
A Transcriptomic Response to Lactiplantibacillus plantarum-KCC48 against High-Fat Diet-Induced Fatty Liver Diseases in Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
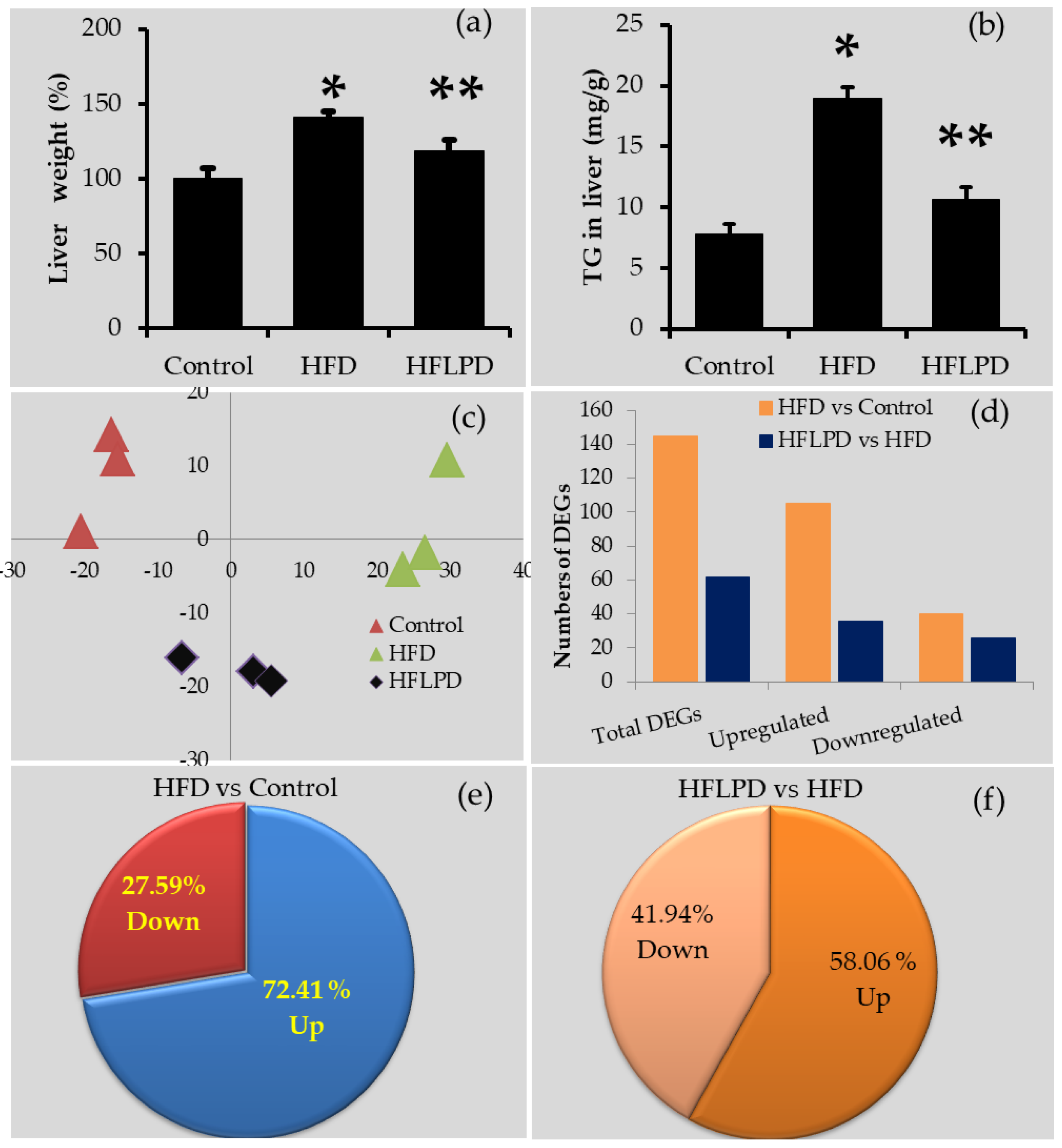
2.1. Transcriptome Validation ASSESSMENT
2.2. Overview of Transcriptome Changes
2.3. Location of Differentially Expressed Genes in Liver
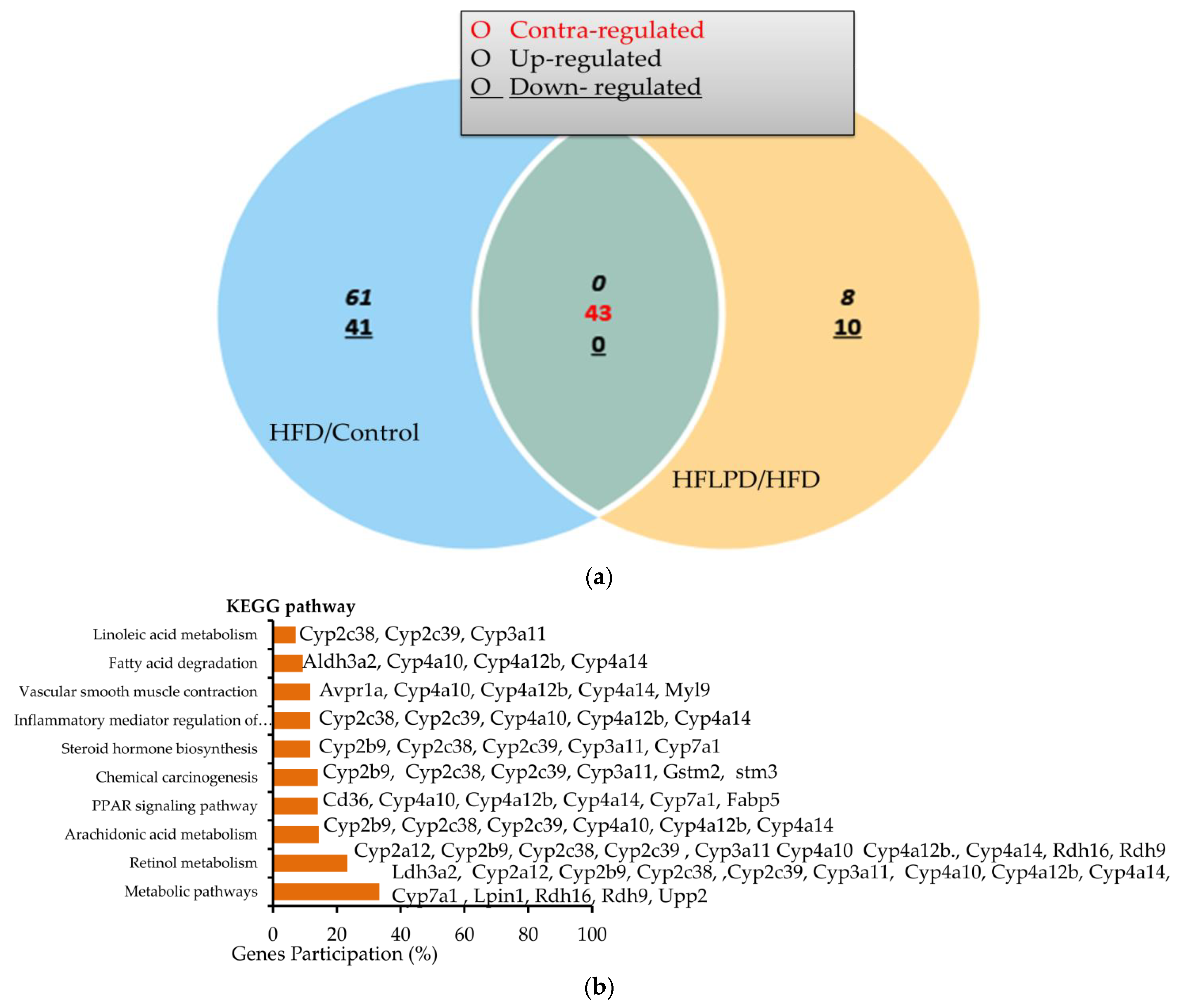
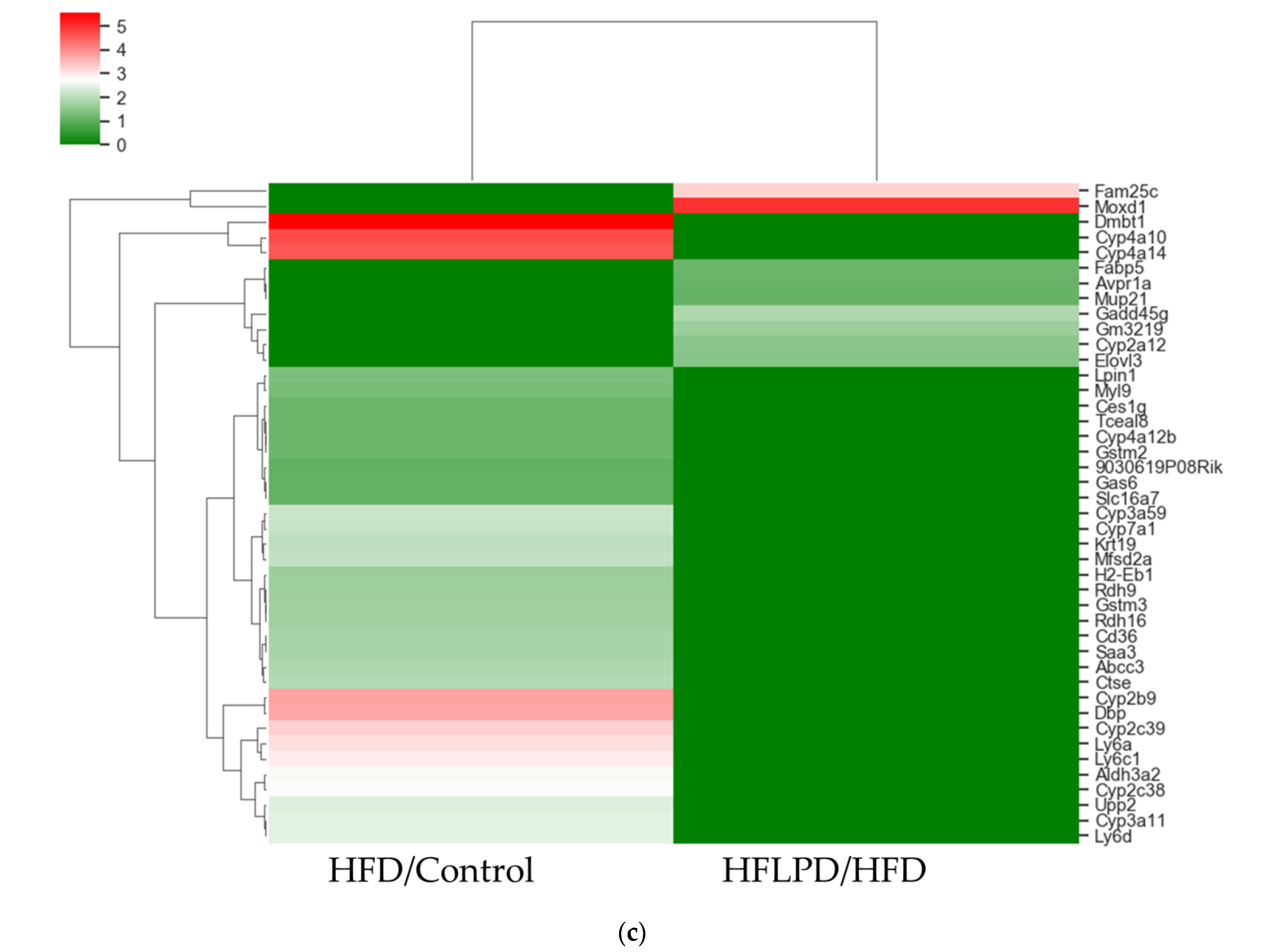
2.4. Common Differentially Expressed Genes between HFD and HFLPD Groups
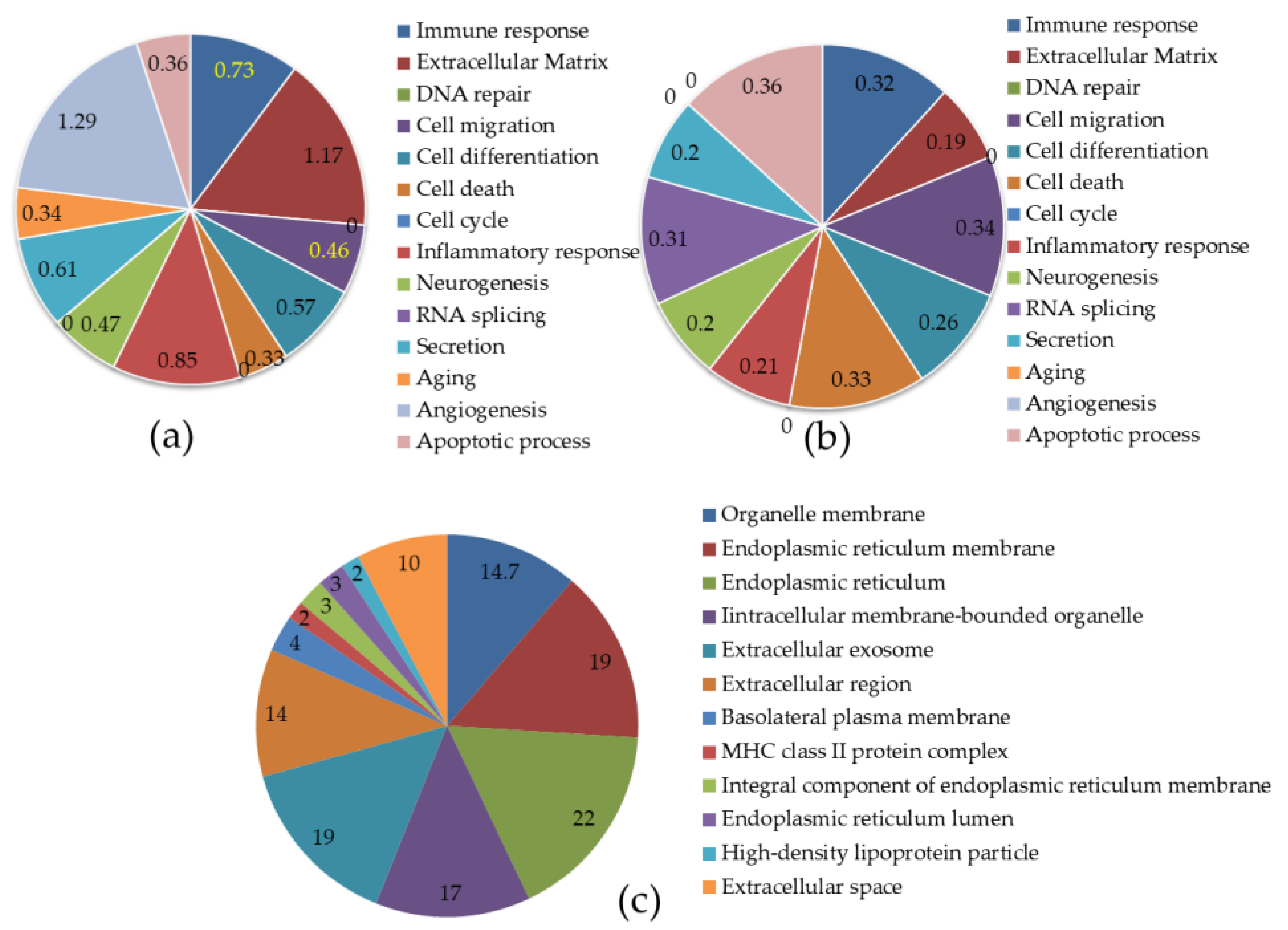
2.5. Functional Characterization of DEGs in HFD and HFLPD
2.6. KEGG Signaling Enrichment Analysis for DEGs in HFD and HFLPD
3. Discussion
4. Materials and Methods
4.1. Diet and Lactiplantibacillus plantarum-KCC48
4.2. Animals and Probiotic Diet Production
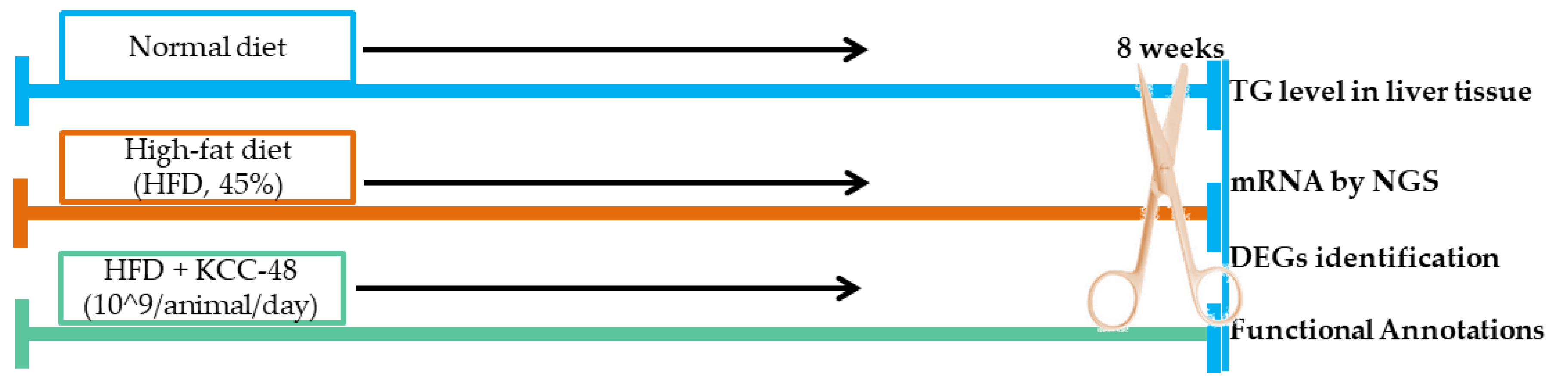
4.3. Experimental Design
4.4. Liver Triglyceride Quantification
4.5. RNA Extraction
4.6. Library Preparation and Sequencing
4.7. Data Analysis and Removal of Low Quality Reads
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Godoy-Matos, A.F.; Silva Júnior, W.S.; Valerio, C.M. NAFLD as a continuum: From obesity to metabolic syndrome and diabetes. Diabetol. Metab. Syndr. 2020, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- NCD Risk Factor Collaboration. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.T.; Makishima, M.; Repa, J.J.; Schoonjans, K.; Kerr, T.A.; Auwerx, J.; Mangelsdorf, D.J. Molecular basis for feedback regulation of bile acid synthesis by nuclear receptors. Mol. Cell 2000, 6, 507–515. [Google Scholar] [CrossRef]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid Res. 2009, 48, 1–26. [Google Scholar] [CrossRef]
- Sullivan, S. Implications of diet on nonalcoholic fatty liver disease. Curr. Opin. Gastroenterol. 2010, 26, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, F.; Ishida, Y.; Aihara, K.; Sawada, D.; Ashida, N.; Sugawara, T.; Aoki, Y.; Takehara, I.; Takano, K.; Fujiwara, S. Effect of fragmented Lactobacillus amylovorus CP1563 on lipid metabolism in overweight and mildly obese individuals: A randomized controlled trial. Microb. Ecol. Health Dis. 2016, 27, 30312. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, M.; Ogawa, A.; Higurashi, S.; Kadooka, Y. Anti-obesity effect of Lactobacillus gasseri SBT2055 accompanied by inhibition of pro-inflammatory gene expression in the visceral adipose tissue in diet-induced obese mice. Eur. J. Nutr. 2014, 53, 599–606. [Google Scholar] [CrossRef]
- Choi, W.J.; Dong, H.J.; Jeong, H.U.; Ryu, D.W.; Song, S.M.; Kim, Y.R.; Jung, H.H.; Kim, T.H.; Kim, Y.-H. Lactobacillus plantarum LMT1-48 exerts anti-obesity effect in high-fat diet-induced obese mice by regulating expression of lipogenic genes. Sci. Rep. 2020, 10, 869. [Google Scholar] [CrossRef] [Green Version]
- Soundharrajan, I.; Kuppusamy, P.; Srisesharam, S.; Lee, J.C.; Sivanesan, R.; Kim, D.; Choi, K.C. Positive metabolic effects of selected probiotic bacteria on diet-induced obesity in mice are associated with improvement of dysbiotic gut microbiota. FASEB J. 2020, 34, 12289–12307. [Google Scholar] [CrossRef]
- Tang, S.; Liu, J.; Xu, C.; Shang, D.; Chen, H.; Zhang, G. Effects of probiotics on the improvement and regulation of intestinal barrier dysfunction and immune imbalance in intra-abdominal infections (Review). Int. J. Funct. Nutr. 2021, 2, 12. [Google Scholar] [CrossRef]
- Jang, H.R.; Park, H.-J.; Kang, D.; Chung, H.; Nam, M.H.; Lee, Y.; Park, J.-H.; Lee, H.-Y. A protective mechanism of probiotic Lactobacillus against hepatic steatosis via reducing host intestinal fatty acid absorption. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-J.; Park, S.H.; Sin, H.-S.; Jang, S.-H.; Lee, S.-W.; Kim, S.-Y.; Kwon, B.; Yu, K.-Y.; Kim, S.Y.; Yang, D.K. Hypocholesterolemic Effects of Probiotic Mixture on Diet-Induced Hypercholesterolemic Rats. Nutrients 2017, 9, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arellano-García, L.; Portillo, M.P.; Martínez, J.A.; Milton-Laskibar, I. Usefulness of Probiotics in the Management of NAFLD: Evidence and Involved Mechanisms of Action from Preclinical and Human Models. Int. J. Mol. Sci. 2022, 23, 3167. [Google Scholar] [CrossRef]
- Han, R.; Ma, J.; Li, H. Mechanistic and therapeutic advances in non-alcoholic fatty liver disease by targeting the gut microbiota. Front. Med. 2018, 12, 645–657. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Li, X.; Huang, Y.; Song, L.; Xiao, Y.; Lu, S.; Xu, J.; Li, J.; Ren, Z. Lactobacillus plantarum prevents obesity via modulation of gut microbiota and metabolites in high-fat feeding mice. J. Funct. Foods 2020, 73, 104103. [Google Scholar] [CrossRef]
- Takemura, N.; Okubo, T.; Sonoyama, K. Lactobacillus plantarum strain No. 14 reduces adipocyte size in mice fed high-fat diet. Exp. Biol. Med. 2010, 235, 849–856. [Google Scholar] [CrossRef]
- Srivastava, A.; George, J.; Karuturi, R.K.M. Transcriptome Analysis. In Encyclopedia of Bioinformatics and Computational Biology; Ranganathan, S., Gribskov, M., Nakai, K., Schönbach, C., Eds.; Academic Press: Oxford, UK, 2019; pp. 792–805. [Google Scholar] [CrossRef]
- Fang, T.J.; Guo, J.T.; Lin, M.K.; Lee, M.S.; Chen, Y.L.; Lin, W.H. Protective effects of Lactobacillus plantarum against chronic alcohol-induced liver injury in the murine model. Appl. Microbiol. Biotechnol. 2019, 103, 8597–8608. [Google Scholar] [CrossRef]
- Feng, D.; Lazar, M.A. Clocks, metabolism, and the epigenome. Mol. Cell 2012, 47, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Tahara, Y.; Shibata, S. Chronobiology and nutrition. Neuroscience 2013, 253, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M. Pathology of nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Cao, F.; Lai, S.; Zhuge, H.; Chang, K.; Valencak, T.G.; Liu, J.; Li, S.; Ren, D. Lactobacillus plantarum ZY08 relieves chronic alcohol-induced hepatic steatosis and liver injury in mice via restoring intestinal flora homeostasis. Food Res. Int. 2022, 157, 111259. [Google Scholar] [CrossRef]
- Shukla, P.K.; Meena, A.S.; Manda, B.; Gomes-Solecki, M.; Dietrich, P.; Dragatsis, I.; Rao, R. Lactobacillus plantarum prevents and mitigates alcohol-induced disruption of colonic epithelial tight junctions, endotoxemia, and liver damage by an EGF receptor-dependent mechanism. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, fj201800351R. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Lee, Y.-S.; Kim, S.M.; Park, G.-S.; Lee, Y.H.; Jeong, D.Y.; Kang, J.; Lee, H.-J. Beneficial Effects of Lactobacillus plantarum Strains on Non-Alcoholic Fatty Liver Disease in High Fat/High Fructose Diet-Fed Rats. Nutrients 2020, 12, 542. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.Y.; Shin, M.J.; Youn, G.S.; Yoon, S.J.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Han, S.H.; Kim, B.K.; Lee, D.Y.; et al. Lactobacillus attenuates progression of nonalcoholic fatty liver disease by lowering cholesterol and steatosis. Clin. Mol. Hepatol. 2021, 27, 110–124. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef]
- Lau, J.K.C.; Zhang, X.; Yu, J. Animal models of non-alcoholic fatty liver disease: Current perspectives and recent advances. J. Pathol. 2017, 241, 36–44. [Google Scholar] [CrossRef]
- Nseir, W.; Hellou, E.; Assy, N. Role of diet and lifestyle changes in nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 9338–9344. [Google Scholar] [CrossRef]
- Sheedfar, F.; Sung, M.M.; Aparicio-Vergara, M.; Kloosterhuis, N.J.; Miquilena-Colina, M.E.; Vargas-Castrillón, J.; Febbraio, M.; Jacobs, R.L.; de Bruin, A.; Vinciguerra, M.; et al. Increased hepatic CD36 expression with age is associated with enhanced susceptibility to nonalcoholic fatty liver disease. Aging 2014, 6, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, A.; Miyazaki, T.; Iwamoto, J.; Hirayama, T.; Morishita, Y.; Monma, T.; Ueda, H.; Mizuno, S.; Sugiyama, F.; Takahashi, S.; et al. Regulation of bile acid metabolism in mouse models with hydrophobic bile acid composition. J. Lipid Res. 2020, 61, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Hoek-van den Hil, E.F.; van Schothorst, E.M.; van der Stelt, I.; Swarts, H.J.; van Vliet, M.; Amolo, T.; Vervoort, J.J.; Venema, D.; Hollman, P.C.; Rietjens, I.M.; et al. Direct comparison of metabolic health effects of the flavonoids quercetin, hesperetin, epicatechin, apigenin and anthocyanins in high-fat-diet-fed mice. Genes Nutr. 2015, 10, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, A.; Trac, C.; Du, J.; Natarajan, R.; Schones, D.E. Persistent Chromatin Modifications Induced by High Fat Diet. J. Biol. Chem. 2016, 291, 10446–10455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteng, A.B.; Higuchi, S.; Banks, A.S.; Haeusler, R.A. Cyp2c-deficiency depletes muricholic acids and protects against high-fat diet-induced obesity in male mice but promotes liver damage. Mol. Metab. 2021, 53, 101326. [Google Scholar] [CrossRef]
- Xiang, L.; Jiao, Y.; Qian, Y.; Li, Y.; Mao, F.; Lu, Y. Comparison of hepatic gene expression profiles between three mouse models of Nonalcoholic Fatty Liver Disease. Genes Dis. 2022, 9, 201–215. [Google Scholar] [CrossRef]
- Ahn, J.; Cho, I.; Kim, S.; Kwon, D.; Ha, T. Dietary resveratrol alters lipid metabolism-related gene expression of mice on an atherogenic diet. J. Hepatol. 2008, 49, 1019–1028. [Google Scholar] [CrossRef]
- He, Y.; Yang, T.; Du, Y.; Qin, L.; Ma, F.; Wu, Z.; Ling, H.; Yang, L.; Wang, Z.; Zhou, Q.; et al. High fat diet significantly changed the global gene expression profile involved in hepatic drug metabolism and pharmacokinetic system in mice. Nutr. Metab. 2020, 17, 37. [Google Scholar] [CrossRef]
- Kirpich, I.A.; Gobejishvili, L.N.; Bon Homme, M.; Waigel, S.; Cave, M.; Arteel, G.; Barve, S.S.; McClain, C.J.; Deaciuc, I.V. Integrated hepatic transcriptome and proteome analysis of mice with high-fat diet-induced nonalcoholic fatty liver disease. J. Nutr. Biochem. 2011, 22, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Ning, M.; Jeong, H. High-Fat Diet Feeding Alters Expression of Hepatic Drug-Metabolizing Enzymes in Mice. Drug Metab. Dispos. Biol. Fate Chem. 2017, 45, 707–711. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, S.; Zhou, Y.; Su, W.; Ruan, X.; Wang, B.; Zheng, F.; Warner, M.; Gustafsson, J.; Guan, Y. Ablation of cytochrome P450 omega-hydroxylase 4A14 gene attenuates hepatic steatosis and fibrosis. Proc. Natl. Acad. Sci. USA 2017, 114, 3181–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatuphonprasert, W.; Nemoto, N.; Sakuma, T.; Jarukamjorn, K. Modulations of cytochrome P450 expression in diabetic mice by berberine. Chem.-Biol. Interact. 2012, 196, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, K.; Takagi, S.; Sugatani, J.; Miwa, M. Changes in the expression of cytochromes P450 and nuclear receptors in the liver of genetically diabetic db/db mice. Biol. Pharm. Bull. 2006, 29, 1634–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez, A.; Leclercq, I.; Farrell, G.C.; Robertson, G. Altered expression of hepatic CYP2E1 and CYP4A in obese, diabetic ob/ob mice, and fa/fa Zucker rats. Biochem. Biophys. Res. Commun. 1999, 255, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xiaoling, G.; Pingting, L.; Shuqiang, L.; Yuaner, Z. Chronic unpredictable mild stress combined with a high-fat diets aggravates atherosclerosis in rats. Lipids Health Dis. 2014, 13, 77. [Google Scholar] [CrossRef] [Green Version]
- Jiao, N.; Baker, S.S.; Chapa-Rodriguez, A.; Liu, W.; Nugent, C.A.; Tsompana, M.; Mastrandrea, L.; Buck, M.J.; Baker, R.D.; Genco, R.J.; et al. Suppressed hepatic bile acid signalling despite elevated production of primary and secondary bile acids in NAFLD. Gut 2018, 67, 1881–1891. [Google Scholar] [CrossRef]
- Boyer, J.L. Bile formation and secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [CrossRef] [Green Version]
- La Frano, M.R.; Hernandez-Carretero, A.; Weber, N.; Borkowski, K.; Pedersen, T.L.; Osborn, O.; Newman, J.W. Diet-induced obesity and weight loss alter bile acid concentrations and bile acid-sensitive gene expression in insulin target tissues of C57BL/6J mice. Nutr. Res. 2017, 46, 11–21. [Google Scholar] [CrossRef]
- More, V.R.; Slitt, A.L. Alteration of hepatic but not renal transporter expression in diet-induced obese mice. Drug Metab. Dispos. Biol. Fate Chem. 2011, 39, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Amr, K.; El-Bassyouni, H.T.; Ismail, S.; Youness, E.; El-Daly, S.M.; Ebrahim, A.Y.; El-Kamah, G. Genetic assessment of ten Egyptian patients with Sjögren-Larsson syndrome: Expanding the clinical spectrum and reporting a novel ALDH3A2 mutation. Arch. Dermatol. Res. 2019, 311, 721–730. [Google Scholar] [CrossRef]
- Kitamura, T.; Takagi, S.; Naganuma, T.; Kihara, A. Mouse aldehyde dehydrogenase ALDH3B2 is localized to lipid droplets via two C-terminal tryptophan residues and lipid modification. Biochem. J. 2015, 465, 79–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, N.K.; Nookaew, I.; Sandberg, A.-S.; Gabrielsson, B.G. Eicosapentaenoic and docosahexaenoic acid-enriched high fat diet delays the development of fatty liver in mice. Lipids Health Dis. 2015, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, V.J.; Silver, D.L. The lipid droplet as a potential therapeutic target in NAFLD. Semin. Liver Dis. 2013, 33, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shibamoto, T.; Kuda, Y.; Shinomiya, S.; Kurata, Y. The responses of the hepatic and splanchnic vascular beds to vasopressin in rats. Biomed. Res. 2012, 33, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Dünser, M.W.; Westphal, M. Arginine vasopressin in vasodilatory shock: Effects on metabolism and beyond. Curr. Opin. Anaesthesiol. 2008, 21, 122–127. [Google Scholar] [CrossRef]
- Lede, V.; Meusel, A.; Garten, A.; Popkova, Y.; Penke, M.; Franke, C.; Ricken, A.; Schulz, A.; Kiess, W.; Huster, D.; et al. Altered hepatic lipid metabolism in mice lacking both the melanocortin type 4 receptor and low density lipoprotein receptor. PLoS ONE 2017, 12, e0172000. [Google Scholar] [CrossRef]
- Ma, X.; Bodary, P.F. Abstract 296: Decreased Hepatic Avpr1a Gene Expression and Elevated Serum Bile Acid in Mouse Model of Metabolic Syndrome. Arter. Thromb. Vasc. Biol. 2012, 32, A296. [Google Scholar] [CrossRef]
- Rada, P.; González-Rodríguez, Á.; García-Monzón, C.; Valverde, Á.M. Understanding lipotoxicity in NAFLD pathogenesis: Is CD36 a key driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef]
- Houben, T.; Oligschlaeger, Y.; Hendrikx, T.; Bitorina, A.V.; Walenbergh, S.M.A.; van Gorp, P.J.; Gijbels, M.J.J.; Friedrichs, S.; Plat, J.; Schaap, F.G.; et al. Cathepsin D regulates lipid metabolism in murine steatohepatitis. Sci. Rep. 2017, 7, 3494. [Google Scholar] [CrossRef] [Green Version]
- Yadati, T.; Houben, T.; Bitorina, A.; Oligschlaeger, Y.; Gijbels, M.J.; Mohren, R.; Lütjohann, D.; Khurana, P.; Goyal, S.; Kulkarni, A.; et al. Inhibition of Extracellular Cathepsin D Reduces Hepatic Lipid Accumulation and Leads to Mild Changes in Inflammationin NASH Mice. Front. Immunol. 2021, 12, 675535. [Google Scholar] [CrossRef]
- Mollenhauer, J.; Wiemann, S.; Scheurlen, W.; Korn, B.; Hayashi, Y.; Wilgenbus, K.K.; von Deimling, A.; Poustka, A. DMBT1, a new member of the SRCR superfamily, on chromosome 10q25.3-26.1 is deleted in malignant brain tumours. Nat. Genet. 1997, 17, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zhang, X.; Cao, T.; Wang, J.; Li, C.; Yue, L.; Niu, K.; Shen, Y.; Ma, G.; Chen, F. Hepatic Transcriptome Analysis Revealing the Molecular Pathogenesis of Type 2 Diabetes Mellitus in Zucker Diabetic Fatty Rats. Front. Endocrinol. 2020, 11, 565858. [Google Scholar] [CrossRef] [PubMed]
- Gachon, F.; Olela, F.F.; Schaad, O.; Descombes, P.; Schibler, U. The circadian PAR-domain basic leucine zipper transcription factors DBP, TEF, and HLF modulate basal and inducible xenobiotic detoxification. Cell Metab. 2006, 4, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, C.; Ushijima, K.; Ando, H.; Kitamura, H.; Horiguchi, M.; Akita, T.; Yamashita, C.; Fujimura, A. Induction of Dbp by a histone deacetylase inhibitor is involved in amelioration of insulin sensitivity via adipocyte differentiation in ob/ob mice. Chronobiol. Int. 2019, 36, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Petta, S.; Valenti, L.; Marra, F.; Grimaudo, S.; Tripodo, C.; Bugianesi, E.; Cammà, C.; Cappon, A.; Di Marco, V.; Di Maira, G.; et al. MERTK rs4374383 polymorphism affects the severity of fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 682–690. [Google Scholar] [CrossRef]
- Do, G.M.; Oh, H.Y.; Kwon, E.Y.; Cho, Y.Y.; Shin, S.K.; Park, H.J.; Jeon, S.M.; Kim, E.; Hur, C.G.; Park, T.S.; et al. Long-term adaptation of global transcription and metabolism in the liver of high-fat diet-fed C57BL/6J mice. Mol. Nutr. Food Res. 2011, 55 (Suppl. S2), S173–S185. [Google Scholar] [CrossRef]
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular Reaction in Liver Diseases: Pathological Mechanisms and Translational Significances. Hepatology 2019, 69, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wang, X.; Jia, F.; Tanaka, N.; Kimura, T.; Nakajima, T.; Sato, Y.; Moriya, K.; Koike, K.; Gonzalez, F.J.; et al. A trans-fatty acid-rich diet promotes liver tumorigenesis in HCV core gene transgenic mice. Carcinogenesis 2020, 41, 159–170. [Google Scholar] [CrossRef]
- Chen, H.C.; Frissora, F.; Durbin, J.E.; Muthusamy, N. Activation induced differential regulation of stem cell antigen-1 (Ly-6A/E) expression in murine B cells. Cell. Immunol. 2003, 225, 42–52. [Google Scholar] [CrossRef]
- Ge, Q.; Zhang, S.; Chen, L.; Tang, M.; Liu, L.; Kang, M.; Gao, L.; Ma, S.; Yang, Y.; Lv, P.; et al. Mulberry Leaf Regulates Differentially Expressed Genes in Diabetic Mice Liver Based on RNA-Seq Analysis. Front. Physiol. 2018, 9, 1051. [Google Scholar] [CrossRef] [Green Version]
- Berger, J.H.; Charron, M.J.; Silver, D.L. Major facilitator superfamily domain-containing protein 2a (MFSD2A) has roles in body growth, motor function, and lipid metabolism. PLoS ONE 2012, 7, e50629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryaboshapkina, M.; Hammar, M. Human hepatic gene expression signature of non-alcoholic fatty liver disease progression, a meta-analysis. Sci. Rep. 2017, 7, 12361. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Luo, L.; Chen, X. The landscape of prognostic and immunological role of myosin light chain 9 (MYL9) in human tumors. Immun. Inflamm. Dis. 2022, 10, 241–254. [Google Scholar] [CrossRef] [PubMed]
- den Hartigh, L.J.; Wang, S.; Goodspeed, L.; Ding, Y.; Averill, M.; Subramanian, S.; Wietecha, T.; O’Brien, K.D.; Chait, A. Deletion of Serum Amyloid A3 Improves High Fat High Sucrose Diet-Induced Adipose Tissue Inflammation and Hyperlipidemia in Female Mice. PLoS ONE 2014, 9, e108564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wopereis, S.; Radonjic, M.; Rubingh, C.; van Erk, M.; Smilde, A.; van Duyvenvoorde, W.; Cnubben, N.; Kooistra, T.; van Ommen, B.; Kleemann, R. Identification of prognostic and diagnostic biomarkers of glucose intolerance in ApoE3Leiden mice. Physiol. Genom. 2012, 44, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, Y.; Pizzorno, G.; Le, T.T. Chronic Uridine Administration Induces Fatty Liver and Pre-Diabetic Conditions in Mice. PLoS ONE 2016, 11, e0146994. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Leffert, J.J.; McCabe, J.; Kim, B.; Pizzorno, G. Abnormalities in Uridine Homeostatic Regulation and Pyrimidine Nucleotide Metabolism as a Consequence of the Deletion of the Uridine Phosphorylase Gene*. J. Biol. Chem. 2005, 280, 21169–21175. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Repa, J.J.; Inoue, Y.; Hayhurst, G.P.; Gonzalez, F.J.; Mangelsdorf, D.J. Identification of a Liver-Specific Uridine Phosphorylase that Is Regulated by Multiple Lipid-Sensing Nuclear Receptors. Mol. Endocrinol. 2004, 18, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: https://www.R-project.org (accessed on 1 April 2022).
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Gene Symbol | Gene Name | Fold Changes | |
|---|---|---|---|---|
| HFD/Control | HFLPD/HFD | |||
| 1 | 9030619P08Rik | lymphocyte antigen 6 complex pseudogene (9030619P08Rik) | 2.094 | 0.493 |
| 2 | Abcc3 | ATP-binding cassette, sub-family C (CFTR/MRP), member 3 (Abcc3) | 3.739 | 0.373 |
| 3 | Aldh3a2 | aldehyde dehydrogenase family 3, subfamily A2 (Aldh3a2) | 6.414 | 0.448 |
| 4 | Avpr1a | arginine vasopressin receptor 1A (Avpr1a) | 0.454 | 2.216 |
| 5 | Cd36 | CD36 antigen (Cd36) | 3.558 | 0.375 |
| 6 | Ces1g | carboxylesterase 1G (Ces1g) | 2.228 | 0.356 |
| 7 | Ctse | cathepsin E (Ctse) | 3.918 | 0.382 |
| 8 | Cyp2a12 | cytochrome P450, family 2, subfamily a, polypeptide 12 (Cyp2a12) | 0.427 | 2.902 |
| 9 | Cyp2b9 | cytochrome P450, family 2, subfamily b, polypeptide 9 (Cyp2b9) | 13.813 | 0.351 |
| 10 | Cyp2c38 | cytochrome P450, family 2, subfamily c, polypeptide 38 (Cyp2c38) | 6.591 | 0.273 |
| 11 | Cyp2c39 | cytochrome P450, family 2, subfamily c, polypeptide 39 (Cyp2c39) | 9.718 | 0.243 |
| 12 | Cyp3a11 | cytochrome P450, family 3, subfamily a, polypeptide 11 (Cyp3a11) | 5.499 | 0.460 |
| 13 | Cyp3a59 | cytochrome P450, family 3, subfamily a, polypeptide 59 (Cyp3a59) | 4.626 | 0.397 |
| 14 | Cyp4a10 | cytochrome P450, family 4, subfamily a, polypeptide 10 (Cyp4a10) | 26.691 | 0.186 |
| 15 | Cyp4a12b | cytochrome P450, family 4, subfamily a, polypeptide 12B (Cyp4a12b) | 2.282 | 0.456 |
| 16 | Cyp4a14 | cytochrome P450, family 4, subfamily a, polypeptide 14 (Cyp4a14) | 24.394 | 0.180 |
| 17 | Cyp7a1 | cytochrome P450, family 7, subfamily a, polypeptide 1 (Cyp7a1) | 4.482 | 0.454 |
| 18 | Dbp | D site albumin promoter binding protein (Dbp) | 13.345 | 0.355 |
| 19 | Dmbt1 | deleted in malignant brain tumors 1 (Dmbt1) | 47.186 | 0.024 |
| 20 | Elovl3 | elongation of very long chain fatty acids | 0.271 | 2.786 |
| 21 | Fabp5 | fatty acid binding protein 5, epidermal (Fabp5) | 0.076 | 2.275 |
| 22 | Fam25c | family with sequence similarity 25, member C (Fam25c) | 0.168 | 9.567 |
| 23 | Gadd45g | growth arrest and DNA-damage-inducible 45 gamma (Gadd45g) | 0.267 | 3.741 |
| 24 | Gas6 | growth arrest specific 6 (Gas6) | 2.145 | 0.455 |
| 25 | Gm3219 | B-cell CLL/lymphoma 7C pseudogene (Gm3219) | 0.401 | 3.253 |
| 26 | Gstm2 | glutathione S-transferase, mu 2 (Gstm2) | 2.275 | 0.430 |
| 27 | Gstm3 | glutathione S-transferase, mu 3 (Gstm3) | 3.361 | 0.249 |
| 28 | H2-Eb1 | histocompatibility 2, class II antigen E beta (H2-Eb1) | 3.240 | 0.361 |
| 29 | Krt19 | keratin 19 (Krt19) | 4.125 | 0.287 |
| 30 | Lpin1 | lipin 1 (Lpin1) | 2.546 | 0.403 |
| 31 | Ly6a | lymphocyte antigen 6 complex, locus A (Ly6a) | 8.768 | 0.172 |
| 32 | Ly6c1 | lymphocyte antigen 6 complex, locus C1 (Ly6c1) | 8.054 | 0.166 |
| 33 | Ly6d | lymphocyte antigen 6 complex, locus D (Ly6d) | 5.567 | 0.463 |
| 34 | Mfsd2a | major facilitator superfamily domain containing 2A (Mfsd2a) | 4.328 | 0.352 |
| 35 | Moxd1 | monooxygenase, DBH-like 1 (Moxd1) | 0.020 | 31.917 |
| 36 | Mup21 | major urinary protein 21 (Mup21) | 0.456 | 2.180 |
| 37 | Myl9 | myosin, light polypeptide 9, regulatory (Myl9) | 2.474 | 0.405 |
| 38 | Rdh16 | retinol dehydrogenase 16 (Rdh16) | 3.376 | 0.283 |
| 39 | Rdh9 | retinol dehydrogenase 9 (Rdh9) | 3.309 | 0.350 |
| 40 | Saa3 | serum amyloid A 3 (Saa3) | 3.576 | 0.448 |
| 41 | Slc16a7 | solute carrier family 16 (monocarboxylic acid transporters), member 7 | 2.132 | 0.487 |
| 42 | Tceal8 | transcription elongation factor A (SII)-like 8 (Tceal8) | 2.259 | 0.420 |
| 43 | Upp2 | uridine phosphorylase 2 (Upp2) | 5.374 | 0.443 |
| S. No. | Term | Count | % | p-Value |
|---|---|---|---|---|
| 1. | Lipid metabolism | 21 | 14.5 | 9 × 10−13 |
| 2. | Cholesterol metabolism | 11 | 7.6 | 1.1 × 10−12 |
| 3. | Steroid metabolism | 11 | 7.6 | 2.4 × 10−11 |
| 4. | Cholesterol metabolic process | 12 | 8.3 | 3.3 × 10−11 |
| 5. | Fatty acid metabolic process | 11 | 7.6 | 1.6 × 10−7 |
| 6. | Lipid biosynthesis | 12 | 8.3 | 3.8 × 10−9 |
| 7. | Lipid metabolic process | 23 | 15.9 | 1.2 × 10−12 |
| 8. | Steroid metabolic process | 10 | 6.9 | 7.9 × 10−9 |
| 9. | Steroid hormone biosynthesis | 10 | 6.9 | 1.9 × 10−7 |
| 10. | Endoplasmic reticulum | 32 | 22.1 | 3.3 × 10−14 |
| 11. | Organelle membrane | 14 | 9.7 | 5.9 × 10−14 |
| 12. | Microsome | 15 | 10.3 | 6.5 × 10−14 |
| 13. | Heme binding | 17 | 11.7 | 6.6 × 10−14 |
| 14. | Endoplasmic reticulum membrane | 28 | 19.3 | 1.9 × 10−13 |
| 15. | Metal ion binding site:iron (heme axial ligand) | 14 | 9.7 | 8.9 × 10−13 |
| 16. | Iron ion binding | 17 | 11.7 | 1.3 × 10−12 |
| 17. | Secondary metabolites biosynthesis, transport, and catabolism | 17 | 11.7 | 2.5 × 10−12 |
| 18. | Sterol metabolism | 11 | 7.6 | 3.8 × 10−12 |
| 19. | Cytochrome p450, e-class, group i | 12 | 8.3 | 5.8 × 10−12 |
| 20. | Endoplasmic reticulum | 35 | 24.1 | 6 × 10−12 |
| 21. | Metabolic pathways | 40 | 27.6 | 6.5 × 10−12 |
| 22. | Iron | 19 | 13.1 | 1.3 × 10−11 |
| 23. | PPAR signaling pathway | 13 | 9 | 1.7 × 10−11 |
| 24. | Intracellular membrane-bounded organelle | 25 | 17.2 | 1.8 × 10−10 |
| 25. | Chemical carcinogenesis | 12 | 8.3 | 1.6 × 10−9 |
| 26. | Arachidonic acid metabolism | 10 | 6.9 | 2.3 × 10−7 |
| 27. | Disulfide bond | 40 | 27.6 | 5.1 × 10−6 |
| 28. | Metabolic process | 14 | 9.7 | 0.000025 |
| 29. | Biosynthesis of antibiotics | 11 | 7.6 | 0.000054 |
| 30. | Signal | 47 | 32.4 | 0.00013 |
| 31. | Extracellular exosome | 33 | 22.8 | 0.00068 |
| 32. | Glycoprotein | 39 | 26.9 | 0.00091 |
| 33. | Extracellular space | 22 | 15.2 | 0.001 |
| 34. | Secreted | 22 | 15.2 | 0.0012 |
| 35. | Disulfide bond | 31 | 21.4 | 0.0013 |
| 36. | Extracellular region | 24 | 16.6 | 0.0014 |
| 37. | Acetylation | 33 | 22.8 | 0.0017 |
| 38. | Catalytic activity | 11 | 7.6 | 0.0017 |
| 39. | Signal peptide | 35 | 24.1 | 0.0029 |
| 40. | Protein homodimerization activity | 13 | 9 | 0.0089 |
| 41. | Membrane | 59 | 40.7 | 0.029 |
| 42. | Metal binding | 30 | 20.7 | 0.032 |
| 43. | Lipoprotein | 10 | 6.9 | 0.047 |
| 44. | Hydrolase activity | 17 | 11.7 | 0.061 |
| S. No. | Term | Count | % | p-Value |
|---|---|---|---|---|
| 1. | Monooxygenase | 13 | 21.7 | 5.50 × 10−16 |
| 2. | Cytochrome p450, conserved site | 12 | 20 | 1.20 × 10−15 |
| 3. | Cytochrome p450 | 12 | 20 | 3.10 × 10−15 |
| 4. | Retinol metabolism | 12 | 20 | 1.90 × 10−14 |
| 5. | Oxidoreductase activity, acting on paired donors | 11 | 18.3 | 2.30 × 10−13 |
| 6. | Heme | 12 | 20 | 4.40 × 10−13 |
| 7. | Iron ion binding | 13 | 21.7 | 7.90 × 10−13 |
| 8. | Heme binding | 12 | 20 | 2.50 × 10−12 |
| 9. | Organelle membrane | 10 | 16.7 | 3.90 × 10−12 |
| 10. | Monooxygenase activity | 10 | 16.7 | 2.30 × 10−11 |
| 11. | Secondary metabolites biosynthesis, transport | 12 | 20 | 3.10 × 10−11 |
| 12. | Metal ion binding site:iron (heme axial ligand) | 10 | 16.7 | 3.50 × 10−11 |
| 13. | Microsome | 10 | 16.7 | 4.10 × 10−11 |
| 14. | Iron | 13 | 21.7 | 1.30 × 10−10 |
| 15. | Oxidoreductase | 15 | 25 | 4.30 × 10−10 |
| 16. | Endoplasmic reticulum | 17 | 28.3 | 1.80 × 10−9 |
| 17. | Endoplasmic reticulum membrane | 15 | 25 | 4.80 × 10−9 |
| 18. | Intracellular membrane-bounded organelle | 13 | 21.7 | 6.90 × 10−7 |
| 19. | Endoplasmic reticulum | 16 | 26.7 | 1.70 × 10−6 |
| 20. | Oxidation-reduction process | 12 | 20 | 2.20 × 10−6 |
| 21. | Metabolic pathways | 17 | 28.3 | 6.90 × 10−6 |
| 22. | Oxidoreductase activity | 11 | 18.3 | 8.10 × 10−6 |
| 23. | Metal binding | 17 | 28.3 | 7.90 × 10−3 |
| 24. | Disulfide bond | 16 | 26.7 | 8.80 × 10−3 |
| 25. | Glycoprotein | 18 | 30 | 1.10 × 10−2 |
| 26. | Signal | 19 | 31.7 | 2.60 × 10−2 |
| 27. | Disulfide bond | 13 | 21.7 | 5.10 × 10−2 |
| 28. | Membrane | 29 | 48.3 | 6.20 × 10−2 |
| Term | Counts | % | p-Value | |
|---|---|---|---|---|
| 1. | Retinol metabolism | 18 | 12.4 | 8.4 × 10−18 |
| 2. | Metabolic pathways | 40 | 27.6 | 6.5 × 10−12 |
| 3. | PPAR signaling pathway | 13 | 9 | 1.7 × 10−11 |
| 4. | Chemical carcinogenesis | 12 | 8.3 | 1.6 × 10−9 |
| 5. | Steroid hormone biosynthesis | 10 | 6.9 | 0.00000019 |
| 6. | Arachidonic acid metabolism | 10 | 6.9 | 0.00000023 |
| 7. | Fatty acid degradation | 7 | 4.8 | 0.0000089 |
| 8. | Linoleic acid metabolism | 7 | 4.8 | 0.00001 |
| 9. | Inflammatory mediator regulation of TRP channels | 9 | 6.2 | 0.000037 |
| 10. | Biosynthesis of antibiotics | 11 | 7.6 | 0.000054 |
| 11. | Terpenoid backbone biosynthesis | 4 | 2.8 | 0.0015 |
| 12. | Propanoate metabolism | 4 | 2.8 | 0.0025 |
| 13. | Serotonergic synapse | 6 | 4.1 | 0.011 |
| 14. | Fatty acid metabolism | 4 | 2.8 | 0.015 |
| 15. | Steroid biosynthesis | 3 | 2.1 | 0.016 |
| 16. | Valine, leucine, and isoleucine degradation | 4 | 2.8 | 0.018 |
| 17. | Asthma | 3 | 2.1 | 0.024 |
| 18. | Metabolism of xenobiotics by cytochrome P450 | 4 | 2.8 | 0.027 |
| 19. | Circadian rhythm | 3 | 2.1 | 0.039 |
| 20. | Vascular smooth muscle contraction | 5 | 3.4 | 0.04 |
| 21. | beta-alanine metabolism | 3 | 2.1 | 0.044 |
| 22. | Antigen processing and presentation | 4 | 2.8 | 0.05 |
| 23. | Intestinal immune network for IgA production | 3 | 2.1 | 0.067 |
| 24. | Tryptophan metabolism | 3 | 2.1 | 0.082 |
| 25. | Staphylococcus aureus infection | 3 | 2.1 | 0.091 |
| 26. | Drug metabolism—other enzymes | 3 | 2.1 | 0.094 |
| 27. | Graft-versus-host disease | 3 | 2.1 | 0.097 |
| Term | Count | % | p-Value | |
|---|---|---|---|---|
| 1. | Retinol metabolism | 12 | 20 | 1.90 × 10−14 |
| 2. | PPAR signaling pathway | 8 | 13.3 | 2.20 × 10−8 |
| 3. | Chemical carcinogenesis | 8 | 13.3 | 5.80 × 10−8 |
| 4. | Arachidonic acid metabolism | 7 | 11.7 | 1.20 × 10−6 |
| 5. | Metabolic pathways | 17 | 28.3 | 6.90 × 10−6 |
| 6. | Steroid hormone biosynthesis | 6 | 10 | 2.20 × 10−5 |
| 7. | Fatty acid degradation | 4 | 6.7 | 9.60 × 10−4 |
| 8. | Inflammatory mediator regulation of TRP channels | 5 | 8.3 | 1.50 × 10−3 |
| 9. | Vascular smooth muscle contraction | 5 | 8.3 | 1.60 × 10−3 |
| 10. | Linoleic acid metabolism | 3 | 5 | 1.70 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soundharrajan, I.; Karnan, M.; Jung, J.-S.; Lee, K.-D.; Lee, J.-C.; Ramesh, T.; Kim, D.; Choi, K.-C. A Transcriptomic Response to Lactiplantibacillus plantarum-KCC48 against High-Fat Diet-Induced Fatty Liver Diseases in Mice. Int. J. Mol. Sci. 2022, 23, 6750. https://doi.org/10.3390/ijms23126750
Soundharrajan I, Karnan M, Jung J-S, Lee K-D, Lee J-C, Ramesh T, Kim D, Choi K-C. A Transcriptomic Response to Lactiplantibacillus plantarum-KCC48 against High-Fat Diet-Induced Fatty Liver Diseases in Mice. International Journal of Molecular Sciences. 2022; 23(12):6750. https://doi.org/10.3390/ijms23126750
Chicago/Turabian StyleSoundharrajan, Ilavenil, Muthusamy Karnan, Jeong-Sung Jung, Kyung-Dong Lee, Jeong-Chae Lee, Thiyagarajan Ramesh, Dahye Kim, and Ki-Choon Choi. 2022. "A Transcriptomic Response to Lactiplantibacillus plantarum-KCC48 against High-Fat Diet-Induced Fatty Liver Diseases in Mice" International Journal of Molecular Sciences 23, no. 12: 6750. https://doi.org/10.3390/ijms23126750