
Collagen Mimetic Peptides Promote Adherence and Migration of ARPE-19 Cells While Reducing Inflammatory and Oxidative Stress

 , and
, and 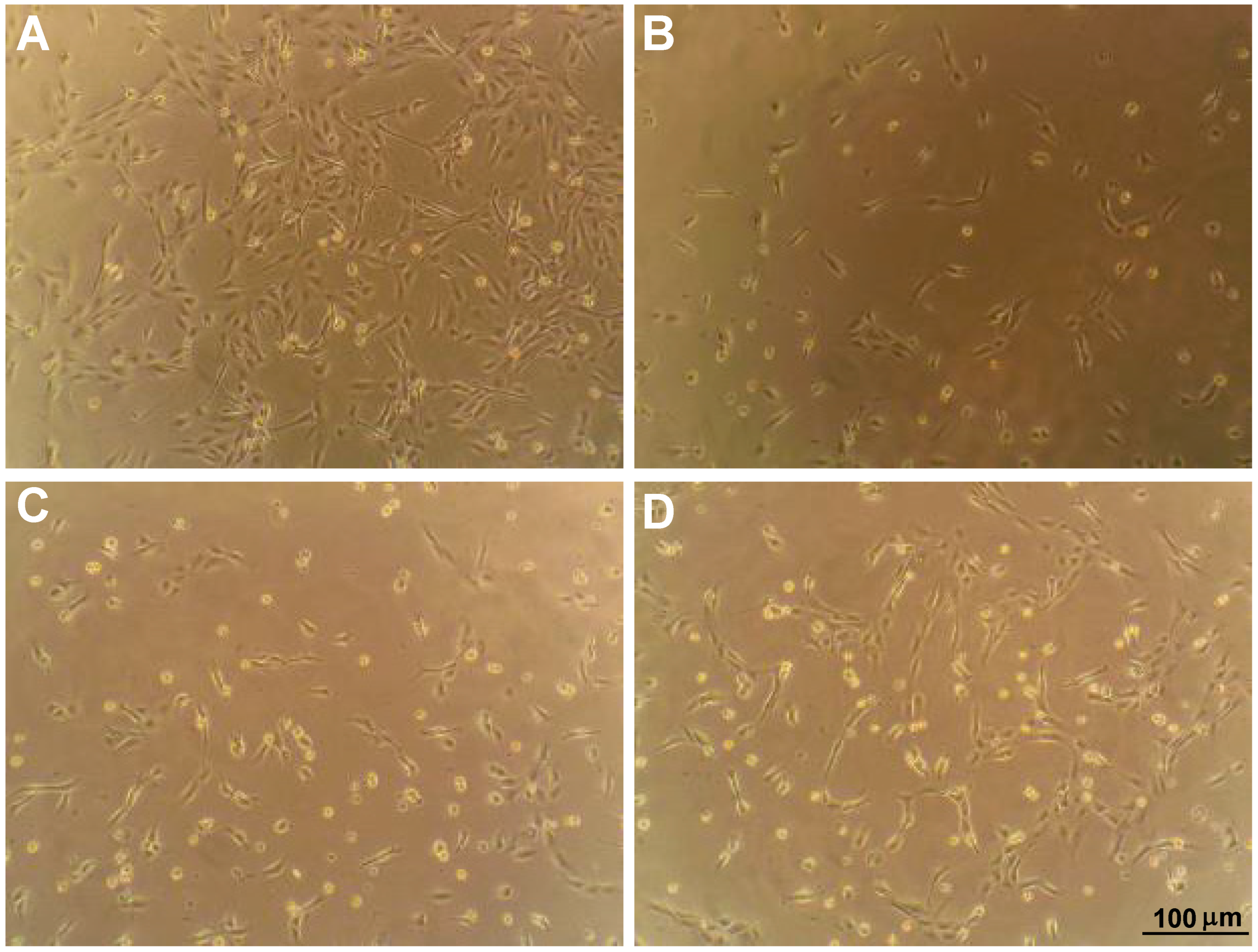{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
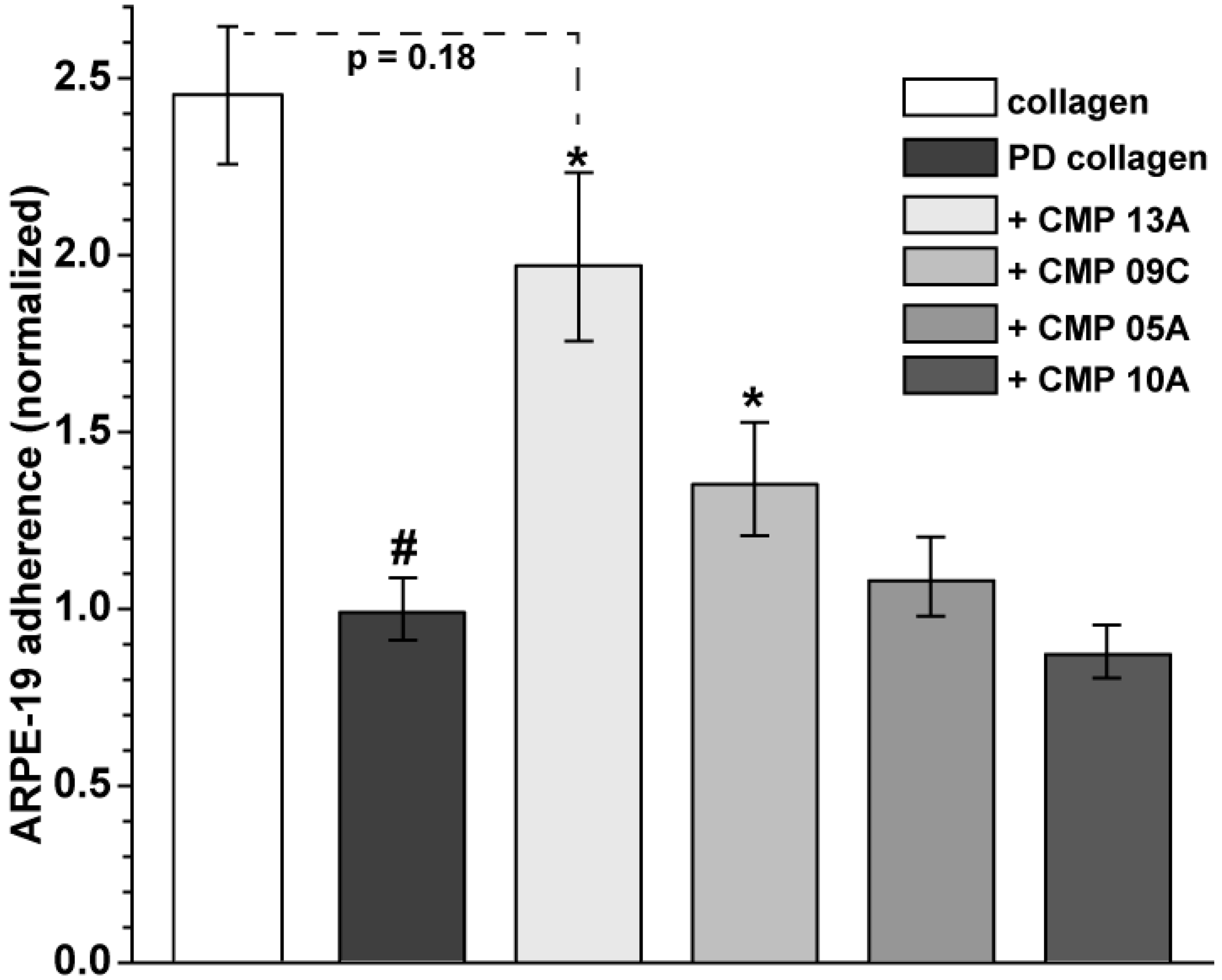
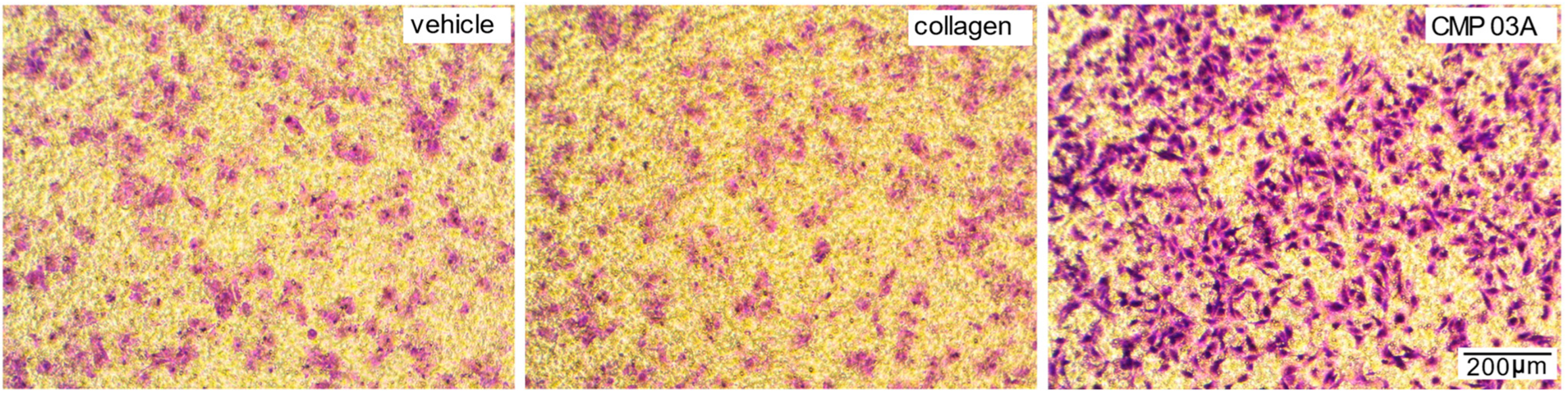
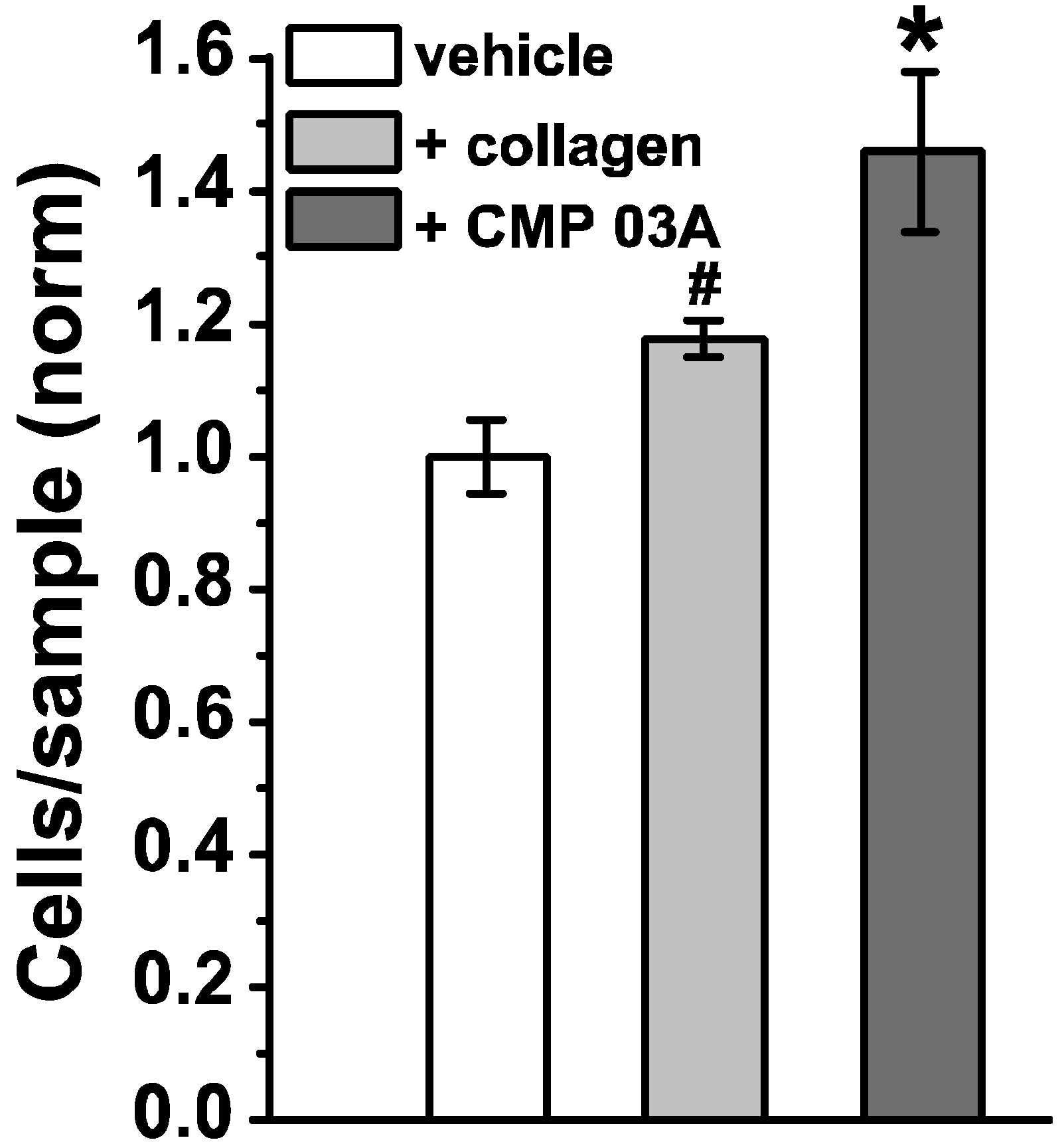
2.1. Collagen Mimetic Peptides Promote ARPE-19 Cell Adherence and Migration
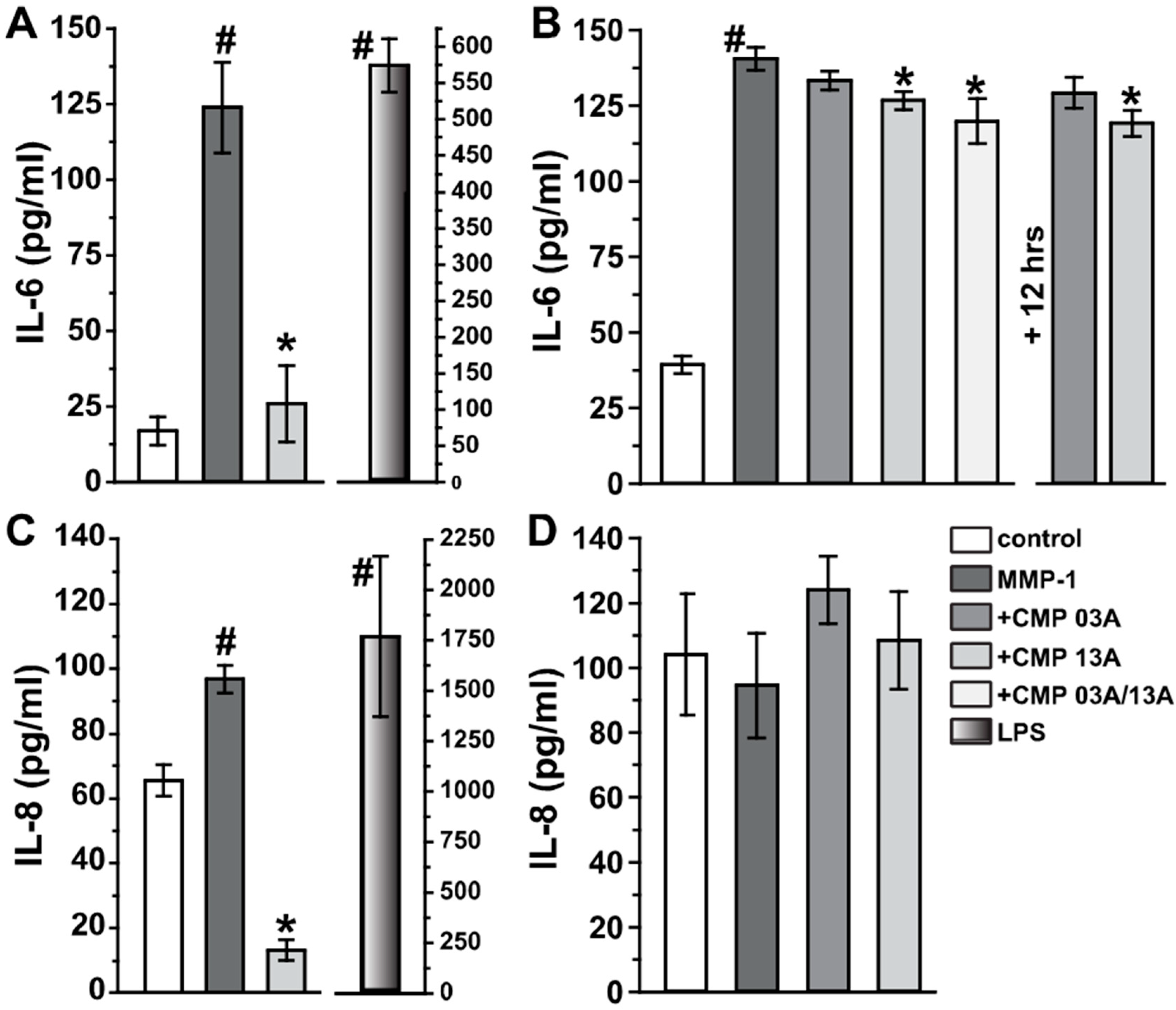
2.2. Collagen Mimetic Peptides Modulate ARPE-19 Cell Inflammatory and Oxidative Stress Mediators
3. Discussion
4. Materials and Methods
4.1. ARPE-19 Cell Cultures
4.2. Adherence and Migration Assays
4.3. ELISA
4.4. Reactive Oxygen Species and Caspase-3 Measurements
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baratta, R.O.; Schlumpf, E.; Del Buono, B.J.; DeLorey, S.S.; Calkins, D.J. Corneal collagen as a potential therapeutic target in dry eye disease. Surv. Ophthalmol. 2022, 67, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.V.; Gupta, S.; Bansal, H.; Singla, K. Collagen in health and disease. J. Orofac. Res. 2012, 2, 153–159. [Google Scholar] [CrossRef]
- Morrison, J.C.; Jerdan, J.A.; L’Hernault, N.L.; Quigley, H.A. The extracellular matrix composition of the monkey optic nerve head. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1141–1150. [Google Scholar]
- Huang, W.; Fan, Q.; Wang, W.; Zhou, M.; Laties, A.M.; Zhang, X. Collagen: A potential factor involved in the pathogenesis of glaucoma. Med. Sci. Monit. Basic Res. 2013, 19, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romaniuk, W.; Nita, M.; Strzalka-Mrozik, B.; Grzybowski, A.; Mazurek, U. Age-related macular degeneration and changes in the extracellular matrix. Med. Sci. Monit. 2014, 20, 1003–1016. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J. The 2014 Bowman lecture—Bowman’s and Bruch’s: A tale of two membranes during the laser revolution. Eye 2015, 29, 46–64. [Google Scholar] [CrossRef] [Green Version]
- Nandrot, E.F.; Kim, Y.; Brodie, S.; Huang, X.; Sheppard, D.; Finnemann, S.C. Loss of synchronized retinal phagocytosis and age-related blindness in mice lacking αvβ5 integrin. J. Exp. Med. 2004, 200, 1539–1545. [Google Scholar] [CrossRef]
- Bandyopadhyay, M.; Rohrer, B. Matrix Metalloproteinase activity creates pro-angiogenic environment in primary human retinal pigment epithelial cells exposed to complement. Investig. Opthalmol. Vis. Sci. 2012, 53, 1953–1961. [Google Scholar] [CrossRef] [Green Version]
- Alcazar, O.; Cousins, S.W.; Striker, G.E.; Marin-Castano, M.E. (Pro)renin receptor is expressed in human retinal pigment epithelium and participates in extracellular matrix remodeling. Exp. Eye Res. 2009, 89, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [Green Version]
- Kigasawa, K.; Ishikawa, H.; Obazawa, H.; Minamoto, T.; Nagai, Y.; Tanaka, Y. Collagen production by cultured human retinal pigment epithelial cells. Tokai J. Exp. Clin. Med. 1998, 23, 147–151. [Google Scholar] [PubMed]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Fallas, J.A.; O’Leary, L.E.R.; Hartgerink, J.D. Synthetic collagen mimics: Self-assembly of homotrimers, heterotrimers and higher order structures. Chem. Soc. Rev. 2010, 39, 3510–3527. [Google Scholar] [CrossRef] [PubMed]
- Hapach, L.; VanderBurgh, J.; Miller, J.P.; Reinhart-King, C. Manipulation ofin vitrocollagen matrix architecture for scaffolds of improved physiological relevance. Phys. Biol. 2015, 12, 61002. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Murphy, C.J.; McAnulty, J.F.; Raines, R.T. Peptides that anneal to natural collagen in vitro and ex vivo. Org. Biomol. Chem. 2012, 10, 5892–5897. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Guthrie, K.M.; Teixeira, L.; Murphy, C.J.; Dubielzig, R.R.; McAnulty, J.F.; Raines, R.T. Anchoring a cytoactive factor in a wound bed promotes healing. J. Tissue Eng. Regen. Med. 2016, 10, 1012–1020. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Raines, R.T. Collagen-based biomaterials for wound healing. Biopolymers 2014, 101, 821–833. [Google Scholar] [CrossRef] [Green Version]
- Baratta, R.O.; Del Buono, B.J.; Schlumpf, E.; Ceresa, B.P.; Calkins, D.J. Collagen Mimetic Peptides Promote Corneal Epithelial Cell Regeneration. Front. Pharmacol. 2021, 12, 705623. [Google Scholar] [CrossRef]
- McGrady, N.R.; Pasini, S.; Baratta, R.O.; Del Buono, B.J.; Schlumpf, E.; Calkins, D.J. Restoring the extracellular matrix: A neuroprotective role for collagen mimetic peptides in experimental glaucoma. Front. Pharmacol 2021, 12, 764709. [Google Scholar] [CrossRef]
- Alizadeh, M.; Wada, M.; Gelfman, C.M.; Handa, J.T.; Hjelmeland, L.M. Downregulation of differentiation specific gene expression by oxidative stress in ARPE-19 cells. Investig. Ophthalmol. Vis. Sci 2001, 42, 2706–2713. [Google Scholar]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a human retinal pigment epithelial cell line with differentiated properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- Samuel, W.; Jaworski, C.; Postnikova, O.A.; Kutty, R.K.; Duncan, T.; Tan, L.X.; Poliakov, E.; Lakkaraju, A.; Redmond, T.M. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol. Vis. 2017, 23, 60–89. [Google Scholar]
- Zhang, C.; Baffi, J.; Cousins, S.W.; Csaky, K.G. Oxidant-induced cell death in retinal pigment epithelium cells mediated through the release of apoptosis-inducing factor. J. Cell Sci. 2003, 116, 1915–1923. [Google Scholar] [CrossRef] [Green Version]
- Afshari, F.T.; Fawcett, J.W. Improving RPE adhesion to Bruch’s membrane. Eye 2009, 23, 1890–1893. [Google Scholar] [CrossRef]
- Gullapalli, V.K.; Sugino, I.K.; Van Patten, Y.; Shah, S.; Zarbin, M.A. Impaired RPE survival on aged submacular human Bruch’s membrane. Exp. Eye Res. 2005, 80, 235–248. [Google Scholar] [CrossRef]
- Sun, K.; Cai, H.; Tezel, T.H.; Paik, D.; Gaillard, E.R.; Del Priore, L.V. Bruch’s membrane aging decreases phagocytosis of outer segments by retinal pigment epithelium. Mol. Vis. 2007, 13, 2310–2319. [Google Scholar]
- Nicolò, M.; Piccolino, F.C.; Zardi, L.; Giovannini, A.; Mariotti, C. Detection of tenascin-C in surgically excised choroidal neovascular membranes. Graefes Arch. Clin. Exp. Ophthalmol. 2000, 238, 107–111. [Google Scholar] [CrossRef]
- Siwik, D.A.; Pagano, P.J.; Colucci, W.S. Oxidative stress regulates collagen synthesis and matrix metalloproteinase activity in cardiac fibroblasts. Am. J. Physiol. Cell. Physiol. 2001, 280, C53–C60. [Google Scholar] [CrossRef]
- Fisher, G.J.; Quan, T.; Purohit, T.; Shao, Y.; Cho, M.K.; He, T.; Varani, J.; Kang, S.; Voorhees, J.J. Collagen fragmentation promotes oxidative stress and elevates matrix metalloproteinase-1 in fibroblasts in aged human skin. Am. J. Pathol. 2009, 174, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Wang, S. Not All Stressors Are Equal: Mechanism of Stressors on RPE Cell Degeneration. Front. Cell Dev. Biol. 2020, 8, 591067. [Google Scholar] [CrossRef]
- Kurz, T.; Karlsson, M.; Brunk, U.T.; Nilsson, S.E.; Frennesson, C. ARPE-19 retinal pigment epithelial cells are highly resistant to oxidative stress and exercise strict control over their lysosomal redox-active iron. Autophagy 2009, 5, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.; McGrady, N.R.; Baratta, R.O.; Del Buono, B.J.; Schlumpf, E.; Calkins, D.J. Intraocular Delivery of a Collagen Mimetic Peptide Repairs Retinal Ganglion Cell Axons in Chronic and Acute Injury Models. Int J. Mol. Sci 2022, 23, 2911. [Google Scholar] [CrossRef]
- Booij, J.C.; Baas, D.C.; Beisekeeva, J.; Gorgels, T.G.; Bergen, A.A. The dynamic nature of Bruch’s membrane. Prog. Retin. Eye Res. 2010, 29, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Murali, A.; Krishnakumar, S.; Subramanian, A.; Parameswaran, S. Bruch’s membrane pathology: A mechanistic perspective. Eur. J. Ophthalmol. 2020, 30, 1195–1206. [Google Scholar] [CrossRef]
- Binder, S.; Stanzel, B.V.; Krebs, I.; Glittenberg, C. Transplantation of the RPE in AMD. Prog. Retin. Eye Res. 2007, 26, 516–554. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, G.M.; Van Rossem, M.; de Vos, A.F.; Willekens, B.; Peek, R.; Kijlstra, A. Polarized secretion of IL-6 and IL-8 by human retinal pigment epithelial cells. Clin. Exp. Immunol. 1998, 112, 34–43. [Google Scholar] [CrossRef]
- Leung, K.W.; Barnstable, C.J.; Tombran-Tink, J. Bacterial endotoxin activates retinal pigment epithelial cells and induces their degeneration through IL-6 and IL-8 autocrine signaling. Mol. Immunol. 2009, 46, 1374–1386. [Google Scholar] [CrossRef]
- Juel, H.B.; Faber, C.; Udsen, M.S.; Folkersen, L.; Nissen, M.H. Chemokine expression in retinal pigment epithelial ARPE-19 cells in response to coculture with activated T cells. Invest. Ophthalmol. Vis. Sci. 2012, 53, 8472–8480. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Sato, Y.; Kudo, A.; Watanabe, T.; Hirakata, A.; Okada, A.A.; Umezawa, K.; Keino, H. Anti-inflammatory effects of the NF-κB inhibitor dehydroxymethylepoxyquinomicin on ARPE-19 cells. Mol. Med. Rep. 2020, 22, 582–590. [Google Scholar] [CrossRef]
- Fuchs, H.R.; Meister, R.; Lotke, R.; Framme, C. The microRNAs miR-302d and miR-93 inhibit TGFB-mediated EMT and VEGFA secretion from ARPE-19 cells. Exp. Eye Res. 2020, 201, 108258. [Google Scholar] [CrossRef]
- Galazka, G.; Windsor, L.J.; Birkedal-Hansen, H.; Engler, J.A. APMA (4-aminophenylmercuric acetate) activation of stromelysin-1 involves protein interactions in addition to those with cysteine-75 in the propeptide. Biochemistry 1996, 35, 11221–11227. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.A.; Jacob, R. Agonist-induced calcium and oxidative stress responses in endothelial cells. Biochem. Soc. Trans. 2003, 31, 960–962. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, R.W.; Johnson, D.K.; Ziegler, L.M.; DiMattina, A.J.; Stone, R.I.; Holland, J.A. Endothelial NADPH oxidase: Mechanism of activation by low-density lipoprotein. Endothelium 2003, 10, 291–297. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, M.; Pasini, S.; Baratta, R.O.; Del Buono, B.J.; Schlumpf, E.; Calkins, D.J. Collagen Mimetic Peptides Promote Adherence and Migration of ARPE-19 Cells While Reducing Inflammatory and Oxidative Stress. Int. J. Mol. Sci. 2022, 23, 7004. https://doi.org/10.3390/ijms23137004
Ribeiro M, Pasini S, Baratta RO, Del Buono BJ, Schlumpf E, Calkins DJ. Collagen Mimetic Peptides Promote Adherence and Migration of ARPE-19 Cells While Reducing Inflammatory and Oxidative Stress. International Journal of Molecular Sciences. 2022; 23(13):7004. https://doi.org/10.3390/ijms23137004
Chicago/Turabian StyleRibeiro, Marcio, Silvia Pasini, Robert O. Baratta, Brian J. Del Buono, Eric Schlumpf, and David J. Calkins. 2022. "Collagen Mimetic Peptides Promote Adherence and Migration of ARPE-19 Cells While Reducing Inflammatory and Oxidative Stress" International Journal of Molecular Sciences 23, no. 13: 7004. https://doi.org/10.3390/ijms23137004