
miR-218-5p/RUNX2 Axis Positively Regulates Proliferation and Is Associated with Poor Prognosis in Cervical Cancer

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
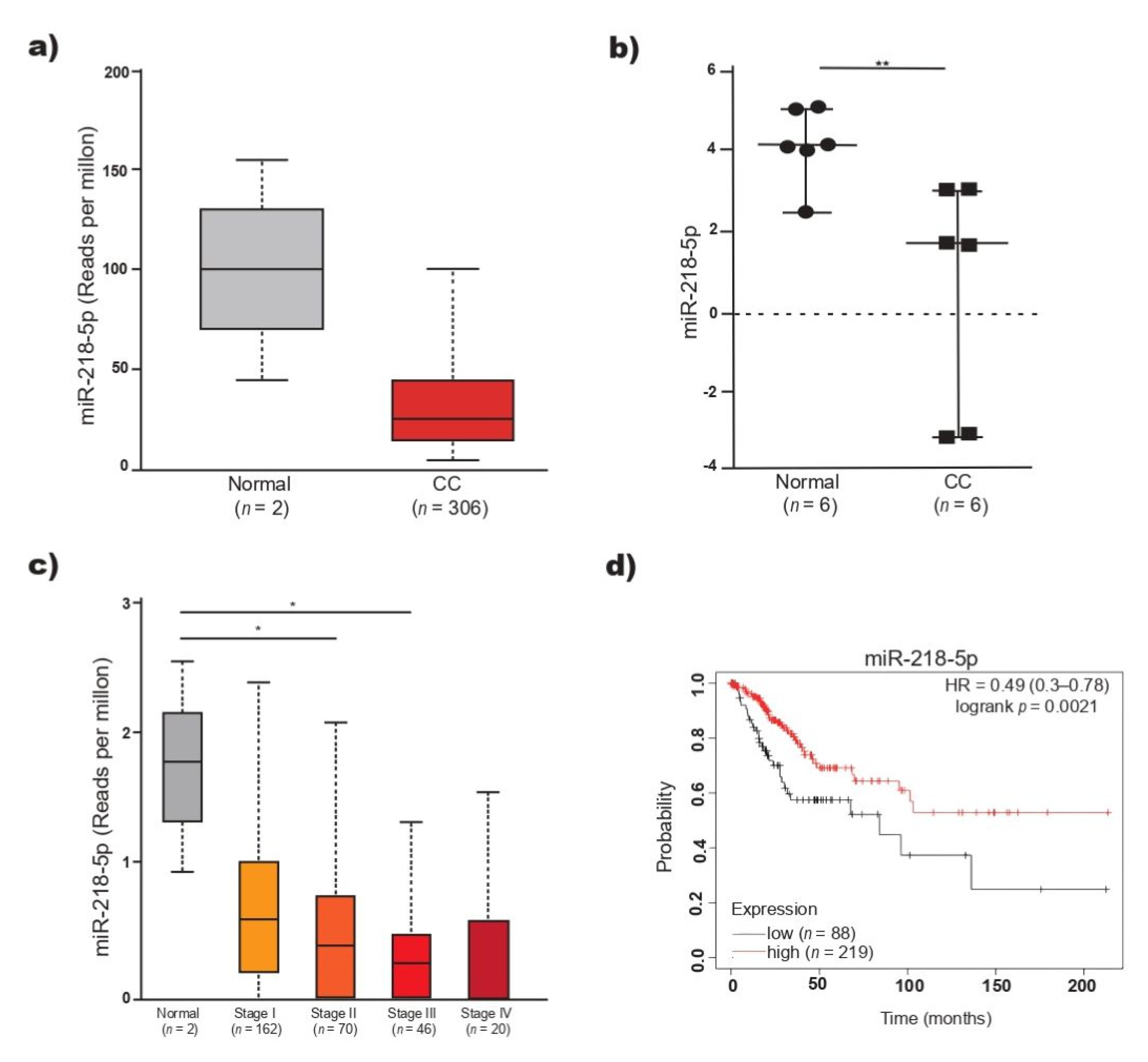
2.1. Low miR-218-5p Expression Is Associated with Poor Prognosis in Patients with CC
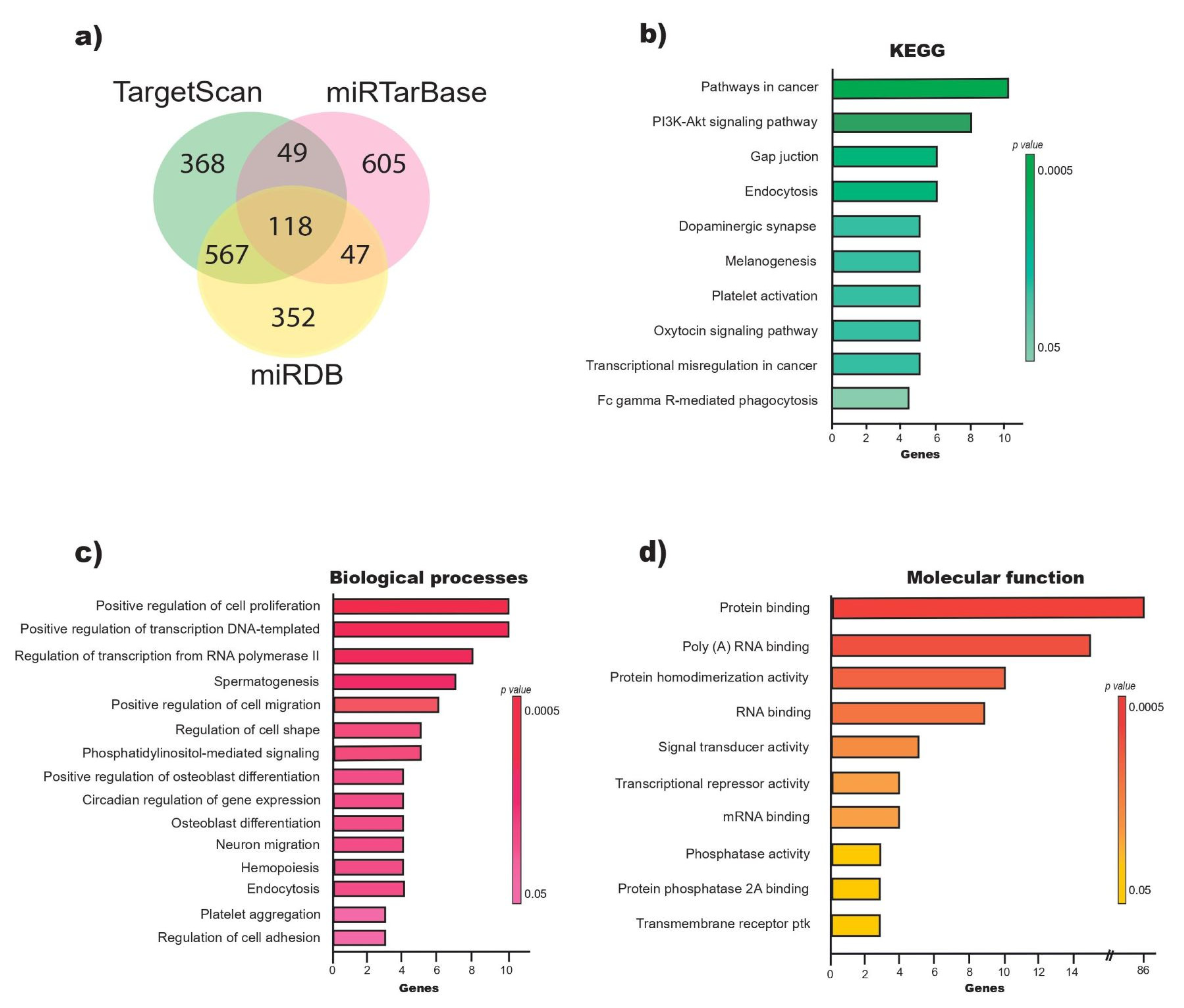
2.2. miR-218-5p Regulates Genes Involved in the Positive Regulation of Cell Proliferation
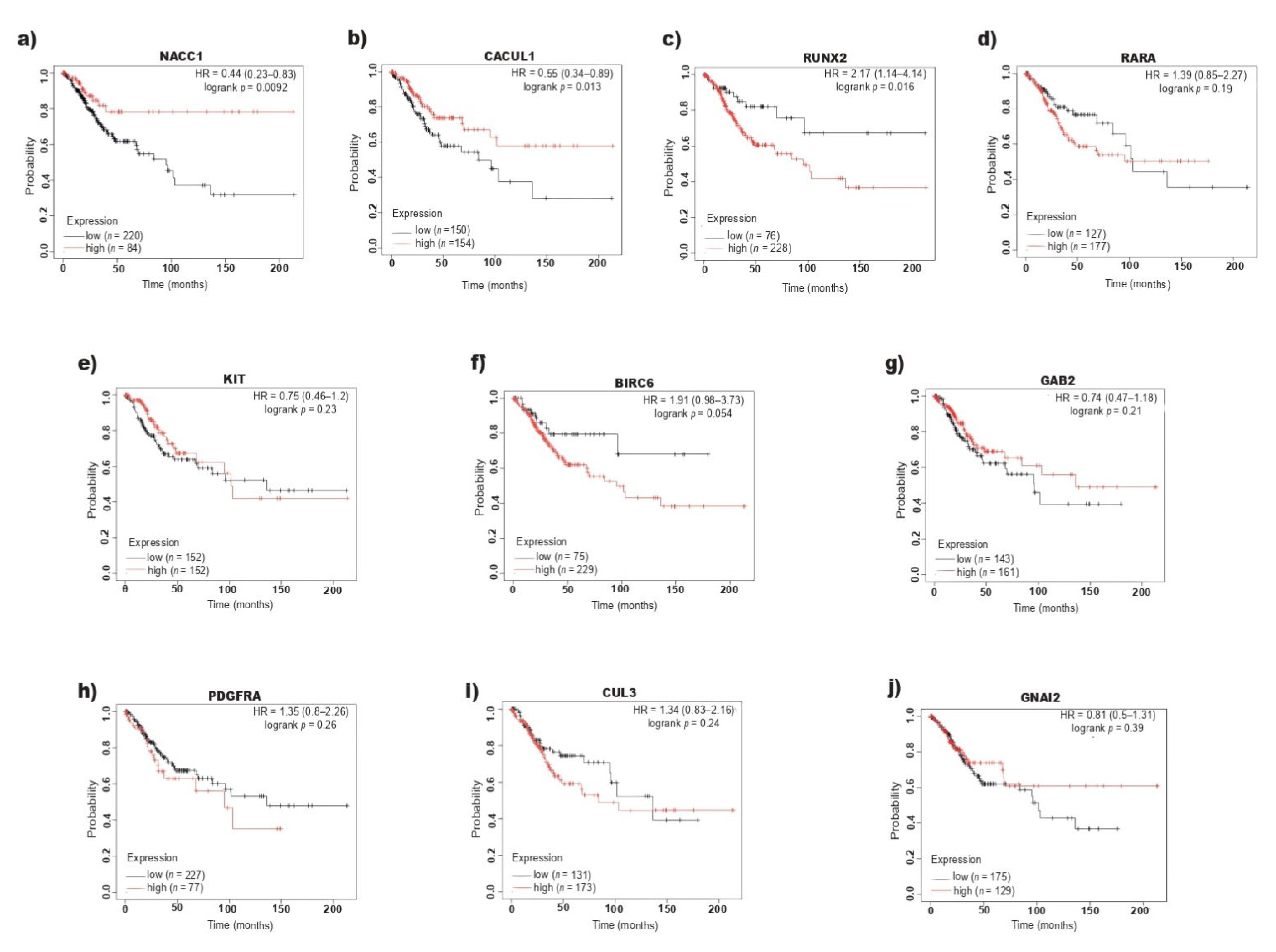
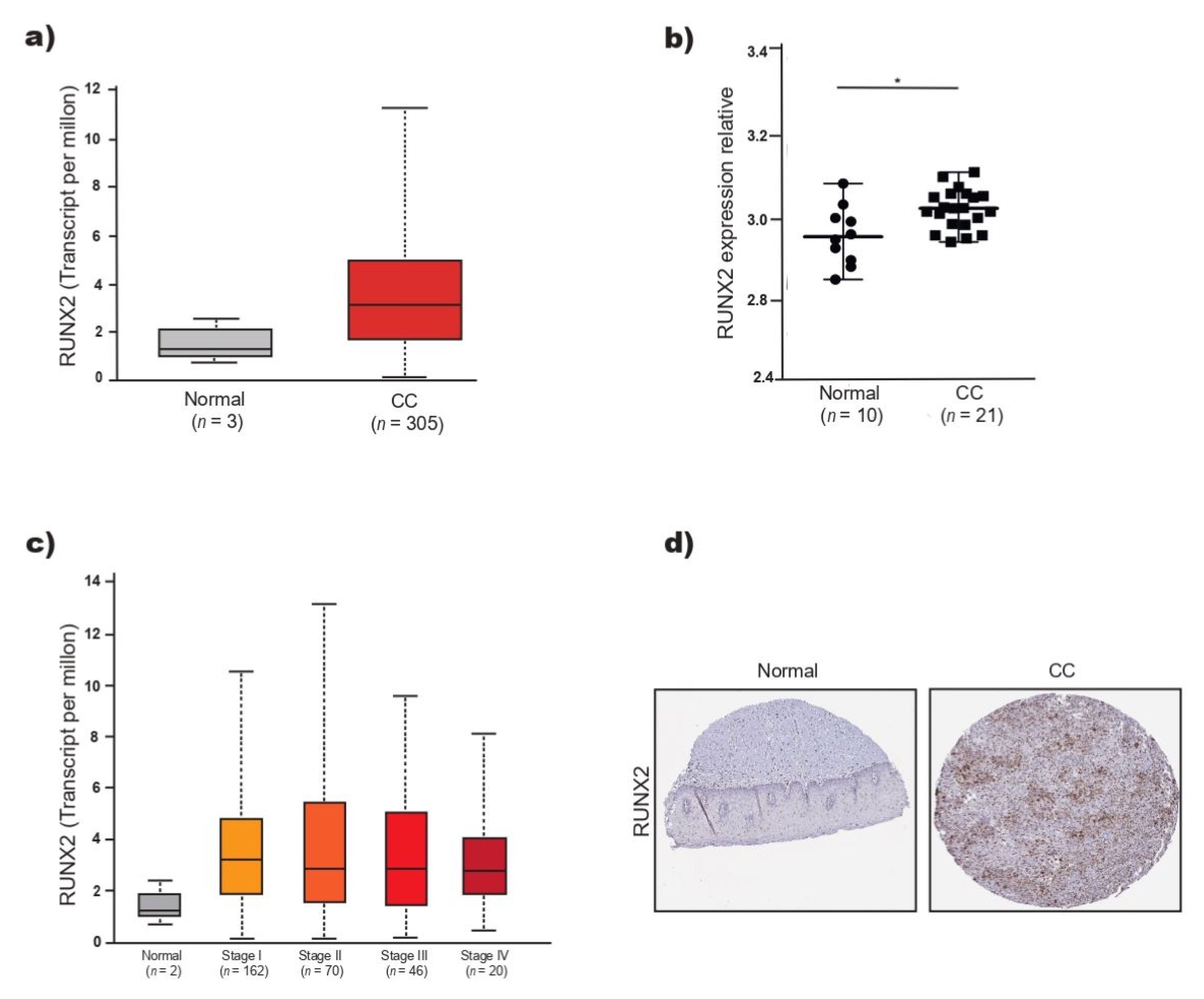
2.3. RUNX2 Overexpression Correlates with Poor Overall Survival in CC
2.4. RUNX2 Participates in the Positive Regulation of Cell Proliferation in CC
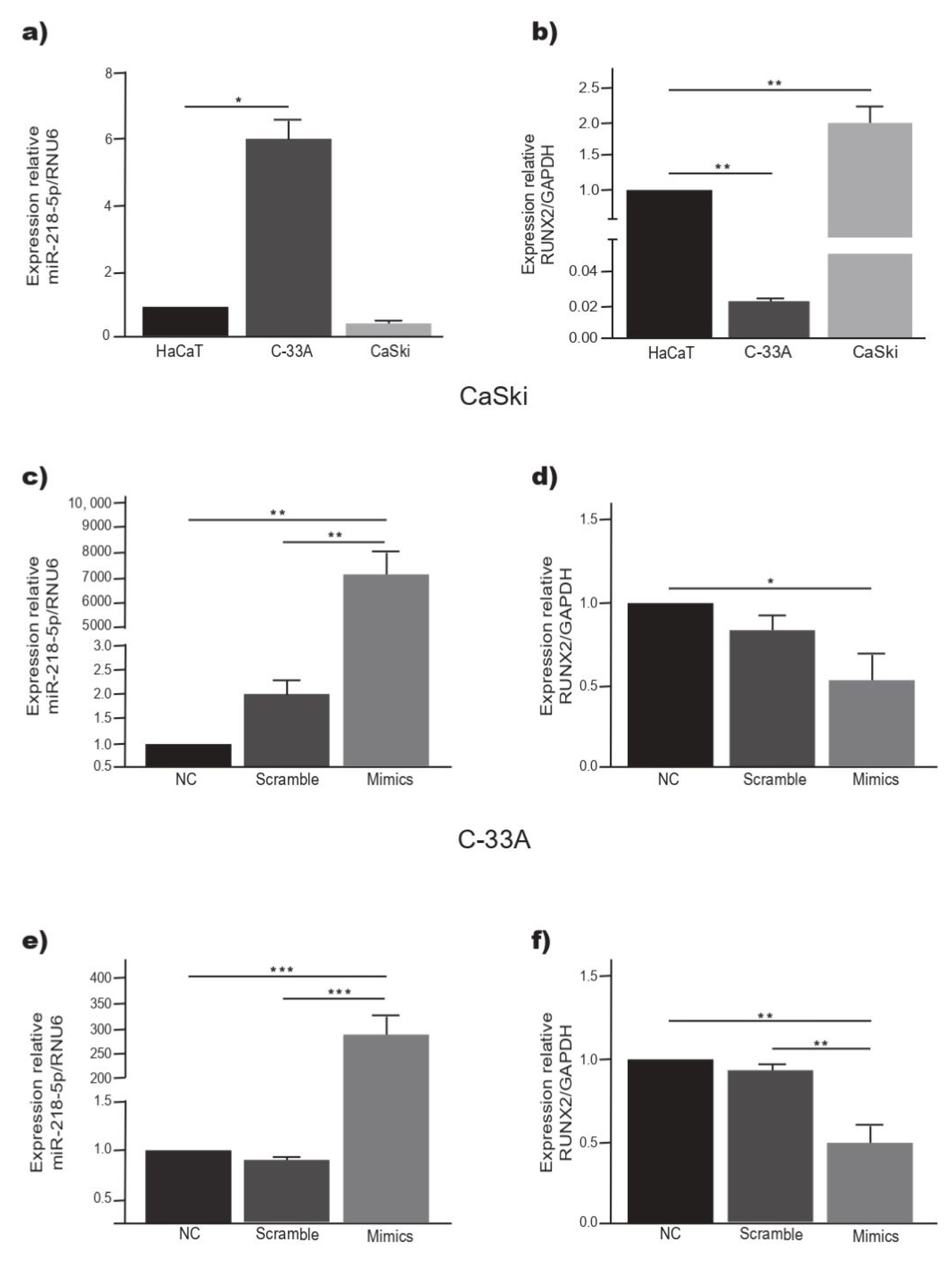
2.5. Overexpression of miR-218-5p Decreases the Expression of RUNX2 in C-33A and CaSki Cells
3. Discussion
4. Materials and Methods
4.1. Prediction of miR-218-5p Target Genes
4.2. GO Enrichment and KEGG Pathway Analysis of the miR-218-5p Target Genes
4.3. Expression Analysis in CC Samples from the TCGA, GEO and HPA Databases
4.4. Survival Analysis
4.5. Cell Culture
4.6. Cell Transfection
4.7. RNA Extraction and qRT–PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Benard, V.B.; Watson, M.; Castle, P.E.; Saraiya, M. Cervical carcinoma rates among young females in the United States. Obstet. Gynecol. 2012, 120, 1117. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, H.; Zhang, L.; Qiao, Y. Cervical cancer: Epidemiology, risk factors and screening. Chin. J. Cancer Res. 2020, 32, 720. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Akinlotan, M.; Bolin, J.N.; Helduser, J.; Ojinnaka, C.; Lichorad, A.; McClellan, D. Cervical cancer screening barriers and risk factor knowledge among uninsured women. J. Community Health 2017, 42, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Alfaro-Mora, Y.; Herrera, L.A.; Cáceres-Gutiérrez, R.; Andonegui-Elguera, M.A.; Dominguez-Gómez, G.; Díaz-Chávez, J. The Role of Epigenetics in Cervical Cancer. In Gynaecological Malignancies-Updates and Advances; IntechOpen: London, UK, 2019. [Google Scholar]
- Pardini, B.; De Maria, D.; Francavilla, A.; Di Gaetano, C.; Ronco, G.; Naccarati, A. MicroRNAs as markers of progression in cervical cancer: A systematic review. BMC Cancer 2018, 18, 696. [Google Scholar] [CrossRef]
- Causin, R.L.; Freitas, A.J.A.D.; Hidalgo Filho, T.; Murilo, C.; Reis, R.D.; Reis, R.M.; Marques, M.M.C. A Systematic Review of MicroRNAs Involved in Cervical Cancer Progression. Cells 2021, 10, 668. [Google Scholar] [CrossRef]
- Miao, J.; Regenstein, J.M.; Xu, D.; Zhou, D.; Li, H.; Zhang, H.; Li, C.; Qiu, J.; Chen, X. The roles of microRNA in human cervical cancer. Arch. Biochem. Biophys. 2020, 690, 108480. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Gardiner, A.S.; Board, K.F.; Monzon, F.A.; Edwards, R.P.; Khan, S.A. Human papillomavirus type 16 reduces the expression of microRNA-218 in cervical carcinoma cells. Oncogene 2008, 27, 2575–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Kinoshita, T.; Nohata, N.; Itesako, T.; Yoshino, H.; Enokida, H.; Nakagawa, M.; Shozu, M.; Seki, N. Tumor suppressive microRNA-218 inhibits cancer cell migration and invasion by targeting focal adhesion pathways in cervical squamous cell carcinoma. Int. J. Oncol. 2013, 42, 1523–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Zhai, G.; Bai, Y. Values of miR-34a and miR-218 expression in the diagnosis of cervical cancer and the prediction of prognosis. Oncol. Lett. 2018, 15, 3580–3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Tu, H.; Liang, Y.; Tang, D. MiR-218 produces anti-tumor effects on cervical cancer cells in vitro. World J. Surg. Oncol. 2018, 16, 204. [Google Scholar] [CrossRef]
- Liu, Z.; Mao, L.; Wang, L.; Zhang, H.; Hu, X. miR-218 functions as a tumor suppressor gene in cervical cancer. Mol. Med. Rep. 2020, 21, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, S.; Li, Y.; Liu, H.; Zhang, Y.; Zhang, Q. miRNA-218 regulates the proliferation and apoptosis of cervical cancer cells via targeting Gli3. Exp. Ther. Med. 2018, 16, 2433–2441. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Xiaoyun, H.; Haifeng, Q.; Jing, L.; Weixu, H.; Ruofan, D.; Jinjin, Y.; Zongji, S. MicroRNA-218 enhances the radiosensitivity of human cervical cancer via promoting radiation induced apoptosis. Int. J. Med. Sci. 2014, 11, 691. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, J.; Yuan, C.; Cui, B.; Zou, X.; Qiao, Y. High-risk human papillomavirus reduces the expression of microRNA-218 in women with cervical intraepithelial neoplasia. J. Int. Med. Res. 2010, 38, 1730–1736. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; He, Q.; Lu, Y.; Tao, F.; Zhao, L.; Ou, R. MicroRNA-218-5p inhibits cell growth and metastasis in cervical cancer via LYN/NF-κB signaling pathway. Cancer Cell Int. 2018, 18, 198. [Google Scholar] [CrossRef]
- Jia, Z.; Jiang, L.; Wang, H.; Gao, W. MiR-218 restrains proliferation of cervical cancer cells via targeted regulation of HMGB1/RAGE pathway. J. BUON 2020, 25, 2683–2689. [Google Scholar]
- Kogo, R.; How, C.; Chaudary, N.; Bruce, J.; Shi, W.; Hill, R.P.; Zahedi, P.; Yip, K.; Liu, F. The microRNA-218~ Survivin axis regulates migration, invasion, and lymph node metastasis in cervical cancer. Oncotarget 2015, 6, 1090. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Song, Q.; Zeng, R.; Li, J.; Lin, X.; Zheng, Y. MicroRNA-218 inhibits EMT, migration and invasion by targeting SFMBT1 and DCUN1D1 in cervical cancer. Oncotarget 2016, 7, 45622. [Google Scholar] [CrossRef] [Green Version]
- Kagoshima, H.; Shigesada, K.; Satake, M.; Ito, Y.; Miyoshi, H.; Ohki, M.; Pepling, M.; Gergern, P. The runt domain identifies a new family of heterometric transcriptional regulators. Trends Genet. 1993, 9, 338–341. [Google Scholar] [CrossRef]
- Rennert, J.; Coffman, J.A.; Mushegian, A.R.; Robertson, A.J. The evolution of Runx genes I. A comparative study of sequences from phylogenetically diverse model organisms. BMC Evol. Biol. 2003, 3, 4. [Google Scholar] [CrossRef]
- Komori, T. Runx2, an inducer of osteoblast and chondrocyte differentiation. Histochem. Cell Biol. 2018, 149, 313–323. [Google Scholar] [CrossRef]
- Ferrari, N.; McDonald, L.; Morris, J.S.; Cameron, E.R.; Blyth, K. RUNX2 in mammary gland development and breast cancer. J. Cell. Physiol. 2013, 228, 1137–1142. [Google Scholar] [CrossRef]
- Chang, C.H.; Fan, T.C.; Yu, J.C.; Liao, G.S.; Lin, Y.C.; Shih, A.C.C.; Li, W.H.; Yu, A.L.T. The prognostic significance of RUNX2 and miR-10a/10b and their inter-relationship in breast cancer. J. Transl. Med. 2014, 12, 257. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Yu, M.; Yin, D.; Sun, D.; Zhu, Y.; Bu, Y.; Sang, M. Impact of RUNX2 on drug-resistant human pancreatic cancer cells with p53 mutations. BMC Cancer 2018, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.; Zhong, C.; Yang, S.; Bell, A.M.; Cohen, M.B.; Roy-Burman, P. Runx2 regulates survivin expression in prostate cancer cells. Lab. Investig. 2010, 90, 222–233. [Google Scholar] [CrossRef]
- Herreño, A.M.; Ramírez, A.C.; Chaparro, V.P.; Fernandez, M.J.; Cañas, A.; Morantes, C.F.; Moreno, O.M.; Brúges, R.E.; Mejía, J.A.; Bustos, F.J.; et al. Role of RUNX2 transcription factor in epithelial mesenchymal transition in non-small cell lung cancer lung cancer: Epigenetic control of the RUNX2 P1 promoter. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2019, 41, 1010428319851014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Yang, H.; Chai, J.; Xing, L. RUNX2 as a promising therapeutic target for malignant tumors. Cancer Manag. Res. 2021, 13, 2539. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Solal, K.A.; Boregowda, R.K.; Lasfar, A. RUNX2 and the PI3K/AKT axis reciprocal activation as a driving force for tumor progression. Mol. Cancer 2015, 14, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratap, J.; Javed, A.; Languino, L.R.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. The Runx2 osteogenic transcription factor regulates matrix metalloproteinase 9 in bone metastatic cancer cells and controls cell invasion. Mol. Cell. Biol. 2005, 25, 8581–8591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Chen, L.; Wang, Y. miR-218 overexpression suppresses tumorigenesis of papillary thyroid cancer via inactivation of PTEN/PI3K/AKT pathway by targeting Runx2. OncoTargets Ther. 2018, 11, 6305. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, P.; Wu, K.; Su, L.; An, W.; Bie, Y.; Zhang, H.; Kang, H.; Jiang, E.; Zhu, W.; Yao, Y.; et al. Inhibition of human cervical cancer cell invasion by IL-37 involving runt related transcription factor 2 suppression. Ann. Transl. Med. 2019, 7, 568. [Google Scholar] [CrossRef]
- Laengsri, V.; Kerdpin, U.; Plabplueng, C.; Treeratanapiboon, L.; Nuchnoi, P. Cervical cancer markers: Epigenetics and microRNAs. Lab. Med. 2018, 49, 97–111. [Google Scholar] [CrossRef]
- He, S.; Shi, J.; Mao, J.; Luo, X.; Liu, W.; Liu, R.; Yang, F. The expression of miR-375 in prostate cancer: A study based on GEO, TCGA data and bioinformatics analysis. Pathol.-Res. Pract. 2019, 215, 152375. [Google Scholar] [CrossRef]
- Mathew, L.K.; Skuli, N.; Mucaj, V.; Lee, S.S.; Zinn, P.O.; Sathyan, P.; Imtiyaz, H.Z.; Zhang, Z.; Davuluri, R.V.; Rao, S.; et al. miR-218 opposes a critical RTK-HIF pathway in mesenchymal glioblastoma. Proc. Natl. Acad. Sci. USA 2014, 111, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Melnik, S.; Gabler, J.; Dreher, S.I.; Hecht, N.; Hofmann, N.; Großner, T.; Richter, W. MiR-218 affects hypertrophic differentiation of human mesenchymal stromal cells during chondrogenesis via targeting RUNX2, MEF2C, and COL10A1. Stem Cell Res. Ther. 2020, 11, 532. [Google Scholar] [CrossRef]
- Li, N.; Wang, L.; Tan, G.; Guo, Z.; Liu, L.; Yang, M.; He, J. MicroRNA-218 inhibits proliferation and invasion in ovarian cancer by targeting Runx2. Oncotarget 2017, 8, 91530. [Google Scholar] [CrossRef]
- Yao, R.; Yao, X.; Liu, R.; Peng, J.; Tian, T. Glucose-induced microRNA-218 suppresses the proliferation and promotes the apoptosis of human retinal pigment epithelium cells by targeting RUNX2. Biosci. Rep. 2019, 39, BSR20192580. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Yu, F.; Li, D.; Zhu, X.; Zhang, X.; Lv, Z. MicroRNA-218 regulates cisplatin (DPP) chemosensitivity in non-small cell lung cancer by targeting RUNX2. Tumor Biol. 2016, 37, 1197–1204. [Google Scholar] [CrossRef]
- Liang, R.; Cao, X.; Li, Y.; Chen, S.; Wu, Y.; Ma, Z. MicroRNA-466 regulates the proliferation, migration and invasion of the human lung cancer cells by targeting transcription factor RUNX2. J. BUON 2020, 25, 2650–2656. [Google Scholar]
- Colden, M.; Dar, A.A.; Saini, S.; Dahiya, P.V.; Shahryari, V.; Yamamura, S.; Tanaka, Y.; Stein, G.; Dahiya, R.; Majid, S. MicroRNA-466 inhibits tumor growth and bone metastasis in prostate cancer by direct regulation of osteogenic transcription factor RUNX2. Cell Death Dis. 2018, 8, e2572. [Google Scholar] [CrossRef]
- Zhou, L.L.; Shen, Y.; Gong, J.M.; Sun, P.; Sheng, J.H. MicroRNA-466 with tumor markers for cervical cancer screening. Oncotarget 2017, 8, 70821. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Q.; Lu, J.T.; Tan, C.C.; Wang, Q.S.; Feng, Y.M. RUNX2 promotes breast cancer bone metastasis by increasing integrin α5-mediated colonization. Cancer Lett. 2016, 380, 78–86. [Google Scholar] [CrossRef]
- Ben, W.; Yang, Y.; Yuan, J.; Sun, J.; Huang, M.; Zhang, D.; Zheng, J. Human papillomavirus 16 E6 modulates the expression of host microRNAs in cervical cancer. Taiwan. J. Obstet. Gynecol. 2015, 54, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Song, L.; Yao, H.; Zhang, L.; Xu, D.; Gao, F.; Li, Q. MiR-375 is epigenetically downregulated by HPV-16 E6 mediated DNMT1 upregulation and modulates EMT of cervical cancer cells by suppressing lncRNA MALAT1. PLoS ONE 2016, 11, e0163460. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Wences, H.; Martínez-Carrillo, D.N.; Peralta-Zaragoza, O.; Campos-Viguri, G.E.; Hernández-Sotelo, D.; Jiménez-López, M.A.; Muñoz-Camacho, J.G.; Garzón-Barrientos, V.H.; Illades-Aguiar, B.; Fernández-Tilapa, G. Methylation and expression of miRNAs in precancerous lesions and cervical cancer with HPV16 infection. Oncol. Rep. 2016, 35, 2297–2305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Liu, L.; Zhang, L.; He, X.; Xu, X.; Lu, Y.; Li, F. Runx2 is required for activity of CD44+/CD24-/low breast cancer stem cell in breast cancer development. Am. J. Transl. Res. 2020, 12, 2305. [Google Scholar]
- Kim, B.; Kim, H.; Jung, S.; Moon, A.; Noh, D.Y.; Lee, Z.H.; Kim, H.J.; Kim, H.H. A CTGF-RUNX2-RANKL axis in breast and prostate cancer cells promotes tumor progression in bone. J. Bone Miner. Res. 2020, 35, 155–166. [Google Scholar] [CrossRef]
- He, F.; Matsumoto, Y.; Asano, Y.; Yamamura, Y.; Katsuyama, T.; La Rose, J.; Tomonobu, N.; Komalarasi, N.L.G.Y.; Sakaguchi, M.; Rottapel, R.; et al. RUNX2 Phosphorylation by Tyrosine Kinase ABL Promotes Breast Cancer Invasion. Front. Oncol. 2021, 11, 1929. [Google Scholar]
- Bai, Y.; Yang, Y.; Yan, Y.; Zhong, J.; Blee, A.M.; Pan, Y.; Ma, T.; Karnes, R.J.; Jimenez, R.; Xu, W.; et al. RUNX2 overexpression and PTEN haploinsufficiency cooperate to promote CXCR7 expression and cellular trafficking, AKT hyperactivation and prostate tumorigenesis. Theranostics 2019, 9, 3459. [Google Scholar] [CrossRef]
- Guo, Z.; Zhou, K.; Wang, Q.; Huang, Y.; Ji, J.; Peng, Y.; Zhang, X.; Zheng, T.; Zhang, Z.; Chong, D.; et al. The transcription factor RUNX2 fuels YAP1 signaling and gastric cancer tumorigenesis. Cancer Sci. 2021, 112, 3533. [Google Scholar] [CrossRef] [PubMed]
- Nishio, M.; To, Y.; Maehama, T.; Aono, Y.; Otani, J.; Hikasa, H.; Kitagaka, A.; Mimori, K.; Sasaki, T.; Nishina, H.; et al. Endogenous YAP1 activation drives immediate onset of cervical carcinoma in situ in mice. Cancer Sci. 2020, 111, 3576–3587. [Google Scholar] [CrossRef]
- Lu, H.; Jiang, T.; Ren, K.; Li, Z.L.; Ren, J.; Wu, G.; Han, X. RUNX2 plays an oncogenic role in esophageal carcinoma by activating the PI3K/AKT and ERK signaling pathways. Cell. Physiol. Biochem. 2018, 49, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Tandon, M.; Chen, Z.; Pratap, J. Runx2 activates PI3K/Akt signaling via mTORC2 regulation in invasive breast cancer cells. Breast Cancer Res. 2014, 16, R16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Cao, W.; Chellaiah, M.A. Integrin αvβ3 and CD44 pathways in metastatic prostate cancer cells support osteoclastogenesis via a Runx2/Smad 5/receptor activator of NF-κB ligand signaling axis. Mol. Cancer 2012, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yan, H.; Cheng, D.; Xu, L.; Shen, T.; Chen, Y.; Han, R.; Xue, Y. Novel lncRNA LINC01614 Facilitates Bladder Cancer Proliferation, Migration and Invasion Through the miR-217/RUNX2/Wnt/β-Catenin Axis. Cancer Manag. Res. 2021, 13, 8387. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhou, F. Comprehensive Analysis of RUNX and TGF-β Mediated Regulation of Immune Cell Infiltration in Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 730380. [Google Scholar] [CrossRef] [PubMed]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.D.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA–target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2012, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Zhang, Y.; Zhu, M.; Liu, S.; Wang, X. miRNA expression profiles of HPV-infected patients with cervical cancer in the Uyghur population in China. PLoS ONE 2016, 11, e0164701. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Kuick, R.; Nan, B.; Ota, I.; Weiss, S.J.; Trimble, C.L.; Fearon, E.R.; Cho, K.R. Gene expression analysis of preinvasive and invasive cervical squamous cell carcinomas identifies HOXC10 as a key mediator of invasion. Cancer Res. 2007, 67, 10163–10172. [Google Scholar] [CrossRef] [Green Version]
- Lánczky, A.; Győrffy, B. Web-based survival analysis tool tailored for medical research (KMplot): Development and implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Nagy, Á.; Munkácsy, G.; Győrffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11, 6047. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 118 Genes Predicted to Be Targeted by miR-218-5p | |||
|---|---|---|---|
| ACTN1 | GLCE | NFATC3 | SH3GL1 |
| ANKRD27 | GNAI2 | NRAS | SH3KBP1 |
| ANKRD52 | GNAS | NUFIP2 | SHMT1 |
| ARPP19 | GNB1 | OTUD7B | SHOC2 |
| ATXN1 | HECTD2 | PAIP2 | SIK2 |
| BCL11B | HNRNPA3 | PDGFRA | SLAIN2 |
| BCL9 | HP1BP3 | PHC3 | SLC38A1 |
| BIRC6 | ITM2C | PKP4 | SLC45A3 |
| BMI1 | KIAA1522 | PPP1CC | SNX4 |
| CACUL1 | KIF21B | PPP2R2A | SOST |
| CCDC6 | KIRREL3 | PRKG1 | SPAG9 |
| CDH2 | KIT | PTP4A1 | SREK1 |
| CNTNAP2 | KLF12 | PUM2 | SRSF10 |
| COL1A1 | KLF9 | PURB | STAM2 |
| CUL3 | KLHL13 | PRLR | THOC2 |
| DAZAP1 | KMT2A | RARA | TOB1 |
| DCBLD2 | LARP4B | RBM18 | TPD52 |
| DCP2 | LASP1 | RET | TTYH3 |
| DDX6 | LGR4 | RHOQ | UBE2H |
| DOCK9 | LMNB1 | RICTOR | UBN2 |
| DUSP18 | LRIG1 | RNF219 | USP34 |
| ELL2 | LRRFIP1 | RNF38 | VAT1 |
| FAM214A | LYPD6B | ROBO1 | VOPP1 |
| FAM217B | MAFG | RPP25 | WASF3 |
| FBN2 | MARCKS | RPS6KA3 | WIPF2 |
| FBXO41 | MBNL1 | RPS6KB1 | ZFX |
| FYCO1 | MBNL2 | RUNX2 | ZFYVE26 |
| GAB2 | MITF | SATB2 | ZNF609 |
| GFPT1 | NAA15 | SEPHS1 | |
| GJA1 | NACC1 | SETBP1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-De la Rosa, M.I.; Jiménez-Wences, H.; Alarcón-Millán, J.; Romero-López, M.J.; Castañón-Sánchez, C.A.; Salmerón-Bárcenas, E.G.; Fernández-Tilapa, G. miR-218-5p/RUNX2 Axis Positively Regulates Proliferation and Is Associated with Poor Prognosis in Cervical Cancer. Int. J. Mol. Sci. 2022, 23, 6993. https://doi.org/10.3390/ijms23136993
Cruz-De la Rosa MI, Jiménez-Wences H, Alarcón-Millán J, Romero-López MJ, Castañón-Sánchez CA, Salmerón-Bárcenas EG, Fernández-Tilapa G. miR-218-5p/RUNX2 Axis Positively Regulates Proliferation and Is Associated with Poor Prognosis in Cervical Cancer. International Journal of Molecular Sciences. 2022; 23(13):6993. https://doi.org/10.3390/ijms23136993
Chicago/Turabian StyleCruz-De la Rosa, Merlin Itsel, Hilda Jiménez-Wences, Judit Alarcón-Millán, Manuel Joaquín Romero-López, Carlos Alberto Castañón-Sánchez, Eric Genaro Salmerón-Bárcenas, and Gloria Fernández-Tilapa. 2022. "miR-218-5p/RUNX2 Axis Positively Regulates Proliferation and Is Associated with Poor Prognosis in Cervical Cancer" International Journal of Molecular Sciences 23, no. 13: 6993. https://doi.org/10.3390/ijms23136993