
Immunogenicity of MultiTEP-Platform-Based Recombinant Protein Vaccine, PV-1950R, Targeting Three B-Cell Antigenic Determinants of Pathological α-Synuclein

 ,
,
Abstract
:1. Introduction
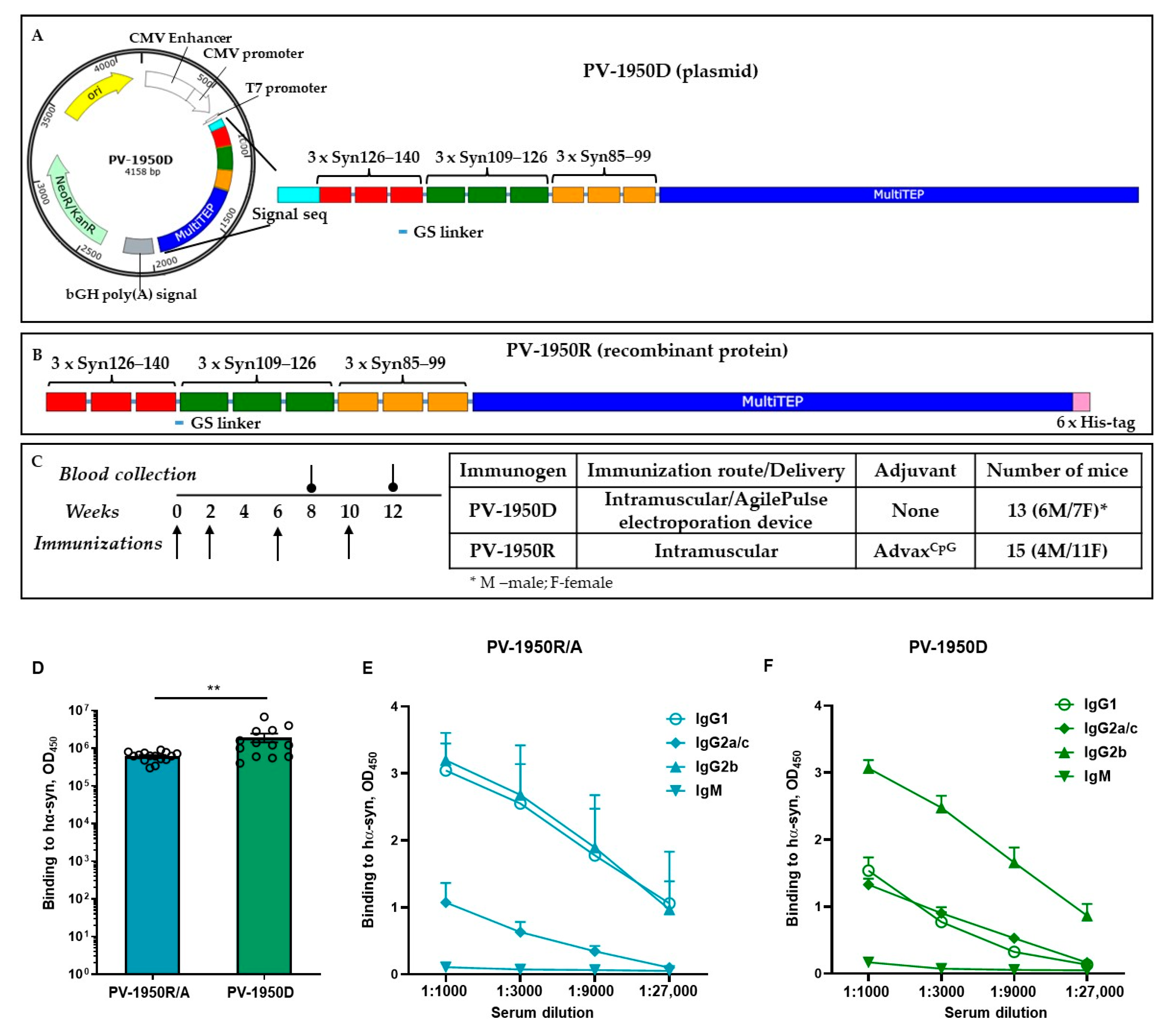
2. Results
2.1. Immunogenicity of Recombinant Protein Vaccine, PV-1950R/A, in Young and Old D-Line Mice
2.2. Comparison of Humoral Immune Responses in D-Line Mice Vaccinated with PV-1950D and PV-1950R/A
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Vaccines
4.3. Immunization of Mice
4.4. Detection of Antibody Titers and Isotypes
4.5. Measuring Avidity of Antibodies
4.6. Detection of hα-Syn in Human Brain Tissues from AD/DLB Cases
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luk, K.C.; Kehm, V.M.; Zhang, B.; O’Brien, P.; Trojanowski, J.Q.; Lee, V.M. Intracerebral inoculation of pathological alpha-synuclein initiates a rapidly progressive neurodegenerative alpha-synucleinopathy in mice. J. Exp. Med. 2012, 209, 975–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castonguay, A.M.; Gravel, C.; Levesque, M. Treating Parkinson’s Disease with Antibodies: Previous Studies and Future Directions. J. Parkinson’s Dis. 2021, 11, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Chung, C.H.; Iba, M.; Zhang, B.; Trojanowski, J.Q.; Luk, K.C.; Lee, V.M. Alpha-synuclein immunotherapy blocks uptake and templated propagation of misfolded alpha-synuclein and neurodegeneration. Cell. Rep. 2014, 7, 2054–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutajangout, A.; Ingadottir, J.; Davies, P.; Sigurdsson, E.M. Passive immunization targeting pathological phospho-tau protein in a mouse model reduces functional decline and clears tau aggregates from the brain. J. Neurochem. 2011, 118, 658–667. [Google Scholar] [CrossRef] [Green Version]
- Yanamandra, K.; Kfoury, N.; Jiang, H.; Mahan, T.E.; Ma, S.; Maloney, S.E.; Wozniak, D.F.; Diamond, M.I.; Holtzman, D.M. Anti-tau antibodies that block tau aggregate seeding in vitro markedly decrease pathology and improve cognition in vivo. Neuron 2013, 80, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Masliah, E.; Rockenstein, E.; Mante, M.; Crews, L.; Spencer, B.; Adame, A.; Patrick, C.; Trejo, M.; Ubhi, K.; Rohn, T.T.; et al. Passive immunization reduces behavioral and neuropathological deficits in an alpha-synuclein transgenic model of Lewy body disease. PLoS ONE 2011, 6, e19338. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.J.; Lee, H.J.; Rockenstein, E.; Ho, D.H.; Park, E.B.; Yang, N.Y.; Desplats, P.; Masliah, E.; Lee, S.J. Antibody-aided clearance of extracellular alpha-synuclein prevents cell-to-cell aggregate transmission. J. Neurosci 2012, 32, 13454–13469. [Google Scholar] [CrossRef]
- Valastyan, J.S.; Lindquist, S. Mechanisms of protein-folding diseases at a glance. Dis. Model. Mech 2014, 7, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Gros-Louis, F.; Soucy, G.; Lariviere, R.; Julien, J.P. Intracerebroventricular infusion of monoclonal antibody or its derived Fab fragment against misfolded forms of SOD1 mutant delays mortality in a mouse model of ALS. J. Neurochem. 2010, 113, 1188–1199. [Google Scholar] [CrossRef]
- Wolfgang, W.J.; Miller, T.W.; Webster, J.M.; Huston, J.S.; Thompson, L.M.; Marsh, J.L.; Messer, A. Suppression of Huntington’s disease pathology in Drosophila by human single-chain Fv antibodies. Proc. Natl. Acad. Sci. USA 2005, 102, 11563–11568. [Google Scholar] [CrossRef] [Green Version]
- Schenk, D.B.; Koller, M.; Ness, D.K.; Griffith, S.G.; Grundman, M.; Zago, W.; Soto, J.; Atiee, G.; Ostrowitzki, S.; Kinney, G.G. First-in-human assessment of PRX002, an anti-alpha-synuclein monoclonal antibody, in healthy volunteers. Mov. Disord. 2017, 32, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.S.; Ooi, Y.Y.; Chye, S.M.; Ling, A.P.K.; Koh, R.Y. Immunotherapies for Parkinson’s Disease: Progression of Clinical Development. CNS Neurol. Disord. Drug Targets 2021, 20, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Brys, M.; Fanning, L.; Hung, S.; Ellenbogen, A.; Penner, N.; Yang, M.; Welch, M.; Koenig, E.; David, E.; Fox, T.; et al. Randomized phase I clinical trial of anti-alpha-synuclein antibody BIIB054. Mov. Disord. 2019, 34, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Haeberlein, S.B.; von Hehn, C.; Tian, Y.; Chalkias, S.; Muralidharan, K.K.; Chen, T.; Wu, S.; Li, J.; Skordos, L.; Nisenbaum, L.; et al. EMERGE and ENGAGE Topline Results: Two Phase 3 Studies to Evaluate Aducanumab in Patients with Early Alzheimer’s Disease. In Proceedings of the 12th Clinical Trials on Alzheimer’s Disease (CTAD), San Diego, CA, USA, 4–7 December 2019. [Google Scholar]
- Avgerinos, K.I.; Ferrucci, L.; Kapogiannis, D. Effects of monoclonal antibodies against amyloid-beta on clinical and biomarker outcomes and adverse event risks: A systematic review and meta-analysis of phase III RCTs in Alzheimer’s disease. Ageing Res. Rev. 2021, 68, 101339. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Toth, B.; Wildsmith, K.; Bohorquez, S.S.; Brunstein, B.; Madsen, A.; Dolton, M.; Ramakrishnan, V.; Abramzon, D.; Teng, E. Phase 2 Trial of Semorinemab in Mild-to-Moderate Alzheimer’s Disease (Lauriet): Topline Results. In Proceedings of the Clinical Trials on Alzheimer’s Disease, Boston, MA, USA, 9–12 November 2021. [Google Scholar]
- Vaz, M.; Silvestre, S. Alzheimer’s disease: Recent treatment strategies. Eur. J. Pharmacol. 2020, 887, 173554. [Google Scholar] [CrossRef]
- Penn, D.J. Major Histocompatibility Complex (MHC). Encycl. Life Sci. 2002, 1–7. [Google Scholar]
- Tumer, G.; Simpson, B.; Roberts, T.K. Genetics, Human Major Histocompatibility Complex (MHC). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Davtyan, H.; Zagorski, K.; Petrushina, I.; Kazarian, K.; Goldberg, N.R.S.; Petrosyan, J.; Blurton-Jones, M.; Masliah, E.; Cribbs, D.H.; Agadjanyan, M.G.; et al. MultiTEP platform-based DNA vaccines for alpha-synucleinopathies: Preclinical evaluation of immunogenicity and therapeutic potency. Neurobiol. Aging 2017, 59, 156–170. [Google Scholar] [CrossRef]
- Kim, C.; Hovakimyan, A.; Zagorski, K.; Antonyan, T.; Petrushina, I.; Davtyan, H.; Chailyan, G.; Hasselmann, J.; Iba, M.; Adame, A.; et al. efficacy and immunogenicity of MultiTEP-based DNA vaccines targeting human alpha-synuclein: Prelude for IND enabling studies. NPJ Vaccines 2022, 7, 1. [Google Scholar] [CrossRef]
- Eusebio, D.; Neves, A.R.; Costa, D.; Biswas, S.; Alves, G.; Cui, Z.; Sousa, A. Methods to improve the immunogenicity of plasmid DNA vaccines. Drug Discov Today 2021, 26, 2575–2592. [Google Scholar] [CrossRef]
- Momin, T.; Kansagra, K.; Patel, H.; Sharma, S.; Sharma, B.; Patel, J.; Mittal, R.; Sanmukhani, J.; Maithal, K.; Dey, A.; et al. Safety and Immunogenicity of a DNA SARS-CoV-2 vaccine (ZyCoV-D): Results of an open-label, non-randomized phase I part of phase I/II clinical study by intradermal route in healthy subjects in India. EClinicalMedicine 2021, 38, 101020. [Google Scholar] [CrossRef]
- Shafaati, M.; Saidijam, M.; Soleimani, M.; Hazrati, F.; Mirzaei, R.; Amirheidari, B.; Tanzadehpanah, H.; Karampoor, S.; Kazemi, S.; Yavari, B.; et al. A brief review on DNA vaccines in the era of COVID-19. Future Virol. 2021, 17, 49–66. [Google Scholar] [CrossRef]
- Grodeland, G.; Fossum, E.; Bogen, B. Polarizing T and B Cell Responses by APC-Targeted Subunit Vaccines. Front. Immunol. 2015, 6, 367. [Google Scholar] [CrossRef] [PubMed]
- Finkelman, F.D.; Holmes, J.; Katona, I.M.; Urban, J.F.; Beckmann, M.P.; Park, L.S.; Schooley, K.A.; Coffman, R.L.; Mossmann, T.R.; Paul, W.E. Lymphokine control of in vivo immunoglobulin isotype selection. Ann. Rev. Immunol. 1990, 8, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Coffman, R.L. Heterogeneity of cytokine secretion patterns and functions of helper T cells. Adv. Immunol. 1989, 46, 111–147. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.L.; Bossie, A.; Sanders, V.M.; Fernandez-Botran, R.; Coffman, R.L.; Mosmann, T.R.; Vitetta, E.S. Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature 1988, 334, 255–258. [Google Scholar] [CrossRef]
- Snapper, C.M.; Paul, W.E. Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science 1987, 236, 944–947. [Google Scholar] [CrossRef]
- Liu, M.A. A Comparison of Plasmid DNA and mRNA as Vaccine Technologies. Vaccines 2019, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Hogan, M.J.; Weissman, D. Recent advances in mRNA vaccine technology. Curr. Opin. Immunol. 2020, 65, 14–20. [Google Scholar] [CrossRef]
- Plotkin, S.; Orenstein, W.; Offit, P.; Edwards, K.M. Plotkin’s Vaccines, 7th ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Ghochikyan, A.; Petrushina, I.; Davtyan, H.; Hovakimyan, A.; Saing, T.; Davtyan, A.; Cribbs, D.H.; Agadjanyan, M.G. Immunogenicity of epitope vaccines targeting different B cell antigenic determinants of human alpha-synuclein: Feasibility study. Neurosci. Lett. 2014, 560, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Davtyan, H.; Hovakimyan, A.; Kiani Shabestari, S.; Antonyan, T.; Coburn, M.A.; Zagorski, K.; Chailyan, G.; Petrushina, I.; Svystun, O.; Danhash, E.; et al. Testing a MultiTEP-based combination vaccine to reduce Abeta and tau pathology in Tau22/5xFAD bigenic mice. Alzheimers Res. Ther. 2019, 11, 107. [Google Scholar] [CrossRef]
- Petrushina, I.; Hovakimyan, A.; Harahap-Carrillo, I.S.; Davtyan, H.; Antonyan, T.; Chailyan, G.; Kazarian, K.; Antonenko, M.; Jullienne, A.; Hamer, M.M.; et al. Characterization and preclinical evaluation of the cGMP grade DNA based vaccine, AV-1959D to enter the first-in-human clinical trials. Neurobiol. Dis. 2020, 139, 104823. [Google Scholar] [CrossRef] [PubMed]
- Hovakimyan, A.; Antonyan, T.; Shabestari, S.K.; Svystun, O.; Chailyan, G.; Coburn, M.A.; Carlen-Jones, W.; Petrushina, I.; Chadarevian, J.P.; Zagorski, K.; et al. A MultiTEP platform-based epitope vaccine targeting the phosphatase activating domain (PAD) of tau: Therapeutic efficacy in PS19 mice. Sci. Rep. 2019, 9, 15455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davtyan, H.; Chen, W.W.; Zagorski, K.; Davis, J.; Petrushina, I.; Kazarian, K.; Cribbs, D.H.; Agadjanyan, M.G.; Blurton, -J.M.; Ghochikyan, A. MultiTEP platform-based DNA epitope vaccine targeting N-terminus of tau induces strong immune responses and reduces tau pathology in THY-Tau22 mice. Vaccine 2017, 35, 2015–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeer, P.L.; McGeer, E.G. The alpha-synuclein burden hypothesis of Parkinson disease and its relationship to Alzheimer disease. Exp. Neurol. 2008, 212, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Twohig, D.; Nielsen, H.M. alpha-synuclein in the pathophysiology of Alzheimer’s disease. Mol. Neurodegener 2019, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Ghochikyan, A.; Davtyan, H.; Petrushina, I.; Hovakimyan, A.; Movsesyan, N.; Davtyan, A.; Kiyatkin, A.; Cribbs, D.H.; Agadjanyan, M.G. Refinement of a DNA based Alzheimer’s disease epitope vaccine in rabbits. Hum. Vaccin Immunother. 2013, 9, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Davtyan, H.; Ghochikyan, A.; Petrushina, I.; Hovakimyan, A.; Davtyan, A.; Cribbs, D.H.; Agadjanyan, M.G. The MultiTEP platform-based Alzheimer’s disease epitope vaccine activates a broad repertoire of T helper cells in non-human primates. Alzheimers Dement. 2014, 10, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.F.; Davtyan, H.; Petrushina, I.; Hovakimyan, A.; Davtyan, A.; Hannaman, D.; Cribbs, D.H.; Agadjanyan, M.G.; Ghochikyan, A. Epitope-based DNA vaccine for Alzheimer’s disease: Translational study in macaques. Alzheimers Dement. 2014, 10, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Davtyan, H.; Zagorski, K.; Rajapaksha, H.; Hovakimyan, A.; Davtyan, A.; Petrushina, I.; Kazarian, K.; Cribbs, D.H.; Petrovsky, N.; Agadjanyan, M.G.; et al. Alzheimer’s disease Advax(CpG)-adjuvanted MultiTEP-based dual and single vaccines induce high-titer antibodies against various forms of tau and Abeta pathological molecules. Sci. Rep. 2016, 6, 28912. [Google Scholar] [CrossRef] [Green Version]
- Petrushina, I.; Davtyan, H.; Hovakimyan, A.; Davtyan, A.; Passos, G.F.; Cribbs, D.H.; Ghochikyan, A.; Agadjanyan, M.G. Comparison of Efficacy of Preventive and Therapeutic Vaccines Targeting the N Terminus of beta-Amyloid in an Animal Model of Alzheimer’s Disease. Mol. Ther. 2017, 25, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Goronzy, J.J.; Fang, F.; Cavanagh, M.M.; Qi, Q.; Weyand, C.M. Naive T cell maintenance and function in human aging. J. Immunol 2015, 194, 4073–4080. [Google Scholar] [CrossRef] [PubMed]
- Derhovanessian, E.; Solana, R.; Larbi, A.; Pawelec, G. Immunity, ageing and cancer. Immun. Ageing 2008, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davtyan, H.; Mkrtichyan, M.; Movsesyan, N.; Petrushina, I.; Mamikonyan, G.; Cribbs, D.H.; Agadjanyan, M.G.; Ghochikyan, A. DNA prime-protein boost increased the titer, avidity and persistence of anti-Abeta antibodies in wild-type mice. Gene Ther. 2010, 17, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richmond, J.F.; Lu, S.; Santoro, J.C.; Weng, J.; Hu, S.L.; Montefiori, D.C.; Robinson, H.L. Studies of the neutralizing activity and avidity of anti-human immunodeficiency virus type 1 Env antibody elicited by DNA priming and protein boosting. J. Virol. 1998, 72, 9092–9100. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Age of D-Line Mice at the Start of Immunization # | Endpoint Titers (±SEM) of Antibodies Binding to Three B cell Epitopes of α-Syn and Full-Length α-Syn | |||
|---|---|---|---|---|
| aa 85-99 | aa 109-126 | aa 126-140 | α-Syn | |
| 2-4 mo old | 1:16236 ±6 860 | 1:61000 ± 11685 | 1:179600 ± 43634 | 1:616667 ± 43124 |
| 12-14 mo old | 1:11865 ± 5109 | 1:10453 ± 4196 | 1:12082 ± 7686 | 1:231000 ± 25438 |
| p value | 0.6312 | 0.0016 | 0.0035 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zagorski, K.; Chailyan, G.; Hovakimyan, A.; Antonyan, T.; Kiani Shabestari, S.; Petrushina, I.; Davtyan, H.; Cribbs, D.H.; Blurton-Jones, M.; Masliah, E.; et al. Immunogenicity of MultiTEP-Platform-Based Recombinant Protein Vaccine, PV-1950R, Targeting Three B-Cell Antigenic Determinants of Pathological α-Synuclein. Int. J. Mol. Sci. 2022, 23, 6080. https://doi.org/10.3390/ijms23116080
Zagorski K, Chailyan G, Hovakimyan A, Antonyan T, Kiani Shabestari S, Petrushina I, Davtyan H, Cribbs DH, Blurton-Jones M, Masliah E, et al. Immunogenicity of MultiTEP-Platform-Based Recombinant Protein Vaccine, PV-1950R, Targeting Three B-Cell Antigenic Determinants of Pathological α-Synuclein. International Journal of Molecular Sciences. 2022; 23(11):6080. https://doi.org/10.3390/ijms23116080
Chicago/Turabian StyleZagorski, Karen, Gor Chailyan, Armine Hovakimyan, Tatevik Antonyan, Sepideh Kiani Shabestari, Irina Petrushina, Hayk Davtyan, David H. Cribbs, Mathew Blurton-Jones, Eliezer Masliah, and et al. 2022. "Immunogenicity of MultiTEP-Platform-Based Recombinant Protein Vaccine, PV-1950R, Targeting Three B-Cell Antigenic Determinants of Pathological α-Synuclein" International Journal of Molecular Sciences 23, no. 11: 6080. https://doi.org/10.3390/ijms23116080