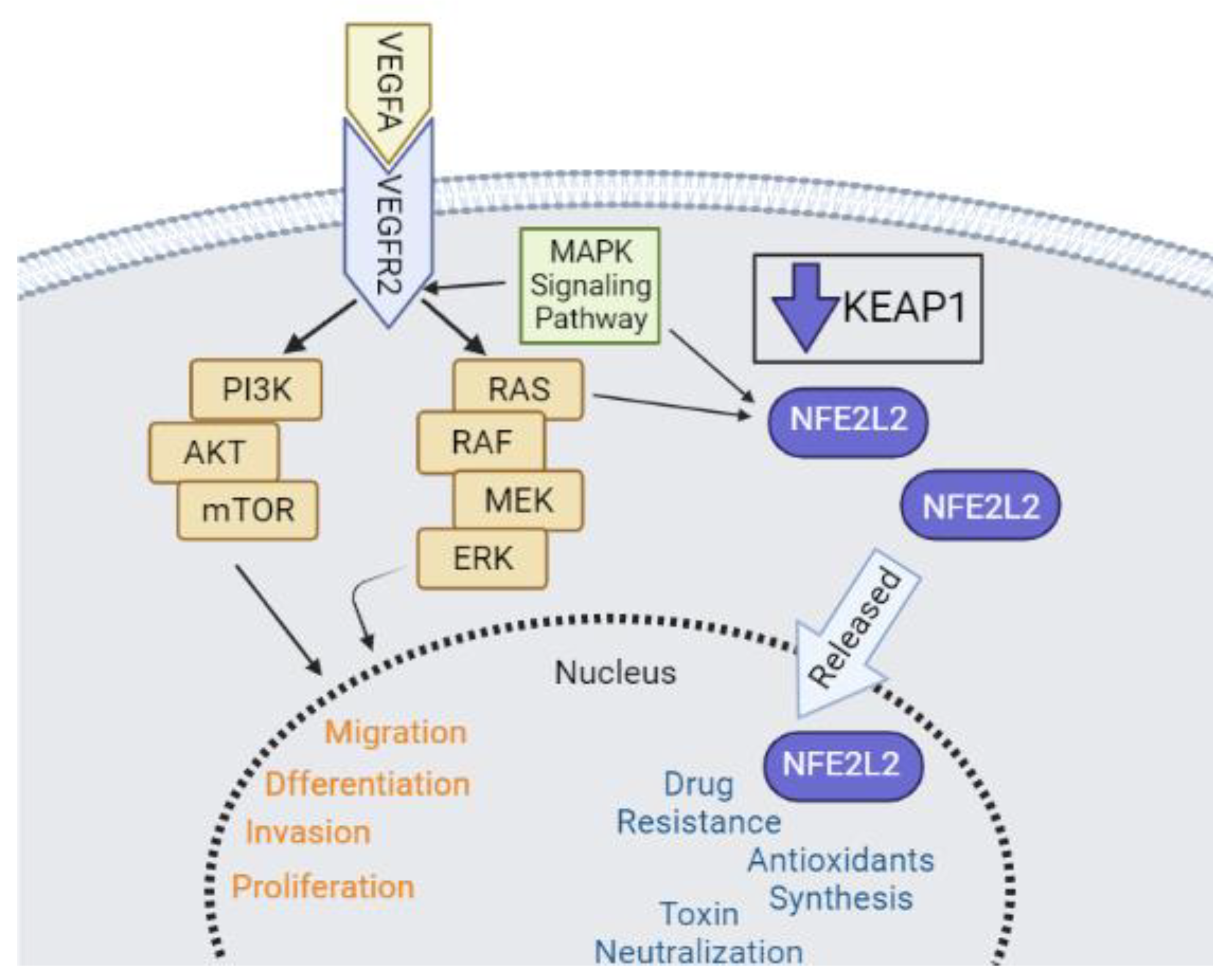
3. Discussion
4. Materials and Methods
4.1. Cell Lines Culture
4.2. Transfection of miRNAs in Cell Lines
4.3. Gene and miRNA Expression
4.4. Protein Quantification and Expression
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaidar-Person, O.; Gil, Z.; Billan, S. Precision medicine in head and neck cancer. Drug Resist. Update 2018, 40, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Svider, P.F.; Blasco, M.A.; Raza, S.N.; Shkoukani, M.; Sukari, A.; Yoo, G.H.; Folbe, A.J.; Lin, H.-S.; Fribley, A.M. Head and Neck Cancer: Underfunded and Understudied? Otolaryngol.–Head Neck Surg. 2016, 156, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, W.M.; Dagan, R.; Bryant, C.M.; Fernandes, R.P. Radiation Oncology for Head and Neck Cancer: Current Standards and Future Changes. Oral. Maxillofac. Surg. Clin. N. Am. 2019, 31, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.; Gonzalez, F.J. The Evolution of Carcinogenesis. Toxicol. Sci. 2018, 165, 272–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Han, J.-W. Targeting epigenetics for cancer therapy. Arch. Pharm. Res. 2019, 42, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Uccelli, A.; Wolff, T.; Valente, P.; Di Maggio, N.; Pellegrino, M.; Gürke, L.; Banfi, A.; Gianni-Barrera, R. Vascular endothelial growth factor biology for regenerative angiogenesis. Swiss Med. Wkly. 2019, 149, w20011. [Google Scholar] [CrossRef]
- Ramjiawan, R.R.; Griffioen, A.W.; Duda, D.G. Anti-angiogenesis for cancer revisited: Is there a role for combinations with immunotherapy? Angiogenesis 2017, 20, 185–204. [Google Scholar] [CrossRef]
- Uprety, D.; Adjei, A.A. KRAS: From undruggable to a druggable Cancer Target. Cancer Treat. Rev. 2020, 89, 102070. [Google Scholar] [CrossRef]
- Ferrer, I.; Zugazagoitia, J.; Herbertz, S.; John, W.; Paz-Ares, L.; Schmid-Bindert, G. KRAS-Mutant non-small cell lung cancer: From biology to therapy. Lung Cancer 2018, 124, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Mo, S.P.; Coulson, J.M.; Prior, I.A. RAS variant signalling. Biochem. Soc. Trans. 2018, 46, 1325–1332. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, S.; Zheng, M.; Dai, L.; Wang, K.; Gao, X.; Cao, M.; Yu, X.; Pang, X.; Zhang, M.; et al. Hypoxia promotes vasculogenic mimicry formation by vascular endothelial growth factor A mediating epithelial-mesenchymal transition in salivary adenoid cystic carcinoma. Cell Prolif. 2019, 52, e12600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, Y.; Takano, O.; Kato, I.; Takahashi, Y.; Shima, F.; Kataoka, T. Ras inhibitors display an anti-metastatic effect by downregulation of lysyl oxidase through inhibition of the Ras-PI3K-Akt-HIF-1α pathway. Cancer Lett. 2017, 410, 82–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menegon, S.; Columbano, A.; Giordano, S. The Dual Roles of NRF2 in Cancer. Trends Mol. Med. 2016, 22, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H.; Motohashi, H. NRF2 addiction in cancer cells. Cancer Sci. 2018, 109, 900–911. [Google Scholar] [CrossRef] [Green Version]
- Hammad, A.; Namani, A.; Elshaer, M.; Wang, X.J.; Tang, X. “NRF2 addiction” in lung cancer cells and its impact on cancer therapy. Cancer Lett. 2019, 467, 40–49. [Google Scholar] [CrossRef]
- Simonson, B.; Das, S. MicroRNA Therapeutics: The Next Magic Bullet? Mini Rev. Med. Chem. 2015, 15, 467–474. [Google Scholar] [CrossRef]
- Ambs, S.; Prueitt, R.L.; Yi, M.; Hudson, R.S.; Howe, T.M.; Petrocca, F.; Wallace, T.A.; Liu, C.-G.; Volinia, S.; Calin, G.A.; et al. Genomic Profiling of MicroRNA and Messenger RNA Reveals Deregulated MicroRNA Expression in Prostate Cancer. Cancer Res. 2008, 68, 6162–6170. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.-U.; Prieto-Vila, M.; Hironaka, A.; Ochiya, T. The role of extracellular vesicle microRNAs in cancer biology. Clin. Chem. Lab. Med. 2017, 55, 648–656. [Google Scholar] [CrossRef]
- Yang, H.; Fang, F.; Chang, R.; Yang, L. MicroRNA-140-5p suppresses tumor growth and metastasis by targeting transforming growth factor β receptor 1 and fibroblast growth factor 9 in hepatocellular carcinoma. Hepatology 2013, 58, 205–217. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Pathol. 2014, 9, 287–314. [Google Scholar] [CrossRef] [Green Version]
- Duan, F.; Yang, Y.; Liu, W.; Zhao, J.; Song, X.; Li, L.; Li, F. Quantifying the prognostic significance of microRNA-17/17-5P in cancers: A meta-analysis based on published studies. Cancer Manag. Res. 2018, 10, 2055–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouza, M.; Bohosova, J.; Stanikova, A.; Pecl, J.; Slaby, O.; Jabandziev, P. MicroRNA as an Early Biomarker of Neonatal Sepsis. Front. Pediatr. 2022, 10, 854324. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xu, Q.; Zhou, M.; Chen, Y.; Jiang, C.; Jiang, Y.; Lin, Y.; He, Q.; Zhao, L.; Dong, Y.; et al. Plasma miR-153 and miR-223 Levels as Potential Biomarkers in Parkinson’s Disease. Front. Neurosci. 2022, 16, 865139. [Google Scholar] [CrossRef] [PubMed]
- Grieco, G.E.; Besharat, Z.M.; Licata, G.; Fignani, D.; Brusco, N.; Nigi, L.; Formichi, C.; Po, A.; Sabato, C.; Dardano, A.; et al. Circulating microRNAs as clinically useful biomarkers for Type 2 Diabetes Mellitus: miRNomics from bench to bedside. Transl. Res. 2022, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Deng, X.; Tian, F.; Li, Z.; Jiang, P.; Zhao, X.; Chen, G.; Chen, Y.; Zheng, P.; Li, D.; et al. The interaction of LOXL2 with GATA6 induces VEGFA expression and angiogenesis in cholangiocarcinoma. Int. J. Oncol. 2019, 55, 657–670. [Google Scholar] [CrossRef] [Green Version]
- Pang, L.; Wang, J.; Fan, Y.; Xu, R.; Bai, Y.; Bai, L. Correlations of TNM staging and lymph node metastasis of gastric cancer with MRI features and VEGF expression. Cancer Biomark. 2018, 23, 53–59. [Google Scholar] [CrossRef]
- Dang, Y.; Luo, D.; Rong, M.; Chen, G. Underexpression of miR-34a in Hepatocellular Carcinoma and Its Contribution towards Enhancement of Proliferating Inhibitory Effects of Agents Targeting c-MET. PLoS ONE 2013, 8, e61054. [Google Scholar] [CrossRef]
- Wang, J.-X.; Jia, X.-J.; Liu, Y.; Dong, J.-H.; Ren, X.-M.; Xu, O.; Liu, S.-H.; Shan, C.-G. Silencing of miR-17-5p suppresses cell proliferation and promotes cell apoptosis by directly targeting PIK3R1 in laryngeal squamous cell carcinoma. Cancer Cell Int. 2020, 20, 14. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Li, X.; Shen, W.; Hu, H.; Li, C.; Han, G. MicroRNA-140 Represses Esophageal Cancer Progression via Targeting ZEB2 to Regulate Wnt/β-Catenin Pathway. J. Surg. Res. 2021, 257, 267–277. [Google Scholar] [CrossRef]
- Zhang, J.R.; Zhu, R.H.; Han, X.P. MiR-140-5p inhibits larynx carcinoma invasion and angiogenesis by targeting VEGF-A. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5994–6001. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Wang, C.; Yang, Z.; Liu, W.; Yuan, Y.; Li, K.; Zhang, Y.; Wang, Y.; Shi, Y.; Qiu, Y.; et al. Dysregulated Sp1/miR-130b-3p/HOXA5 axis contributes to tumor angiogenesis and progression of hepatocellular carcinoma. Theranostics 2020, 10, 5209–5224. [Google Scholar] [CrossRef] [PubMed]
- Castanhole-Nunes, M.M.U.; Tunissiolli, N.M.; Oliveira, A.R.C.P.; Mattos, M.F.; Galbiatti-Dias, A.L.S.; Kawasaki-Oyama, R.S.; Pavarino, E.C.; da Silva, R.F.; Goloni-Bertollo, E.M. MiR-612, miR-637, and miR-874 can Regulate VEGFA Expression in Hepatocellular Carcinoma Cell Lines. Genes 2022, 13, 282. [Google Scholar] [CrossRef]
- Yuan, R.-B.; Zhang, S.-H.; He, Y.; Zhang, X.-Y.; Zhang, Y.-B. MiR-874-3p is an independent prognostic factor and functions as an anti-oncomir in esophageal squamous cell carcinoma via targeting STAT3. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7265–7273. [Google Scholar] [PubMed]
- Cuzziol, C.I.; Castanhole-Nunes, M.M.U.; Pavarino, C.; Goloni-Bertollo, E.M. MicroRNAs as regulators of VEGFA and NFE2L2 in cancer. Gene 2020, 759, 144994. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Liu, B.; Ji, R.; Jiang, X.; Yan, X.; Xin, Y. Targeting the BDNF/TrkB pathway for the treatment of tumors. Oncol. Lett. 2019, 17, 2031–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, P.D.; Guimarães, C.F.; Cardoso, A.P.; Mendonça, S.; Costa, A.M.; Oliveira, M.J.; Velho, S. KRAS Oncogenic Signaling Extends beyond Cancer Cells to Orchestrate the Microenvironment. Cancer Res. 2018, 78, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.P.; Liu, G.L.; Li, S.; Liu, X.L. miR-17-5p knockdown inhibits proliferation, autophagy and promotes apoptosis in thyroid cancer via targeting PTEN. Neoplasma 2020, 67, 249–258. [Google Scholar] [CrossRef]
- Tian, Y.; Li, X.; Bai, C.; Yang, Z.; Zhang, L.; Luo, J. MiR-17-5p promotes the endothelialization of endothelial progenitor cells to facilitate the vascular repair of aneurysm by regulating PTEN-mediated PI3K/AKT/VEGFA pathway. Cell Cycle 2020, 19, 3608–3621. [Google Scholar] [CrossRef]
- De La Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Kim, T.-H.; Hur, E.-G.; Kang, S.-J.; Kim, J.-A.; Thapa, D.; Lee, Y.M.; Ku, S.K.; Jung, Y.; Kwak, M.-K. NRF2 Blockade Suppresses Colon Tumor Angiogenesis by Inhibiting Hypoxia-Induced Activation of HIF-1α. Cancer Res. 2011, 71, 2260–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Qiu, J.; Kang, J.; Xing, X.; Shi, X.; Wei, Y. Exposure to tris(1,3-dichloro-2-propyl) phosphate (TDCPP) induces vascular toxicity through Nrf2-VEGF pathway in zebrafish and human umbilical vein endothelial cells. Environ. Pollut. 2019, 247, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Goswami, D.; Adiseshaiah, P.P.; Burgan, W.; Yi, M.; Guerin, T.M.; Kozlov, S.V.; Nissley, D.V.; McCormick, F. Undermining Glutaminolysis Bolsters Chemotherapy While NRF2 Promotes Chemoresistance in KRAS-Driven Pancreatic Cancers. Cancer Res. 2020, 80, 1630–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeNicola, G.M.; Karreth, F.A.; Humpton, T.J.; Gopinathan, A.; Wei, C.; Frese, K.; Mangal, D.; Yu, K.H.; Yeo, C.J.; Calhoun, E.S.; et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 2011, 475, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Hu, L.; Lu, K.; Wang, Y.; Xie, Y.; Gao, L.; Yang, G.; Xie, B.; He, W.; Chen, G.; et al. Ferroportin downregulation promotes cell proliferation by modulating the Nrf2–miR-17-5p axis in multiple myeloma. Cell Death Dis. 2019, 10, 624. [Google Scholar] [CrossRef] [PubMed]
- Stuchi, L.P.; Castanhole-Nunes, M.M.U.; Maniezzo-Stuchi, N.; Biselli-Chicote, P.M.; Henrique, T.; Neto, J.A.P.; Neto, D.D.-S.; Girol, A.P.; Pavarino, E.C.; Goloni-Bertollo, E.M. VEGFA and NFE2L2 Gene Expression and Regulation by MicroRNAs in Thyroid Papillary Cancer and Colloid Goiter. Genes 2020, 11, 954. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ren, K.; Liu, S.; Li, W.; Huang, C.; Yang, X. MicroRNA-140-5p aggravates hypertension and oxidative stress of atherosclerosis via targeting Nrf2 and Sirt2. Int. J. Mol. Med. 2018, 43, 839–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Liu, H.; Xu, J.; Shi, D.; Zhai, L.; Liu, B.; Wang, L.; Liu, G.; Qin, J. miR-144-3p regulates the resistance of lung cancer to cisplatin by targeting Nrf2. Oncol. Rep. 2018, 40, 3479–3488. [Google Scholar] [CrossRef] [Green Version]
- Hausser, J.; Mayo, A.; Keren, L.; Alon, U. Central dogma rates and the trade-off between precision and economy in gene expression. Nat. Commun. 2019, 10, 68. [Google Scholar] [CrossRef] [Green Version]
- Buccitelli, C.; Selbach, M. mRNAs, proteins and the emerging principles of gene expression control. Nat. Rev. Genet. 2020, 21, 630–644. [Google Scholar] [CrossRef]
- Tokar, T.; Pastrello, C.; Rossos, A.E.M.; Abovsky, M.; Hauschild, A.-C.; Tsay, M.; Lu, R.; Jurisica, I. mirDIP 4.1—Integrative database of human microRNA target predictions. Nucleic Acids Res. 2017, 46, D360–D370. [Google Scholar] [CrossRef] [PubMed]
- Shirdel, E.A.; Xie, W.; Mak, T.W.; Jurisica, I. NAViGaTing the Micronome—Using Multiple MicroRNA Prediction Databases to Identify Signalling Pathway-Associated MicroRNAs. PLoS ONE 2011, 6, e17429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs | Mean RQ (FADU) | Value of p | Mean RQ (HN13) | Value of p |
|---|---|---|---|---|
| miR-17-5p | 407.9 | 0.0087 * | 182.2 | 0.3316 |
| miR-140-5p | 15.620 | 0.0121 * | 3.322 | 0.0024 * |
| miR-874-3p | 2.871 | 0.0313 * | 13.265 | 0.0642 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuzziol, C.I.; Marzochi, L.L.; Possebon, V.S.; Kawasaki-Oyama, R.S.; Mattos, M.F.; Junior, V.S.; Ferreira, L.A.M.; Pavarino, É.C.; Castanhole-Nunes, M.M.U.; Goloni-Bertollo, E.M. Regulation of VEGFA, KRAS, and NFE2L2 Oncogenes by MicroRNAs in Head and Neck Cancer. Int. J. Mol. Sci. 2022, 23, 7483. https://doi.org/10.3390/ijms23137483
Cuzziol CI, Marzochi LL, Possebon VS, Kawasaki-Oyama RS, Mattos MF, Junior VS, Ferreira LAM, Pavarino ÉC, Castanhole-Nunes MMU, Goloni-Bertollo EM. Regulation of VEGFA, KRAS, and NFE2L2 Oncogenes by MicroRNAs in Head and Neck Cancer. International Journal of Molecular Sciences. 2022; 23(13):7483. https://doi.org/10.3390/ijms23137483
Chicago/Turabian StyleCuzziol, Caroline Izak, Ludimila Leite Marzochi, Vitória Scavacini Possebon, Rosa Sayoko Kawasaki-Oyama, Marlon Fraga Mattos, Vilson Serafim Junior, Letícia Antunes Muniz Ferreira, Érika Cristina Pavarino, Márcia Maria Urbanin Castanhole-Nunes, and Eny Maria Goloni-Bertollo. 2022. "Regulation of VEGFA, KRAS, and NFE2L2 Oncogenes by MicroRNAs in Head and Neck Cancer" International Journal of Molecular Sciences 23, no. 13: 7483. https://doi.org/10.3390/ijms23137483