
Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics

Abstract
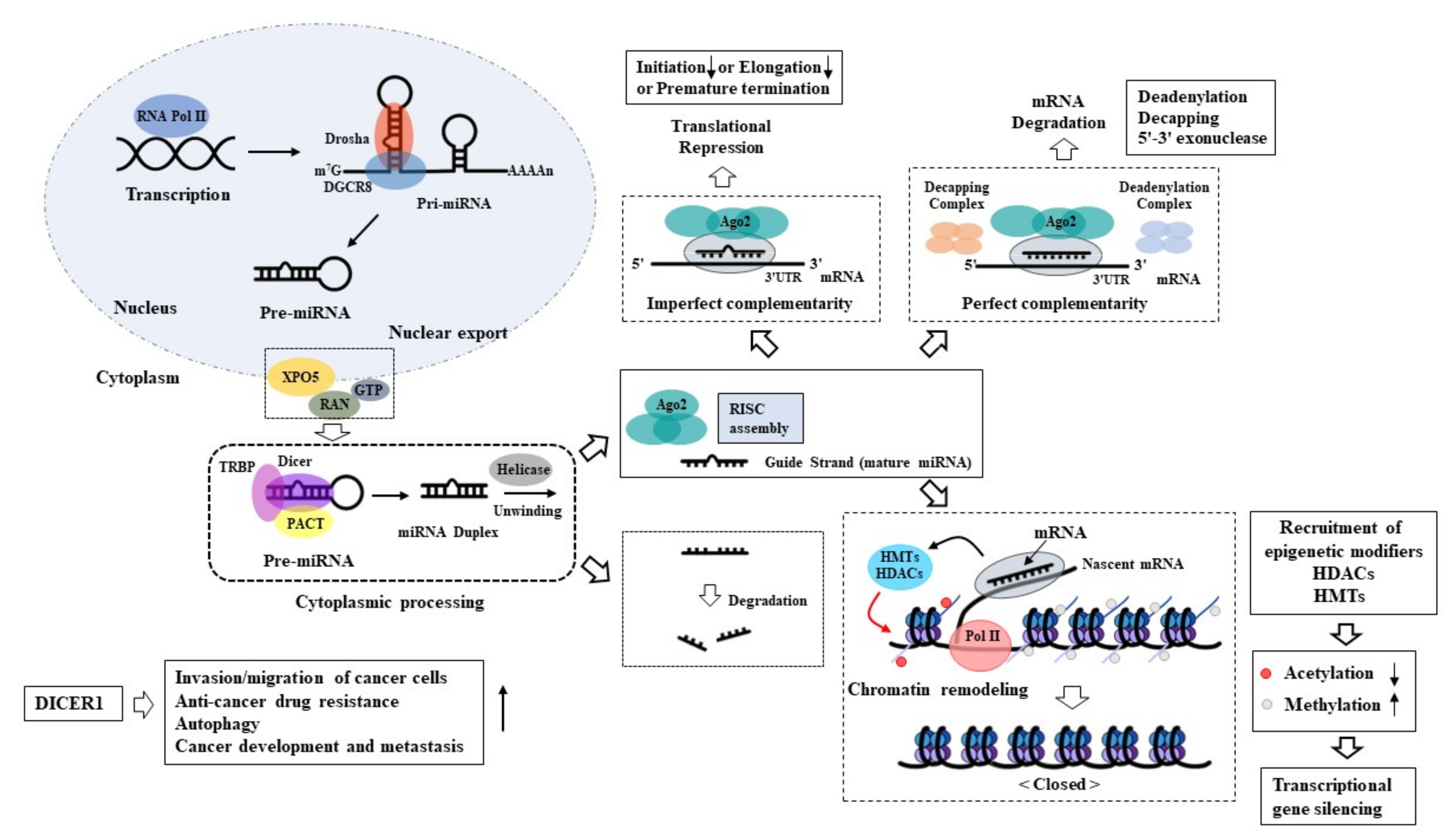
:1. Biogenesis of MicroRNAs
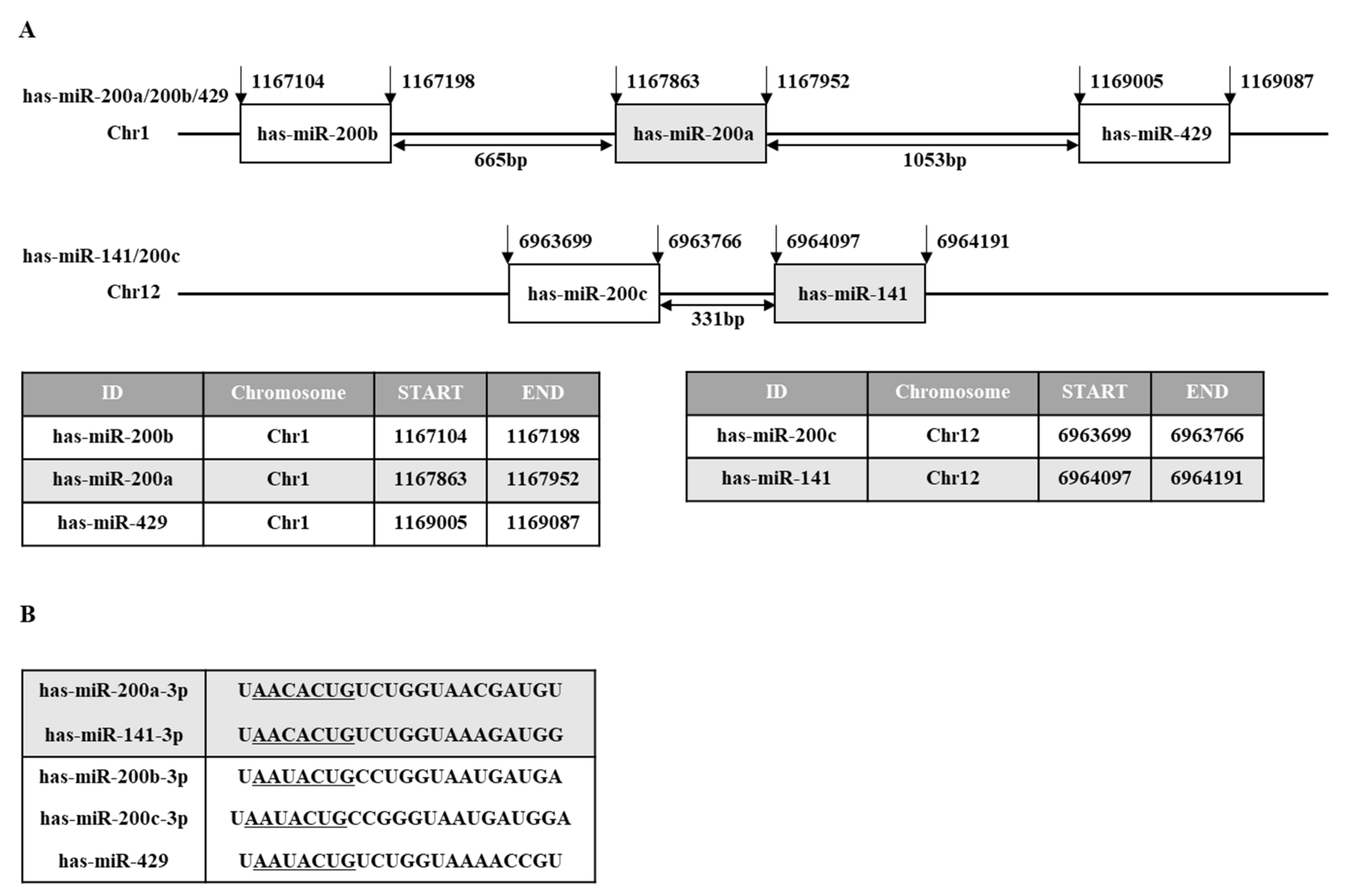
2. miR-200 Family as a Diagnostic and Prognostic Marker
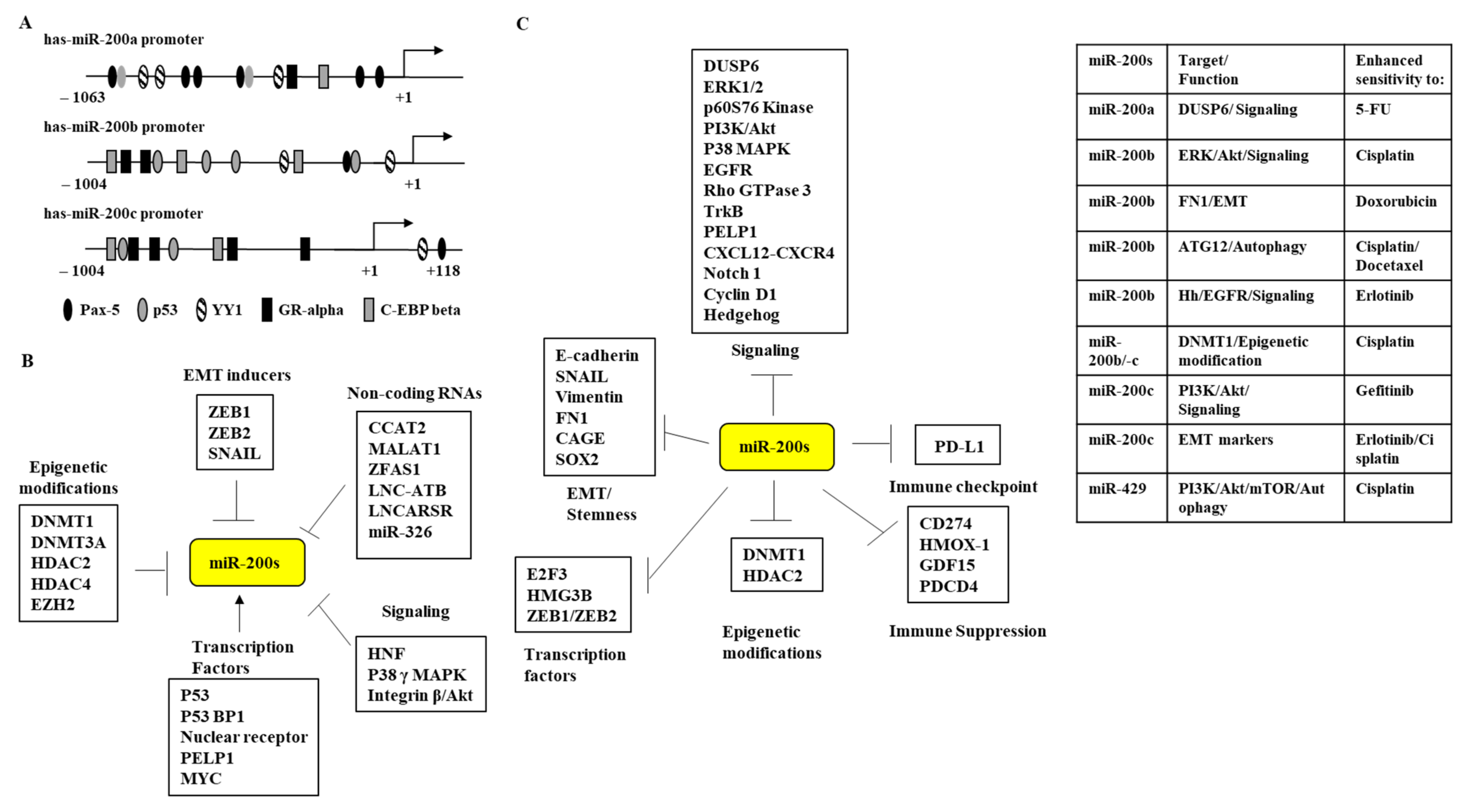
3. Regulation of miR-200 Family Expression
4. miR-200 Family and Targets of the miR-200 Family
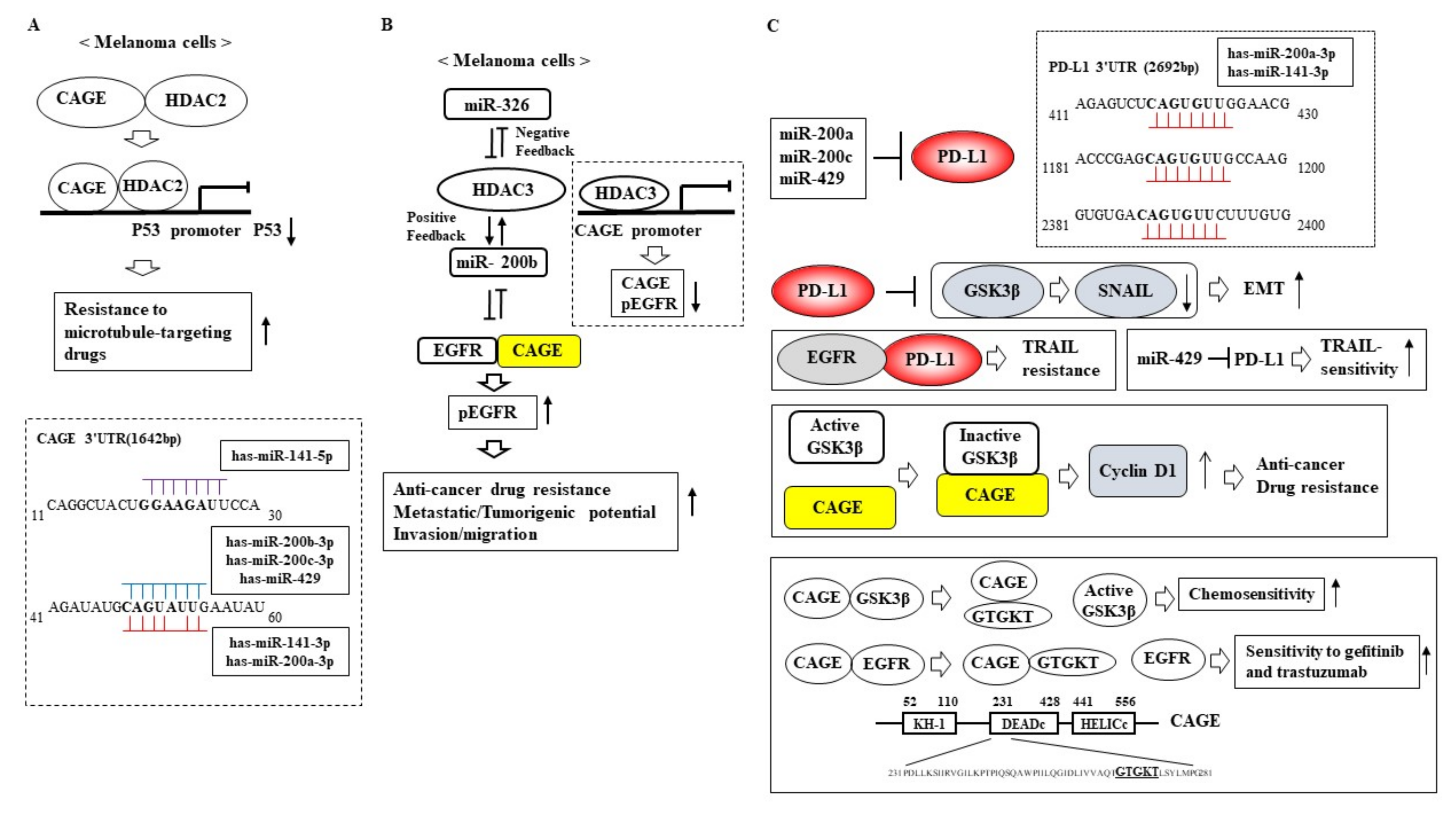
5. Role of the miR-200 Family in Anti-Cancer Drug Resistance
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Liu, W.; Ma, C.; Xu, H.; Wang, L.; Xu, W.; Zhang, H.; Wang, Z.; Li, J.; Zhang, J.; Liu, X.; et al. miR-184-5p inhibits cell proliferation, invasion and predicts prognosis of clear cell renal cell carcinoma by targeting NUS1 dehydrodolichyl diphosphate synthase subunit: Results from large-scale comprehensive identification and validation. J. Cancer 2022, 13, 1398–1409. [Google Scholar] [CrossRef]
- Chen, S.; Tu, Y.; Yuan, H.; Shi, Z.; Guo, Y.; Gong, W.; Tu, S. Regulatory functions of miR200b3p in tumor development (Review). Oncol. Rep. 2022, 47, 96. [Google Scholar] [CrossRef]
- Luo, H.; Lv, W.; Zhang, H.; Lin, C.; Li, F.; Zheng, F.; Zhong, B. miR-204-5p inhibits cell proliferation and induces cell apoptosis in esophageal squamous cell carcinoma by regulating Nestin. Int. J. Med. Sci. 2022, 19, 472–483. [Google Scholar] [CrossRef]
- Zhang, F.; Cheng, N.; Du, J.; Zhang, H.; Zhang, C. MicroRNA-200b-3p promotes endothelial cell apoptosis by targeting HDAC4 in atherosclerosis. BMC Cardiovasc. Disord. 2021, 21, 172. [Google Scholar] [CrossRef]
- Simpson, K.; Conquer-van Heumen, G.; Watson, K.L.; Roth, M.; Martin, C.J.; Moorehead, R.A. Re-expression of miR-200s in claudin-low mammary tumor cells alters cell shape and reduces proliferation and invasion potentially through modulating other miRNAs and SUZ12 regulated genes. Cancer Cell Int. 2021, 21, 89. [Google Scholar] [CrossRef]
- Tang, H.; Wang, X.; Zhang, M.; Yan, Y.; Huang, S.; Ji, J.; Xu, J.; Zhang, Y.; Cai, Y.; Yang, B.; et al. MicroRNA-200b/c-3p regulate epithelial plasticity and inhibit cutaneous wound healing by modulating TGF-beta-mediated RAC1 signaling. Cell Death Dis. 2020, 11, 931. [Google Scholar] [CrossRef]
- Quirico, L.; Orso, F.; Cucinelli, S.; Paradzik, M.; Natalini, D.; Centonze, G.; Dalmasso, A.; La Vecchia, S.; Coco, M.; Audrito, V.; et al. miRNA-guided reprogramming of glucose and glutamine metabolism and its impact on cell adhesion/migration during solid tumor progression. Cell Mol. Life Sci. 2022, 79, 216. [Google Scholar] [CrossRef]
- Lv, Q.; Xia, Q.; Li, A.; Wang, Z. circRNA_101277 Influences Cisplatin Resistance of Colorectal Cancer Cells by Modulating the miR-370/IL-6 Axis. Genet. Res. 2022, 2022, 4237327. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, F.; Guan, Y.; Meng, F.; Zhao, Z.; Su, Q.; Bao, W.; Wang, X.; Zhao, J.; Huo, Z.; et al. The Biogenesis of miRNAs and Their Role in the Development of Amyotrophic Lateral Sclerosis. Cells 2022, 11, 572. [Google Scholar] [CrossRef]
- Tang, L. Recapitulating miRNA biogenesis in cells. Nat. Methods 2022, 19, 35. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Krill, K.T.; Gurdziel, K.; Heaton, J.H.; Simon, D.P.; Hammer, G.D. Dicer deficiency reveals microRNAs predicted to control gene expression in the developing adrenal cortex. Mol. Endocrinol. 2013, 27, 754–768. [Google Scholar] [CrossRef]
- Li, B.; Lu, W.; Qu, J.; Zhang, Y.; Wan, X. DICER1 regulates endometrial carcinoma invasion via histone acetylation and methylation. J. Cancer 2017, 8, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Chen, L.; Song, W.; Peng, B.; Zhu, J.; Fang, L. DICER activates autophagy and promotes cisplatin resistance in non-small cell lung cancer by binding with let-7i-5p. Acta Histochem. 2021, 123, 151788. [Google Scholar] [CrossRef]
- Luan, N.; Mu, Y.; Mu, J.; Chen, Y.; Ye, X.; Zhou, Q.; Xu, M.; Deng, Q.; Hu, Y.; Tang, Z.; et al. Dicer1 Promotes Colon Cancer Cell Invasion and Migration Through Modulation of tRF-20-MEJB5Y13 Expression Under Hypoxia. Front. Genet. 2021, 12, 638244. [Google Scholar] [CrossRef]
- Iliou, M.S.; da Silva-Diz, V.; Carmona, F.J.; Ramalho-Carvalho, J.; Heyn, H.; Villanueva, A.; Munoz, P.; Esteller, M. Impaired DICER1 function promotes stemness and metastasis in colon cancer. Oncogene 2014, 33, 4003–4015. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, Y.; Ge, F. MiR-186 Suppressed Growth, Migration, and Invasion of Lung Adenocarcinoma Cells via Targeting Dicer1. J. Oncol. 2021, 2021, 6217469. [Google Scholar] [CrossRef]
- Li, Y.; Guan, B.; Liu, J.; Zhang, Z.; He, S.; Zhan, Y.; Su, B.; Han, H.; Zhang, X.; Wang, B.; et al. MicroRNA-200b is downregulated and suppresses metastasis by targeting LAMA4 in renal cell carcinoma. EBioMedicine 2019, 44, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Gong, S.; Yan, T.; Yang, Y. MicroRNA-200b expression level is negatively associated with pathological grading in human gliomas. Cancer Manag. Res. 2018, 10, 2825–2834. [Google Scholar] [CrossRef]
- Saleeb, R.; Kim, S.S.; Ding, Q.; Scorilas, A.; Lin, S.; Khella, H.W.; Boulos, C.; Ibrahim, G.; Yousef, G.M. The miR-200 family as prognostic markers in clear cell renal cell carcinoma. Urol. Oncol. 2019, 37, 955–963. [Google Scholar] [CrossRef]
- Xu, T.; Xie, F.; Xu, D.; Xu, W.; Ge, X.; Lv, S.; Li, S. MiR-200b Suppresses Gastric Cancer Cell Migration and Invasion by Inhibiting NRG1 through ERBB2/ERBB3 Signaling. J. Oncol. 2021, 2021, 4470778. [Google Scholar] [CrossRef]
- Wang, L.K.; Xie, X.N.; Song, X.H.; Su, T.; Chang, X.L.; Xu, M.; Liang, B.; Huang, D.Y. Upregulation of miR-200b Inhibits Hepatocellular Carcinoma Cell Proliferation and Migration by Targeting HMGB3 Protein. Technol. Cancer Res. Treat. 2018, 17, 1533033818806475. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Zheng, J.; Xiang, P.; Liu, C.; Fan, Y. Prognostic value of the miR-200 family in bladder cancer: A systematic review and meta-analysis. Medicine 2020, 99, e22891. [Google Scholar] [CrossRef]
- Grenda, A.; Krawczyk, P.; Blach, J.; Chmielewska, I.; Kubiatowski, T.; Kieszko, S.; Wojas-Krawczyk, K.; Kucharczyk, T.; Jarosz, B.; Pasnik, I.; et al. Tissue MicroRNA Expression as a Predictor of Response to Immunotherapy in NSCLC Patients. Front. Oncol. 2020, 10, 563613. [Google Scholar] [CrossRef]
- Gharib, A.F.; Eed, E.M.; Khalifa, A.S.; Raafat, N.; Shehab-Eldeen, S.; Alwakeel, H.R.; Darwiesh, E.; Essa, A. Value of Serum miRNA-96-5p and miRNA-99a-5p as Diagnostic Biomarkers for Hepatocellular Carcinoma. Int. J. Gen. Med. 2022, 15, 2427–2436. [Google Scholar] [CrossRef]
- Santos, A.; Cristobal, I.; Rubio, J.; Carames, C.; Luque, M.; Sanz-Alvarez, M.; Morales-Gallego, M.; Madoz-Gurpide, J.; Rojo, F.; Garcia-Foncillas, J. MicroRNA-199b Deregulation Shows Oncogenic Properties and Promising Clinical Value as Circulating Marker in Locally Advanced Rectal Cancer Patients. Int J. Mol. Sci. 2022, 23, 2203. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, J.; Chen, Q.; Cheng, J.; Chen, X.; Mao, Y.; Chen, W.; Liu, C.; Wu, H.; Lv, Y.; et al. MicroRNA-200a and microRNA-141 have a synergetic effect on the suppression of epithelial-mesenchymal transition in liver cancer by targeting STAT4. Oncol. Lett. 2021, 21, 137. [Google Scholar] [CrossRef]
- Zou, J.G.; Ma, L.F.; Li, X.; Xu, F.L.; Fei, X.Z.; Liu, Q.; Bai, Q.L.; Dong, Y.L. Circulating microRNA array (miR-182, 200b and 205) for the early diagnosis and poor prognosis predictor of non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1108–1115. [Google Scholar]
- Papadaki, C.; Stoupis, G.; Tsalikis, L.; Monastirioti, A.; Papadaki, M.; Maliotis, N.; Stratigos, M.; Mastrostamatis, G.; Mavroudis, D.; Agelaki, S. Circulating miRNAs as a marker of metastatic disease and prognostic factor in metastatic breast cancer. Oncotarget 2019, 10, 966–981. [Google Scholar] [CrossRef] [Green Version]
- Santasusagna, S.; Moreno, I.; Navarro, A.; Martinez Rodenas, F.; Hernandez, R.; Castellano, J.J.; Munoz, C.; Monzo, M. Prognostic Impact of miR-200 Family Members in Plasma and Exosomes from Tumor-Draining versus Peripheral Veins of Colon Cancer Patients. Oncology 2018, 95, 309–318. [Google Scholar] [CrossRef]
- Fischer, C.; Deutsch, T.M.; Feisst, M.; Rippinger, N.; Riedel, F.; Hartkopf, A.D.; Brucker, S.Y.; Domschke, C.; Fremd, C.; Michel, L.; et al. Circulating miR-200 family as predictive markers during systemic therapy of metastatic breast cancer. Arch. Gynecol. Obstet. 2022, 6, e1263412. [Google Scholar] [CrossRef]
- Jing, J.; Xiong, S.; Li, Z.; Wu, J.; Zhou, L.; Gui, J.F.; Mei, J. A feedback regulatory loop involving p53/miR-200 and growth hormone endocrine axis controls embryo size of zebrafish. Sci. Rep. 2015, 5, 15906. [Google Scholar] [CrossRef] [Green Version]
- Alam, F.; Mezhal, F.; El Hasasna, H.; Nair, V.A.; Aravind, S.R.; Saber Ayad, M.; El-Serafi, A.; Abdel-Rahman, W.M. The role of p53-microRNA 200-Moesin axis in invasion and drug resistance of breast cancer cells. Tumour Biol. 2017, 39, 1010428317714634. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Ding, X.; Li, X.; Gao, S.; Yang, Q. 53BP1 suppresses epithelial-mesenchymal transition by downregulating ZEB1 through microRNA-200b/429 in breast cancer. Cancer Sci. 2015, 106, 982–989. [Google Scholar] [CrossRef] [Green Version]
- Benzina, S.; Beauregard, A.P.; Guerrette, R.; Jean, S.; Faye, M.D.; Laflamme, M.; Maicas, E.; Crapoulet, N.; Ouellette, R.J.; Robichaud, G.A. Pax-5 is a potent regulator of E-cadherin and breast cancer malignant processes. Oncotarget 2017, 8, 12052–12066. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, N.; Harquail, J.; Crapoulet, N.; Ouellette, R.J.; Robichaud, G.A. Pax-5 Inhibits Breast Cancer Proliferation Through MiR-215 Up-regulation. Anticancer Res. 2018, 38, 5013–5026. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yao, Q.Y.; Xue, J.S.; Tian, X.Y.; An, Q.M.; Cui, L.X.; Xu, C.; Su, H.; Yang, L.; Feng, Y.Y.; et al. Dexamethasone inhibits pancreatic tumor growth in preclinical models: Involvement of activating glucocorticoid receptor. Toxicol. Appl. Pharmacol. 2020, 401, 115118. [Google Scholar] [CrossRef]
- Rogers, T.J.; Christenson, J.L.; Greene, L.I.; O’Neill, K.I.; Williams, M.M.; Gordon, M.A.; Nemkov, T.; D’Alessandro, A.; Degala, G.D.; Shin, J.; et al. Reversal of Triple-Negative Breast Cancer EMT by miR-200c Decreases Tryptophan Catabolism and a Program of Immunosuppression. Mol. Cancer Res. 2019, 17, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Ha Thi, H.T.; Hong, S. IMP2 and IMP3 cooperate to promote the metastasis of triple-negative breast cancer through destabilization of progesterone receptor. Cancer Lett. 2018, 415, 30–39. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Whitby, R.; Wang, L. Regulation of miR-200c by nuclear receptors PPARalpha, LRH-1 and SHP. Biochem Biophys Res. Commun. 2011, 416, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Hafsi, S.; Candido, S.; Maestro, R.; Falzone, L.; Soua, Z.; Bonavida, B.; Spandidos, D.A.; Libra, M. Correlation between the overexpression of Yin Yang 1 and the expression levels of miRNAs in Burkitt’s lymphoma: A computational study. Oncol. Lett. 2016, 11, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Kopecka, J.; Salaroglio, I.C.; Righi, L.; Libener, R.; Orecchia, S.; Grosso, F.; Milosevic, V.; Ananthanarayanan, P.; Ricci, L.; Capelletto, E.; et al. Loss of C/EBP-beta LIP drives cisplatin resistance in malignant pleural mesothelioma. Lung Cancer 2018, 120, 34–45. [Google Scholar] [CrossRef]
- Shu, C.; Yan, D.; Mo, Y.; Gu, J.; Shah, N.; He, J. Long noncoding RNA lncARSR promotes epithelial ovarian cancer cell proliferation and invasion by association with HuR and miR-200 family. Am. J. Cancer Res. 2018, 8, 981–992. [Google Scholar]
- Chen, J.; Liu, X.; Xu, Y.; Zhang, K.; Huang, J.; Pan, B.; Chen, D.; Cui, S.; Song, H.; Wang, R.; et al. TFAP2C-Activated MALAT1 Modulates the Chemoresistance of Docetaxel-Resistant Lung Adenocarcinoma Cells. Mol. Ther. Nucleic Acids 2019, 14, 567–582. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, S.J.; Fiechter, C.; Burton, J.; Hallion, J.; Paas, M.; Patel, A.; Patel, A.; Rochet, A.; Scheurlen, K.; Gardner, S.; et al. Long non-coding RNA ZFAS1 is a major regulator of epithelial-mesenchymal transition through miR-200/ZEB1/E-cadherin, vimentin signaling in colon adenocarcinoma. Cell Death Discov. 2021, 7, 61. [Google Scholar] [CrossRef]
- Wu, X.; Fan, Y.; Liu, Y.; Shen, B.; Lu, H.; Ma, H. Long Non-Coding RNA CCAT2 Promotes the Development of Esophageal Squamous Cell Carcinoma by Inhibiting miR-200b to Upregulate the IGF2BP2/TK1 Axis. Front. Oncol. 2021, 11, 680642. [Google Scholar] [CrossRef]
- Lin, H.; Yang, L.; Tian, F.; Nie, S.; Zhou, H.; Liu, J.; Chen, W. Up-regulated LncRNA-ATB regulates the growth and metastasis of cholangiocarcinoma via miR-200c signals. Onco. Targets Ther. 2019, 12, 7561–7571. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, Y.; Zuo, C.; Zhang, K.; Lei, X.; Wang, J.; Yang, Y.; Zhang, J.; Ma, K.; Wang, S.; et al. Long Non-Coding RNA H19 Regulates Glioma Cell Growth and Metastasis via miR-200a-Mediated CDK6 and ZEB1 Expression. Front. Oncol. 2021, 11, 757650. [Google Scholar] [CrossRef]
- Kim, S.L.; La, M.T.; Shin, M.W.; Kim, S.W.; Kim, H.K. A novel HDAC1 inhibitor, CBUD1001, exerts anticancer effects by modulating the apoptosis and EMT of colorectal cancer cells. Int. J. Oncol. 2020, 57, 1027–1038. [Google Scholar]
- Neves, R.; Scheel, C.; Weinhold, S.; Honisch, E.; Iwaniuk, K.M.; Trompeter, H.I.; Niederacher, D.; Wernet, P.; Santourlidis, S.; Uhrberg, M. Role of DNA methylation in miR-200c/141 cluster silencing in invasive breast cancer cells. BMC Res. Notes 2010, 3, 219. [Google Scholar] [CrossRef] [Green Version]
- Vrba, L.; Jensen, T.J.; Garbe, J.C.; Heimark, R.L.; Cress, A.E.; Dickinson, S.; Stampfer, M.R.; Futscher, B.W. Role for DNA methylation in the regulation of miR-200c and miR-141 expression in normal and cancer cells. PLoS ONE 2010, 5, e8697. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, E.D.; Bramsen, J.B.; Hulf, T.; Dyrskjot, L.; Ramanathan, R.; Hansen, T.B.; Villadsen, S.B.; Gao, S.; Ostenfeld, M.S.; Borre, M.; et al. Coordinated epigenetic repression of the miR-200 family and miR-205 in invasive bladder cancer. Int. J. Cancer 2011, 128, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Wee, E.J.; Peters, K.; Nair, S.S.; Hulf, T.; Stein, S.; Wagner, S.; Bailey, P.; Lee, S.Y.; Qu, W.J.; Brewster, B.; et al. Mapping the regulatory sequences controlling 93 breast cancer-associated miRNA genes leads to the identification of two functional promoters of the Hsa-mir-200b cluster, methylation of which is associated with metastasis or hormone receptor status in advanced breast cancer. Oncogene 2012, 31, 4182–4195. [Google Scholar] [PubMed] [Green Version]
- Li, B.L.; Lu, W.; Lu, C.; Qu, J.J.; Yang, T.T.; Yan, Q.; Wan, X.P. CpG island hypermethylation-associated silencing of microRNAs promotes human endometrial cancer. Cancer Cell Int. 2013, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Ning, X.; Shi, Z.; Liu, X.; Zhang, A.; Han, L.; Jiang, K.; Kang, C.; Zhang, Q. DNMT1 and EZH2 mediated methylation silences the microRNA-200b/a/429 gene and promotes tumor progression. Cancer Lett. 2015, 359, 198–205. [Google Scholar] [CrossRef]
- Pang, Y.; Liu, J.; Li, X.; Xiao, G.; Wang, H.; Yang, G.; Li, Y.; Tang, S.C.; Qin, S.; Du, N.; et al. MYC and DNMT3A-mediated DNA methylation represses microRNA-200b in triple negative breast cancer. J. Cell Mol. Med. 2018, 22, 6262–6274. [Google Scholar] [CrossRef] [Green Version]
- Kaowinn, S.; Kaewpiboon, C.; Koh, S.S.; Kramer, O.H.; Chung, Y.H. STAT1HDAC4 signaling induces epithelialmesenchymal transition and sphere formation of cancer cells overexpressing the oncogene, CUG2. Oncol. Rep. 2018, 40, 2619–2627. [Google Scholar]
- Chen, D.Q.; Pan, B.Z.; Huang, J.Y.; Zhang, K.; Cui, S.Y.; De, W.; Wang, R.; Chen, L.B. HDAC 1/4-mediated silencing of microRNA-200b promotes chemoresistance in human lung adenocarcinoma cells. Oncotarget 2014, 5, 3333–3349. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Wang, S.; Wang, Y.; Wu, H.; Frank, J.A.; Zhang, Z.; Luo, J. Role of p38gamma MAPK in regulation of EMT and cancer stem cells. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3605–3617. [Google Scholar] [CrossRef]
- Zhang, H.F.; Alshareef, A.; Wu, C.; Li, S.; Jiao, J.W.; Cao, H.H.; Lai, R.; Xu, L.Y.; Li, E.M. Loss of miR-200b promotes invasion via activating the Kindlin-2/integrin β1/AKT pathway in esophageal squamous cell carcinoma: An E-cadherin-independent mechanism. Oncotarget 2015, 6, 28949–28960. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wei, C.; Yang, Y.; Luo, A.; Zhang, X.; Zheng, D.; Lu, X.; Zhang, K.; Duan, X.; Xu, X. Hepatocyte nuclear factor-1beta suppresses the stemness and migration of colorectal cancer cells through promoting miR-200b activity. Mol. Carcinog. 2020, 59, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.N.; Jiang, L.; Jiang, J.H.; Yang, X.; Li, X.Y.; Zeng, J.X.; Shi, R.Y.; Shi, Y.; Pan, X.R.; Han, Z.P.; et al. Hepatocyte nuclear factor-1beta enhances the stemness of hepatocellular carcinoma cells through activation of the Notch pathway. Sci. Rep. 2017, 7, 4793. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, W.; Li, Y.; Huo, Y.; Zhang, H.; Feng, F.; Xi, W.; Zhang, T.; Gao, J.; Yang, F.; et al. MYSM1 inhibits human colorectal cancer tumorigenesis by activating miR-200 family members/CDH1 and blocking PI3K/AKT signaling. J. Exp. Clin. Cancer Res. 2021, 40, 341. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Moya, J.; Baker, A.R.; Slack, F.J.; Santisteban, P. ADAR1-mediated RNA editing is a novel oncogenic process in thyroid cancer and regulates miR-200 activity. Oncogene 2020, 39, 3738–3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, K.L.; Yi, R.; Moorehead, R.A. Transgenic overexpression of the miR-200b/200a/429 cluster inhibits mammary tumor initiation. Transl. Oncol. 2021, 14, 101228. [Google Scholar] [CrossRef]
- Li, D.; Li, Q. MicroRNA-200b-3p restrains gastric cancer cell proliferation, migration, and invasion via C-X-C motif chemokine ligand 12/CXC chemokine receptor 7 axis. Bioengineered 2022, 13, 6509–6520. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, B.; Liang, Y.; Reeves, P.M.; Qu, X.; Ran, C.; Liu, Q.; Callahan, M.V.; Sluder, A.E.; Gelfand, J.A.; et al. Dual blockade of CXCL12-CXCR4 and PD-1-PD-L1 pathways prolongs survival of ovarian tumor-bearing mice by prevention of immunosuppression in the tumor microenvironment. FASEB J. 2019, 33, 6596–6608. [Google Scholar] [CrossRef]
- Qiu, T.Y.; Huang, J.; Wang, L.P.; Zhu, B.S. Inhibition of miR-200b Promotes Angiogenesis in Endothelial Cells by Activating The Notch Pathway. Cell J. 2021, 23, 51–60. [Google Scholar]
- Xue, B.; Chuang, C.H.; Prosser, H.M.; Fuziwara, C.S.; Chan, C.; Sahasrabudhe, N.; Kuhn, M.; Wu, Y.; Chen, J.; Biton, A.; et al. miR-200 deficiency promotes lung cancer metastasis by activating Notch signaling in cancer-associated fibroblasts. Genes Dev. 2021, 35, 1109–1122. [Google Scholar] [CrossRef]
- Sharma, R.; Gogoi, G.; Saikia, S.; Sharma, A.; Kalita, D.J.; Sarma, A.; Limaye, A.M.; Gaur, M.K.; Bhattacharyya, J.; Jaganathan, B.G. BMP4 enhances anoikis resistance and chemoresistance of breast cancer cells through canonical BMP signaling. J. Cell Commun. Signal. 2022, 16, 191–205. [Google Scholar] [CrossRef]
- Tang, Q.; Li, M.; Chen, L.; Bi, F.; Xia, H. miR-200b/c targets the expression of RhoE and inhibits the proliferation and invasion of non-small cell lung cancer cells. Int. J. Oncol. 2018, 53, 1732–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, L.; Li, X. HMGB3 promotes the proliferation and metastasis of glioblastoma and is negatively regulated by miR-200b-3p and miR-200c-3p. Cell Biochem. Funct. 2018, 36, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, I.; Ciccarese, F.; Sharova, E.; Urso, L.; Raimondi, V.; Silic-Benussi, M.; D’Agostino, D.M.; Ciminale, V. The miR-200 Family of microRNAs: Fine Tuners of Epithelial-Mesenchymal Transition and Circulating Cancer Biomarkers. Cancers 2021, 13, 5874. [Google Scholar] [CrossRef]
- Yu, L.; Cao, C.; Li, X.; Zhang, M.; Gu, Q.; Gao, H.; Balic, J.J.; Xu, D.; Zhang, L.; Ying, L.; et al. Complete loss of miR-200 family induces EMT associated cellular senescence in gastric cancer. Oncogene 2022, 41, 26–36. [Google Scholar] [CrossRef]
- Gorecki, I.; Rak, B. The role of microRNAs in epithelial to mesenchymal transition and cancers; focusing on mir-200 family. Cancer Treat. Res. Commun. 2021, 28, 100385. [Google Scholar] [CrossRef] [PubMed]
- Watson, K.L.; Jones, R.A.; Bruce, A.; Moorehead, R.A. The miR-200b/200a/429 cluster prevents metastasis and induces dormancy in a murine claudin-low mammary tumor cell line. Exp. Cell Res. 2018, 369, 17–26. [Google Scholar] [CrossRef]
- Li, J.; Yuan, J.; Yuan, X.; Zhao, J.; Zhang, Z.; Weng, L.; Liu, J. MicroRNA-200b inhibits the growth and metastasis of glioma cells via targeting ZEB2. Int. J. Oncol. 2016, 48, 541–550. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, J.S.; Lee, S.; Lee, H.; Yoon, J.S.; Hong, J.H.; Chun, S.H.; Sun, S.; Won, H.S.; Hong, S.A.; et al. QKI, a miR-200 target gene, suppresses epithelial-to-mesenchymal transition and tumor growth. Int. J. Cancer 2019, 145, 1585–1595. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Song, H.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Wu, T.; Xie, D.; Zhao, J.; et al. The microRNAs miR-200b-3p and miR-429–5p target the LIMK1/CFL1 pathway to inhibit growth and motility of breast cancer cells. Oncotarget 2017, 8, 85276–85289. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Zhu, Z.; Xu, L.; Chen, X.; Li, X.; Lan, T.; Li, W.; Yuan, K.; Zeng, Y. ARHGEF11 promotes proliferation and epithelial-mesenchymal transition of hepatocellular carcinoma through activation of beta-catenin pathway. Aging 2020, 12, 20235–20253. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Ha, J.; Kang, E.; Cho, S. The role of epithelial-mesenchymal transition-regulating transcription factors in anti-cancer drug resistance. Arch. Pharm. Res. 2021, 44, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Kasikci, E.; Aydemir, E.; Bayrak, O.F.; Sahin, F. Inhibition of Migration, Invasion and Drug Resistance of Pancreatic Adenocarcinoma Cells - Role of Snail, Slug and Twist and Small Molecule Inhibitors. Onco. Targets Ther. 2020, 13, 5763–5777. [Google Scholar] [CrossRef] [PubMed]
- Boukouris, A.E.; Zhang, Y.; Saleme, B.; Kinnaird, A.; Zhao, Y.Y.; Liu, Y.; Zervopoulos, S.D.; Das, S.K.; Mittal, R.D.; Haromy, A.; et al. A reversible metabolic stress-sensitive regulation of CRMP2A orchestrates EMT/stemness and increases metastatic potential in cancer. Cell Rep. 2022, 38, 110511. [Google Scholar] [CrossRef] [PubMed]
- Geretto, M.; Pulliero, A.; Rosano, C.; Zhabayeva, D.; Bersimbaev, R.; Izzotti, A. Resistance to cancer chemotherapeutic drugs is determined by pivotal microRNA regulators. Am. J. Cancer Res. 2017, 7, 1350–1371. [Google Scholar]
- Lee, H.; Kim, C.; Kang, H.; Tak, H.; Ahn, S.; Yoon, S.K.; Kuh, H.J.; Kim, W.; Lee, E.K. microRNA-200a-3p increases 5-fluorouracil resistance by regulating dual specificity phosphatase 6 expression. Exp. Mol. Med. 2017, 49, e327. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Oak, P.S.; Wagner, E.; Roidl, A. miR-200c sensitizes breast cancer cells to doxorubicin treatment by decreasing TrkB and Bmi1 expression. PLoS ONE 2012, 7, e50469. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Yu, Y.; Song, Y.; Li, X.; Lan, D.; Zhang, P.; Xiao, Y.; Xing, Y. Activation of BDNF/TrkB pathway promotes prostate cancer progression via induction of epithelial-mesenchymal transition and anoikis resistance. FASEB J. 2020, 34, 9087–9101. [Google Scholar] [CrossRef]
- Jin, H.F.; Wang, J.F.; Song, T.T.; Zhang, J.; Wang, L. MiR-200b Inhibits Tumor Growth and Chemoresistance via Targeting p70S6K1 in Lung Cancer. Front. Oncol. 2020, 10, 643. [Google Scholar] [CrossRef]
- Yu, G.; Jia, B.; Cheng, Y.; Zhou, L.; Qian, B.; Liu, Z.; Wang, Y. MicroRNA-429 sensitizes pancreatic cancer cells to gemcitabine through regulation of PDCD4. Am. J. Transl. Res. 2017, 9, 5048–5055. [Google Scholar]
- Gao, Y.; Zhang, W.; Liu, C.; Li, G. miR-200 affects tamoxifen resistance in breast cancer cells through regulation of MYB. Sci. Rep. 2019, 9, 18844. [Google Scholar] [CrossRef] [PubMed]
- Puhr, M.; Hoefer, J.; Schafer, G.; Erb, H.H.; Oh, S.J.; Klocker, H.; Heidegger, I.; Neuwirt, H.; Culig, Z. Epithelial-to-mesenchymal transition leads to docetaxel resistance in prostate cancer and is mediated by reduced expression of miR-200c and miR-205. Am. J. Pathol. 2012, 181, 2188–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Hu, Q.; Hu, L.X.; Lin, X.R.; Liu, J.Q.; Lin, X.; Dinglin, X.X.; Zeng, J.Y.; Hu, H.; Luo, M.L.; et al. miR-200b regulates epithelial-mesenchymal transition of chemo-resistant breast cancer cells by targeting FN1. Discov. Med. 2017, 24, 75–85. [Google Scholar] [PubMed]
- Liu, J.; Zhang, X.; Huang, Y.; Zhang, Q.; Zhou, J.; Zhang, X.; Wang, X. miR-200b and miR-200c co-contribute to the cisplatin sensitivity of ovarian cancer cells by targeting DNA methyltransferases. Oncol. Lett. 2019, 17, 1453–1460. [Google Scholar] [CrossRef] [Green Version]
- Shindo, T.; Niinuma, T.; Nishiyama, N.; Shinkai, N.; Kitajima, H.; Kai, M.; Maruyama, R.; Tokino, T.; Masumori, N.; Suzuki, H. Epigenetic silencing of miR-200b is associated with cisplatin resistance in bladder cancer. Oncotarget 2018, 9, 24457–24469. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Qu, X.; Zhao, C.; Xu, L.; Hou, K.; Liu, Y.; Zhang, N.; Feng, J.; Shi, S.; Zhang, L.; et al. FEN1 mediates miR-200a methylation and promotes breast cancer cell growth via MET and EGFR signaling. FASEB J. 2019, 33, 10717–10730. [Google Scholar] [CrossRef]
- Aita, V.M.; Liang, X.H.; Murty, V.V.; Pincus, D.L.; Yu, W.; Cayanis, E.; Kalachikov, S.; Gilliam, T.C.; Levine, B. Cloning and genomic organization of beclin 1, a candidate tumor suppressor gene on chromosome 17q21. Genomics 1999, 59, 59–65. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, S.; Xie, P.; Zhang, B.; Yu, M.; Yan, J.; Jin, L.; Zhang, W.; Zhou, B.; Li, X.; et al. Tumor-derived PD1 and PD-L1 could promote hepatocellular carcinoma growth through autophagy induction in vitro. Biochem. Biophys. Res. Commun. 2022, 605, 82–89. [Google Scholar] [CrossRef]
- Lai, M.; Liu, L.; Zhu, L.; Feng, W.; Luo, J.; Liu, Y.; Deng, S. Triptolide reverses epithelial-mesenchymal transition in glioma cells via inducing autophagy. Ann. Transl. Med. 2021, 9, 1304. [Google Scholar] [CrossRef]
- Hu, Y.L.; Jahangiri, A.; Delay, M.; Aghi, M.K. Tumor cell autophagy as an adaptive response mediating resistance to treatments such as antiangiogenic therapy. Cancer Res. 2012, 72, 4294–4299. [Google Scholar] [CrossRef] [Green Version]
- Fedorova, O.; Daks, A.; Parfenyev, S.; Shuvalov, O.; Netsvetay, S.; Vasileva, J.; Gudovich, A.; Golotin, V.; Semenov, O.; Petukhov, A.; et al. Zeb1-mediated autophagy enhances resistance of breast cancer cells to genotoxic drugs. Biochem. Biophys. Res. Commun. 2022, 589, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Yeon, M.; Kim, Y.; Pathak, D.; Kwon, E.; Kim, D.Y.; Jeong, M.S.; Jung, H.S.; Jeoung, D. The CAGE-MiR-181b-5p-S1PR1 Axis Regulates Anticancer Drug Resistance and Autophagy in Gastric Cancer Cells. Front. Cell Dev. Biol. 2021, 9, 666387. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Zhang, Y.; Malmrose, P.K.; Losh, H.A.; Newtson, A.M.; Devor, E.J.; Thiel, K.W.; Leslie, K.K. Blocking autophagy overcomes resistance to dual histone deacetylase and proteasome inhibition in gynecologic cancer. Cell Death Dis. 2022, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Kong, Y.; Shao, C.; Yue, X.; Wang, Z.; Zhang, N. Heme oxygenase-1 induction mediates chemoresistance of breast cancer cells to pharmorubicin by promoting autophagy via PI3K/Akt pathway. J. Cell Mol. Med. 2018, 22, 5311–5321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, J.; Zeng, X.; Nie, M.; Chen, W.; Wang, Y.; Luan, J.; Zhu, Z.; Chang, X.; Ju, D.; et al. Targeting the autophagy promoted antitumor effect of T-DM1 on HER2-positive gastric cancer. Cell Death Dis. 2021, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Jiao, L.; Lin, C.; Lu, C.; Zhang, K.; Hu, C.; Ye, J.; Zhang, D.; Wu, H.; et al. Protective autophagy decreases osimertinib cytotoxicity through regulation of stem cell-like properties in lung cancer. Cancer Lett. 2019, 452, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Feng, B.; Chen, Y.; Huang, G.; Wang, R.; Chen, L.; Song, H. MiR-200b regulates autophagy associated with chemoresistance in human lung adenocarcinoma. Oncotarget 2015, 6, 32805–32820. [Google Scholar] [CrossRef]
- Kim, H.; Kim, Y.; Jeoung, D. DDX53 Promotes Cancer Stem Cell-Like Properties and Autophagy. Mol. Cells 2017, 40, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Alsadi, N.; Mallet, J.F.; Matar, C. miRNA-200b Signature in the Prevention of Skin Cancer Stem Cells by Polyphenol-enriched Blueberry Preparation. J. Cancer Prev. 2021, 26, 162–173. [Google Scholar] [CrossRef]
- Rankovic, B.; Zidar, N.; Zlajpah, M.; Bostjancic, E. Epithelial-Mesenchymal Transition-Related MicroRNAs and Their Target Genes in Colorectal Cancerogenesis. J. Clin. Med. 2019, 8, 1603. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; He, R.; Geng, L.; Yuan, J.; Fan, H. Ginsenoside Rg3 Alleviates Cisplatin Resistance of Gastric Cancer Cells Through Inhibiting SOX2 and the PI3K/Akt/mTOR Signaling Axis by Up-Regulating miR-429. Front. Genet. 2022, 13, 823182. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, H.; Park, D.; Lee, Y.S.; Choe, J.; Hahn, J.H.; Lee, H.; Kim, Y.M.; Jeoung, D. Cancer/testis antigen CAGE exerts negative regulation on p53 expression through HDAC2 and confers resistance to anti-cancer drugs. J. Biol. Chem. 2010, 285, 25957–25968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeon, M.; Byun, J.; Kim, H.; Kim, M.; Jung, H.S.; Jeon, D.; Kim, Y.; Jeoung, D. CAGE Binds to Beclin1, Regulates Autophagic Flux and CAGE-Derived Peptide Confers Sensitivity to Anti-cancer Drugs in Non-small Cell Lung Cancer Cells. Front. Oncol. 2018, 8, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Park, D.; Kim, H.; Choi, M.; Lee, H.; Lee, Y.S.; Choe, J.; Kim, Y.M.; Jeoung, D. miR-200b and cancer/testis antigen CAGE form a feedback loop to regulate the invasion and tumorigenic and angiogenic responses of a cancer cell line to microtubule-targeting drugs. J. Biol. Chem. 2013, 288, 36502–36518. [Google Scholar] [CrossRef] [Green Version]
- Gyvyte, U.; Lukosevicius, R.; Inciuraite, R.; Streleckiene, G.; Gudoityte, G.; Bekampyte, J.; Valentini, S.; Salteniene, V.; Ruzgys, P.; Satkauskas, S.; et al. The Role of miR-375-3p and miR-200b-3p in Gastrointestinal Stromal Tumors. Int J. Mol Sci 2020, 21, 5151. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.; Park, D.; Han, M.; Lee, H.; Lee, Y.S.; Choe, J.; Kim, Y.M.; Jeoung, D. miR-217 and CAGE form feedback loop and regulates the response to anti-cancer drugs through EGFR and HER2. Oncotarget 2016, 7, 10297–10321. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, Y.; Goh, H.; Jeoung, D. Histone Deacetylase-3/CAGE Axis Targets EGFR Signaling and Regulates the Response to Anti-Cancer Drugs. Mol. Cells 2016, 39, 229–241. [Google Scholar]
- Kim, Y.; Kim, H.; Park, H.; Park, D.; Lee, H.; Lee, Y.S.; Choe, J.; Kim, Y.M.; Jeoung, D. miR-326-histone deacetylase-3 feedback loop regulates the invasion and tumorigenic and angiogenic response to anti-cancer drugs. J. Biol. Chem. 2014, 289, 28019–28039. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.; Kim, Y.; Jung, H.S.; Jeoung, D. Role of HDAC3-miRNA-CAGE Network in Anti-Cancer Drug-Resistance. Int. J. Mol. Sci 2018, 20, 51. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, D.R.; Spoelstra, N.S.; Howe, E.N.; Nordeen, S.K.; Richer, J.K. MicroRNA-200c mitigates invasiveness and restores sensitivity to microtubule-targeting chemotherapeutic agents. Mol. Cancer Ther. 2009, 8, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Shadbad, M.A.; Asadzadeh, Z.; Derakhshani, A.; Hosseinkhani, N.; Mokhtarzadeh, A.; Baghbanzadeh, A.; Hajiasgharzadeh, K.; Brunetti, O.; Argentiero, A.; Racanelli, V.; et al. A scoping review on the potentiality of PD-L1-inhibiting microRNAs in treating colorectal cancer: Toward single-cell sequencing-guided biocompatible-based delivery. Biomed. Pharmacother. 2021, 143, 112213. [Google Scholar] [CrossRef] [PubMed]
- Mu, L.; Wang, Y.; Su, H.; Lin, Y.; Sui, W.; Yu, X.; Lv, Z. HIF1A-AS2 Promotes the Proliferation and Metastasis of Gastric Cancer Cells Through miR-429/PD-L1 Axis. Dig. Dis. Sci. 2021, 66, 4314–4325. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wang, K.; Huang, X.; Zhao, Z.; Zhao, Z. LncRNA MALAT1 contributes to non-small cell lung cancer progression via modulating miR-200a-3p/programmed death-ligand 1 axis. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419859699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Li, S.; Xue, J.; Qi, M.; Liu, X.; Huang, Y.; Hu, J.; Dong, H.; Ling, K. PD-L1 tumor-intrinsic signaling and its therapeutic implication in triple-negative breast cancer. JCI Insight 2021, 6, e131458. [Google Scholar] [CrossRef]
- Lv, J.; Guo, T.; Qu, X.; Che, X.; Li, C.; Wang, S.; Gong, J.; Wu, P.; Liu, Y.; Liu, Y.; et al. PD-L1 Under Regulation of miR-429 Influences the Sensitivity of Gastric Cancer Cells to TRAIL by Binding of EGFR. Front. Oncol. 2020, 10, 1067. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.; Park, D.; Lee, H.; Lee, Y.S.; Choe, J.; Kim, Y.M.; Jeon, D.; Jeoung, D. The pentapeptide Gly-Thr-Gly-Lys-Thr confers sensitivity to anti-cancer drugs by inhibition of CAGE binding to GSK3beta and decreasing the expression of cyclinD1. Oncotarget 2017, 8, 13632–13651. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Altwegg, K.A.; Liu, J.; Weintraub, S.T.; Chen, Y.; Lai, Z.; Sareddy, G.R.; Viswanadhapalli, S.; Vadlamudi, R.K. Global Genomic and Proteomic Analysis Identified Critical Pathways Modulated by Proto-Oncogene PELP1 in TNBC. Cancers 2022, 14, 930. [Google Scholar] [CrossRef]
- Zhang, D.; Dai, J.; Pan, Y.; Wang, X.; Qiao, J.; Sasano, H.; Zhao, B.; McNamara, K.M.; Guan, X.; Liu, L.; et al. Overexpression of PELP1 in Lung Adenocarcinoma Promoted E2 Induced Proliferation, Migration and Invasion of the Tumor Cells and Predicted a Worse Outcome of the Patients. Pathol. Oncol. Res. 2021, 27, 582443. [Google Scholar] [CrossRef]
- Roy, S.S.; Gonugunta, V.K.; Bandyopadhyay, A.; Rao, M.K.; Goodall, G.J.; Sun, L.Z.; Tekmal, R.R.; Vadlamudi, R.K. Significance of PELP1/HDAC2/miR-200 regulatory network in EMT and metastasis of breast cancer. Oncogene 2014, 33, 3707–3716. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, L.; Liu, F.; Pan, J.; Yao, Z.; Lin, Y.; Yang, Y.; Xiong, X.; Li, K.; Yang, Y.; et al. Targeting PELP1 Attenuates Angiogenesis and Enhances Chemotherapy Efficiency in Colorectal Cancer. Cancers 2022, 14, 383. [Google Scholar] [CrossRef]
- Krishnan, S.R.; Nair, B.C.; Sareddy, G.R.; Roy, S.S.; Natarajan, M.; Suzuki, T.; Peng, Y.; Raj, G.; Vadlamudi, R.K. Novel role of PELP1 in regulating chemotherapy response in mutant p53-expressing triple negative breast cancer cells. Breast Cancer Res. Treat. 2015, 150, 487–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Feng, C.; Zhang, W.; Long, Y.; Fa, X. The epigenetic silencing of microRNA-433 facilitates the malignant phenotypes of non-small cell lung cancer by targeting CREB1. Am. J. Transl. Res. 2021, 13, 12302–12317. [Google Scholar] [PubMed]
- Stavast, C.J.; van Zuijen, I.; Karkoulia, E.; Ozcelik, A.; van Hoven-Beijen, A.; Leon, L.G.; Voerman, J.S.A.; Janssen, G.M.C.; van Veelen, P.A.; Burocziova, M.; et al. The tumor suppressor MIR139 is silenced by POLR2M to promote AML oncogenesis. Leukemia 2022, 36, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, L.; Hou, L.; Tang, X. LncRNA SNHG1 promotes tumor progression and cisplatin resistance through epigenetically silencing miR-381 in breast cancer. Bioengineered 2021, 12, 9239–9250. [Google Scholar] [CrossRef]
- Farah, E.; Zhang, Z.; Utturkar, S.M.; Liu, J.; Ratliff, T.L.; Liu, X. Targeting DNMTs to Overcome Enzalutamide Resistance in Prostate Cancer. Mol. Cancer Ther. 2022, 21, 193–205. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, Z.; Chai, D.; Gao, Y.; Li, T.; Ma, Y.; Li, J. DNA methyltransferase 1 inhibits microRNA-497 and elevates GPRC5A expression to promote chemotherapy resistance and metastasis in breast cancer. Cancer Cell Int. 2022, 22, 112. [Google Scholar] [CrossRef]
- Sato, H.; Shien, K.; Tomida, S.; Okayasu, K.; Suzawa, K.; Hashida, S.; Torigoe, H.; Watanabe, M.; Yamamoto, H.; Soh, J.; et al. Targeting the miR-200c/LIN28B axis in acquired EGFR-TKI resistance non-small cell lung cancer cells harboring EMT features. Sci. Rep. 2017, 7, 40847. [Google Scholar] [CrossRef]
- Cao, Q.; You, X.; Xu, L.; Wang, L.; Chen, Y. PAQR3 suppresses the growth of non-small cell lung cancer cells via modulation of EGFR-mediated autophagy. Autophagy 2020, 16, 1236–1247. [Google Scholar] [CrossRef]
- Dent, P.; Booth, L.; Poklepovic, A.; Von Hoff, D.; Martinez, J.; Zhou, Y.; Hancock, J.F. Osimertinib-resistant NSCLC cells activate ERBB2 and YAP/TAZ and are killed by neratinib. Biochem. Pharmacol. 2021, 190, 114642. [Google Scholar] [CrossRef]
- Zhou, G.; Zhang, F.; Guo, Y.; Huang, J.; Xie, Y.; Yue, S.; Chen, M.; Jiang, H.; Li, M. miR-200c enhances sensitivity of drug-resistant non-small cell lung cancer to gefitinib by suppression of PI3K/Akt signaling pathway and inhibites cell migration via targeting ZEB1. Biomed. Pharmacother. 2017, 85, 113–119. [Google Scholar] [CrossRef]
- Della Corte, C.M.; Malapelle, U.; Vigliar, E.; Pepe, F.; Troncone, G.; Ciaramella, V.; Troiani, T.; Martinelli, E.; Belli, V.; Ciardiello, F.; et al. Efficacy of continuous EGFR-inhibition and role of Hedgehog in EGFR acquired resistance in human lung cancer cells with activating mutation of EGFR. Oncotarget 2017, 8, 23020–23032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Maitah, M.Y.; Ginnebaugh, K.R.; Li, Y.; Bao, B.; Gadgeel, S.M.; Sarkar, F.H. Inhibition of Hedgehog signaling sensitizes NSCLC cells to standard therapies through modulation of EMT-regulating miRNAs. J. Hematol. Oncol. 2013, 6, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoo, S.R.; Yim, Y.; Kim, Y.K.; Park, I.S.; Na, H.K.; Lee, J.; Jang, H.; Won, C.; Hong, S.; Kim, S.Y.; et al. High-throughput chemical screening to discover new modulators of microRNA expression in living cells by using graphene-based biosensor. Sci. Rep. 2018, 8, 11413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaby, O.; Laga, R.; Sedlacek, O. Therapeutic targeting of non-coding RNAs in cancer. Biochem. J. 2017, 474, 4219–4251. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Type | Sample Type/Size | Expression of miR-200s | Application | Reference |
|---|---|---|---|---|
| Renal cell carcinoma | Eighty tumor tissues and adjacent tissues | ↓ miR-200b in tumor tissues | Prediction of metastasis, prognosis | [18] |
| Glioma | Eighty-nine glioma tissues and 41 non-tumor tissues | ↓ miR-200b in tumor tissues | Diagnosis and prognosis | [19] |
| Clear cell renal cell carcinoma | Normal tissues (n = 23), primary tumor tissues (n = 194), metastatic tumor tissues (n= 10) | ↓ miR-141, ↓ miR-200b in metastatic tumor tissues High levels of miR-200b and miR-200c → longer progression-free survival | Diagnosis and prognosis | [20] |
| Gastric cancer | Sixty tumor tissues and normal control tissues | High level of mR-200b → high overall survival | Diagnosis and prognosis | [21] |
| Hepatocellular carcinoma | 371 tumor tissues | ↓ miR-200b ↑ HMG3B in tumor tissues —miR-200b targets HMG3B | Diagnosis | [22] |
| Bladder cancer | Tumor tissues from 1150 patients | High level of the miR-200 family → better prognosis | Prognosis | [23] |
| Non-small cell lung cancer (NSCLC) | Tumor tissues from 60 NSCLC patients | High level of miR-200b → high progression-free survival | Prognosis in response to systemic immune therapy | [24] |
| Hepatocellular carcinoma | Whole blood from thirty patients and normal controls | ↓ miR-141 ↓ miR-200a in serum of cancer patients | Diagnosis | [27] |
| NSCLC | Fifty cancer patients and 30 normal controls | ↓ miR-200b in serum of cancer patients | Diagnosis | [28] |
| Breast cancer | Whole blood from early (137) and metastatic patients (110) | High levels of miR-200b and miR-200c in metastatic patients compared to early patients High level of miR-200b → shorter overall survival | Diagnosis and prognosis | [29] |
| Colon cancer | Plasma and exosomes from fifty resected patients | Low levels of miR-141 and miR-200c → longer overall survival | Diagnosis | [30] |
| Breast cancer | Serum (n = 47) from metastatic cancer patients | High levels of the miR-200 family (miR-141, miR-200a, miR-200b, miR-429) → reduction in overall survival and progression-free survival | Prognosis | [31] |
| miR-200 Family | Mechanism | Target | Enhances Sensitivity to | Cancer | Reference |
|---|---|---|---|---|---|
| miR-200a | Signaling | DUSP6 | 5-FU | Hepatocellular carcinoma | [86] |
| miR-200b | mTOR pathway | P70S6K1 | Cisplatin | Lung cancer | [89] |
| FAK/Src signaling | FN1 | Doxorubicin | Breast cancer | [93] | |
| Autophagy | ATG-12 | Docetaxel | Lung cancer | [107] | |
| Autophagy | ATG-5 | Cisplatin Docetaxel | Breast cancer | [108] | |
| Autophagy/cancer stemness | CAGE | Microtubule-targeting agents | Melanoma | [112] | |
| Signaling | Hh signaling | Erlotinib | Lung cancer | [142] | |
| miR-200b/-200c | EMT | C-MYB | Tamoxifen | Breast cancer | [91] |
| Epigenetic modification | DNMT1 | Cisplatin | Ovarian cancer | [94] | |
| miR-200c | Signaling | TrkB | Doxorubicin | Breast cancer | [87] |
| EMT | E-cadherin, SNAIL | Docetaxel | Prostate cancer | [92] | |
| EMT | TUBB3, ZEB1/ZEB2 | Microtubule-targeting agents | Ovarian, Breast cancer | [120] | |
| Signaling | PI3K/Akt | Gefitinib | Lung cancer | [137] | |
| miR-200c/-141 | EMT | E-cadherin, Vimentin | Oxaliplatin | Ovarian cancer | [95] |
| miR-429 | Cell death | PDCD4 | Gemcitabine | Pancreatic cancer | [90] |
| Cancer stemness | SOX2 | Cisplatin | Gastric cancer | [111] | |
| Immune suppression | PD-L1 | TRAIL | Gastric cancer | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, H.; Shim, K.; Jeoung, D. Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics. Int. J. Mol. Sci. 2022, 23, 5881. https://doi.org/10.3390/ijms23115881
Jo H, Shim K, Jeoung D. Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics. International Journal of Molecular Sciences. 2022; 23(11):5881. https://doi.org/10.3390/ijms23115881
Chicago/Turabian StyleJo, Hyein, Kyeonghee Shim, and Dooil Jeoung. 2022. "Potential of the miR-200 Family as a Target for Developing Anti-Cancer Therapeutics" International Journal of Molecular Sciences 23, no. 11: 5881. https://doi.org/10.3390/ijms23115881