
Monoclonal Antibodies and Invasive Aspergillosis: Diagnostic and Therapeutic Perspectives

 , ,
, ,
Abstract
:1. Introduction
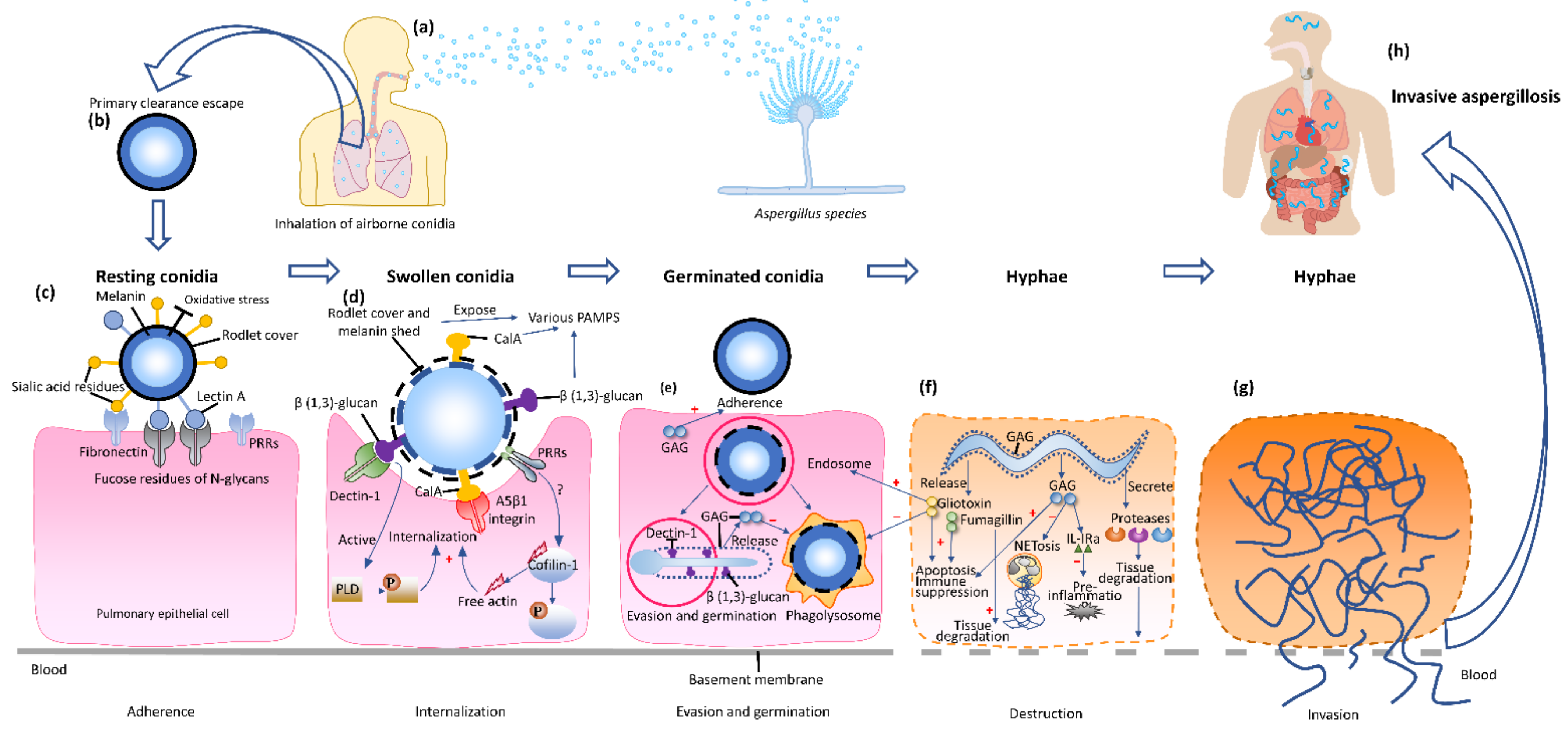
2. Pathogenesis of Invasive Aspergillosis
3. Monoclonal Antibodies and Diagnosis of Invasive Aspergillosis
3.1. Challenges of Invasive Aspergillosis Diagnosis
3.2. Monoclonal Antibody-Based Diagnostic Approaches for Invasive Aspergillosis
3.2.1. The Detection of Aspergillus Antigens in Blood and Urine Using Monoclonal Antibodies
3.2.2. The Use of Aspergillus Monoclonal Antibodies for Microscopy and Imaging
- Microscopy
- Imaging
- Bioluminescence Imaging
- Immune PET/MRI
4. Monoclonal Antibodies and Invasive Aspergillosis Therapy
4.1. Challenges of Invasive Aspergillosis Therapy
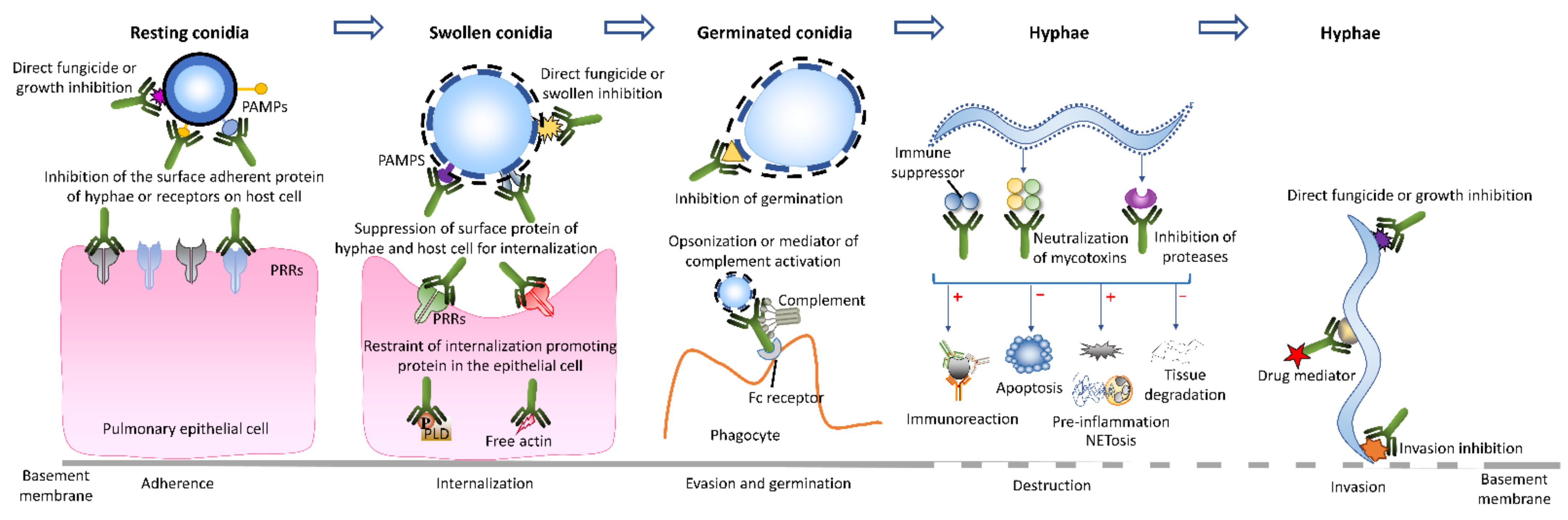
4.2. Anti Aspergillus Activity of Monoclonal Antibodies
4.3. Monoclonal Antibody-Based Immunotherapeutic Modes for Invasive Aspergillosis
5. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IA | invasive aspergillosis |
| mAbs | monoclonal antibodies |
| mAb | monoclonal antibody |
| HSCT | allogeneic hematopoietic stem cell transplant |
| SOT | solid organ transplant |
| PAMPs | pathogen-associated molecular patterns |
| PRRs | pattern recognition receptors |
| Cal A | calcineurin A |
| GAG | galactosaminogalactan |
| NETs | neutrophil extracellular traps |
| ROS | reactive oxygen species |
| PLD | phospholipase D |
| EORTC/MSGERC | European Organization for Research and Treatment of Cancer and the Mycoses Study Group Education and Research Consortium |
| PCR | polymerase chain reaction |
| GM | galactomannan |
| BALF | bronchoalveolar lavage fluid |
| LFD | lateral flow device |
| IHC | immunohistochemistry |
| IF | immunofluorescence |
| PET | positron emission tomography |
| SPECT | single-photon emission computed tomography |
| BLI | bioluminescence imaging |
| LSFM | light sheet fluorescence microscopy |
| ADEPT | antibody-directed enzyme prodrug technology |
| FDA | Food and Drug Administration. |
References
- Imbert, S.; Cassaing, S.; Bonnal, C.; Normand, A.C.; Gabriel, F.; Costa, D.; Blaize, M.; Lachaud, L.; Hasseine, L.; Kristensen, L.; et al. Invasive aspergillosis due to Aspergillus cryptic species: A prospective multicentre study. Mycoses 2021, 64, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Cuenca-Estrella, M. Antifungal susceptibility profile of cryptic species of Aspergillus. Mycopathologia 2014, 178, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Azoulay, E.; Russell, L.; Van De Louw, A.; Metaxa, V.; Bauer, P.; Povoa, P.; Montero, J.G.; Loeches, I.M.; Mehta, S.; Puxty, K.; et al. Diagnosis of severe respiratory infections in immunocompromised patients. Intensive Care Med. 2020, 46, 298–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenks, J.D.; Gangneux, J.P.; Schwartz, I.S.; Alastruey-Izquierdo, A.; Lagrou, K.; Thompson Iii, G.R.; Lass-Florl, C.; Hoenigl, M.; European Confederation of Medical Mycology Council Investigators. Diagnosis of breakthrough fungal infections in the clinical mycology laboratory: An ECMM consensus statement. J. Fungi 2020, 6, 216. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Fortwendel, J.R.; Rogers, P.D. Emerging threat of triazole-resistant Aspergillus fumigatus. J. Antimicrob. Chemother. 2019, 74, 835–842. [Google Scholar] [CrossRef]
- Segrelles-Calvo, G.; Araujo, G.R.S.; Llopis-Pastor, E.; Carrillo, J.; Hernandez-Hernandez, M.; Rey, L.; Rodriguez Melean, N.; Escribano, I.; Anton, E.; Zamarro, C.; et al. Prevalence of opportunistic invasive aspergillosis in COVID-19 patients with severe pneumonia. Mycoses 2021, 64, 144–151. [Google Scholar] [CrossRef]
- Autier, B.; Prattes, J.; White, P.L.; Valerio, M.; Machado, M.; Price, J.; Egger, M.; Gangneux, J.P.; Hoenigl, M. Aspergillus lateral flow assay with digital reader for the diagnosis of COVID-19-associated pulmonary aspergillosis (CAPA): A multicenter study. J. Clin. Microbiol. 2022, 60, e0168921. [Google Scholar] [CrossRef]
- Marr, K.A.; Platt, A.; Tornheim, J.A.; Zhang, S.X.; Datta, K.; Cardozo, C.; Garcia-Vidal, C. Aspergillosis complicating severe coronavirus disease. Emerg. Infect. Dis. 2021, 27, 18–25. [Google Scholar] [CrossRef]
- Sharma, A.; Mishra, T.; Kumar, N.; Soubani, A.O. Influenza-associated aspergillosis: Nationwide trends, predictors and outcomes from 2005 to 2014. Chest 2020, 158, 1857–1866. [Google Scholar] [CrossRef]
- Chen, W.; Li, M.; Younis, M.H.; Barnhart, T.E.; Jiang, D.; Sun, T.; Lang, J.M.; Engle, J.W.; Zhou, M.; Cai, W. ImmunoPET of trophoblast cell-surface antigen 2 (Trop-2) expression in pancreatic cancer. Eur. J. Nucl. Med. Mol. Imaging 2022, 49, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Bagati, A.; Kumar, S.; Jiang, P.; Pyrdol, J.; Zou, A.E.; Godicelj, A.; Mathewson, N.D.; Cartwright, A.N.R.; Cejas, P.; Brown, M.; et al. Integrin alphavbeta6-TGFbeta-SOX4 pathway drives immune evasion in triple-negative breast cancer. Cancer Cell 2021, 39, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Carnasciali, A.; Amoriello, R.; Bonechi, E.; Mazzoni, A.; Ravagli, C.; Doumett, S.; Cappiello, L.; D’Elios, M.M.; Baldi, G.; Ballerini, C. T cell delivery of nanoparticles-bound anti-CD20 monoclonal antibody: Successful B cell depletion in the spinal cord during experimental autoimmune encephalomyelitis. J. Neuroimmune Pharmacol. 2021, 16, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Menzies-Gow, A.; Corren, J.; Bourdin, A.; Chupp, G.; Israel, E.; Wechsler, M.E.; Brightling, C.E.; Griffiths, J.M.; Hellqvist, A.; Bowen, K.; et al. Tezepelumab in adults and adolescents with severe, uncontrolled asthma. N. Engl. J. Med. 2021, 384, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Sirivichayakul, C.; Limkittikul, K.; Chanthavanich, P.; Yoksan, S.; Ratchatatat, A.; Lim, J.K.; Arunsodsai, W.; Sabchareon, A. Monoclonal antibody-based capture ELISA in the diagnosis of previous dengue infection. Virol. J. 2019, 16, 125. [Google Scholar] [CrossRef] [PubMed]
- Weber-Lima, M.M.; Prado-Costa, B.; Becker-Finco, A.; Costa, A.O.; Billilad, P.; Furst, C.; De Moura, J.F.; Alvarenga, L.M. Acanthamoeba spp. monoclonal antibody against a CPA2 transporter: A promising molecular tool for acanthamoebiasis diagnosis and encystment study. Parasitology 2020, 147, 1678–1688. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Drabek, D.; Okba, N.M.A.; Van Haperen, R.; Osterhaus, A.; Van Kuppeveld, F.J.M.; Haagmans, B.L.; Grosveld, F.; Bosch, B.J. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat. Commun. 2020, 11, 2251. [Google Scholar] [CrossRef]
- Kreye, J.; Reincke, S.M.; Kornau, H.C.; Sanchez-Sendin, E.; Corman, V.M.; Liu, H.; Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.D.; et al. A therapeutic non-self-reactive SARS-CoV-2 antibody protects from lung pathology in a COVID-19 hamster model. Cell 2020, 183, 1058–1069. [Google Scholar] [CrossRef]
- Park, S.J.; Mehrad, B. Innate immunity to Aspergillus species. Clin. Microbiol. Rev. 2009, 22, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Kuek, L.E.; Lee, R.J. First contact: The role of respiratory cilia in host-pathogen interactions in the airways. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L603–L619. [Google Scholar] [CrossRef]
- Latgé, J.-P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef] [PubMed]
- Strickland, A.B.; Shi, M. Mechanisms of fungal dissemination. Cell Mol. Life Sci. 2021, 78, 3219–3238. [Google Scholar] [CrossRef] [PubMed]
- Van De Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latge, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Bayry, J.; Bozza, S.; Kniemeyer, O.; Perruccio, K.; Elluru, S.R.; Clavaud, C.; Paris, S.; Brakhage, A.A.; Kaveri, S.V.; et al. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature 2009, 460, 1117–1121. [Google Scholar] [CrossRef]
- Chai, L.Y.; Netea, M.G.; Sugui, J.; Vonk, A.G.; Van De Sande, W.W.; Warris, A.; Kwon-Chung, K.J.; Kullberg, B.J. Aspergillus fumigatus conidial melanin modulates host cytokine response. Immunobiology 2010, 215, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, I.; Dupres, V.; Stephen-Victor, E.; Guijarro, J.I.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.Y.; Lafont, F.; Latge, J.P.; et al. Role of hydrophobins in Aspergillus fumigatus. J. Fungi 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Warwas, M.L.; Watson, J.N.; Bennet, A.J.; Moore, M.M. Structure and role of sialic acids on the surface of Aspergillus fumigatus conidiospores. Glycobiology 2007, 17, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D.C. Molecular mechanism of Aspergillus fumigatus adherence to host constituents. Curr. Opin. Microbiol. 2011, 14, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Houser, J.; Komarek, J.; Kostlanova, N.; Cioci, G.; Varrot, A.; Kerr, S.C.; Lahmann, M.; Balloy, V.; Fahy, J.V.; Chignard, M.; et al. A soluble fucose-specific lectin from Aspergillus fumigatus conidia--structure, specificity and possible role in fungal pathogenicity. PLoS ONE 2013, 8, e83077. [Google Scholar] [CrossRef]
- Rodriguez-De La Noval, C.; Ruiz Mendoza, S.; De Souza Goncalves, D.; Da Silva Ferreira, M.; Honorato, L.; Peralta, J.M.; Nimrichter, L.; Guimaraes, A.J. Protective efficacy of lectin-Fc (IgG) fusion proteins in vitro and in a pulmonary aspergillosis in vivo model. J. Fungi 2020, 6, 250. [Google Scholar] [CrossRef]
- Liu, H.; Lee, M.J.; Solis, N.V.; Phan, Q.T.; Swidergall, M.; Ralph, B.; Ibrahim, A.S.; Sheppard, D.C.; Filler, S.G. Aspergillus fumigatus CalA binds to integrin alpha5beta1 and mediates host cell invasion. Nat. Microbiol. 2016, 2, 16211. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, D.; Namperumalsamy, V.P.; Prajna, L.; Kuppamuthu, D. Human corneal epithelial cells internalize Aspergillus flavus spores by actin-mediated endocytosis. Infect. Immun. 2021, 89, e00794-20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Chen, F.Y.; Liu, X.Y.; Han, X.L.; Hui, Y.S.; Su, X.T.; Chen, Y.; Sun, Y.S.; Han, L. Gliotoxin induces cofilin phosphorylation to promote actin cytoskeleton dynamics and internalization of Aspergillus fumigatus into type II human pneumocyte cells. Front. Microbiol. 2019, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.D.; Chen, F.Y.; Pan, W.H.; Yu, R.T.; Tian, S.G.; Han, G.G.; Fang, H.Q.; Wang, S.; Zhao, J.Y.; Li, X.P.; et al. Gliotoxin promotes Aspergillus fumigatus internalization into type II human pneumocyte A549 cells by inducing host phospholipase D activation. Microb. Infect. 2014, 16, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Gravelat, F.N.; Beauvais, A.; Liu, H.; Lee, M.J.; Snarr, B.D.; Chen, D.; Xu, W.; Kravtsov, I.; Hoareau, C.M.; Vanier, G.; et al. Aspergillus galactosaminogalactan mediates adherence to host constituents and conceals hyphal beta-glucan from the immune system. PLoS Pathog. 2013, 9, e1003575. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Geller, A.M.; Bamford, N.C.; Liu, H.; Gravelat, F.N.; Snarr, B.D.; Le Mauff, F.; Chabot, J.; Ralph, B.; Ostapska, H.; et al. Deacetylation of fungal exopolysaccharide mediates adhesion and biofilm formation. Mbio 2016, 7, e00252-16. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.J.; Liu, H.; Barker, B.M.; Snarr, B.D.; Gravelat, F.N.; Al Abdallah, Q.; Gavino, C.; Baistrocchi, S.R.; Ostapska, H.; Xiao, T.L.; et al. The fungal exopolysaccharide galactosaminogalactan mediates virulence by enhancing resistance to neutrophil extracellular traps. PLoS Pathog. 2015, 11, e1005187. [Google Scholar] [CrossRef]
- Gresnigt, M.S.; Bozza, S.; Becker, K.L.; Joosten, L.A.B.; Abdollahi-Roodsaz, S.; Van Der Berg, W.B.; Dinarello, C.A.; Netea, M.G.; Fontaine, T.; De Luca, A.; et al. A polysaccharide virulence factor from Aspergillus fumigatus elicits anti-inflammatory effects through induction of interleukin-1 receptor antagonist. PLoS Pathog. 2014, 10, e1003936. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Meis, J.F.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.L.H.; Rosa-Milani, E.; Brunaldi, M.O.; Maffei, C.M.L. Murine model of invasive pulmonary Aspergillosis: Follow-up of tissue injury, fungal burden and mortality with distinct elastase production strains. J. Mycol. Med. 2019, 29, 112–119. [Google Scholar] [CrossRef]
- Rowley, J.; Namvar, S.; Gago, S.; Labram, B.; Bowyer, P.; Richardson, M.D.; Herrick, S.E. Differential proinflammatory responses to Aspergillus fumigatus by airway epithelial cells in vitro are protease dependent. J. Fungi 2021, 7, 468. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, L.; Akbarsha, M.A.; Ruckmani, K. In vitro study on aspects of molecular mechanisms underlying invasive aspergillosis caused by gliotoxin and fumagillin, alone and in combination. Sci. Rep. 2020, 10, 14473. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Santiago, L.; Vidal-Garcia, M.; Redrado, S.; Lanuza, P.; Comas, L.; Domingo, M.P.; Rezusta, A.; Galvez, E.M. Preparations for invasion: Modulation of host lung immunity during pulmonary aspergillosis by gliotoxin and other fungal secondary metabolites. Front. Immunol. 2018, 9, 2549. [Google Scholar] [CrossRef]
- Stanzani, M.; Orciuolo, E.; Lewis, R.; Kontoyiannis, D.P.; Martins, S.L.; St John, L.S.; Komanduri, K.V. Aspergillus fumigatus suppresses the human cellular immune response via gliotoxin-mediated apoptosis of monocytes. Blood 2005, 105, 2258–2265. [Google Scholar] [CrossRef] [Green Version]
- Waring, P.; Eichner, R.D.; Mullbacher, A.; Sjaarda, A. Gliotoxin induces apoptosis in macrophages unrelated to its antiphagocytic properties. J. Biol. Chem. 1988, 263, 18493–18499. [Google Scholar] [CrossRef]
- Stevanovic, J.; Stanimirovic, Z.; Radakovic, M.; Stojic, V. In vitro evaluation of the clastogenicity of fumagillin. Environ. Mol. Mutagen. 2008, 49, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Fallon, J.P.; Reeves, E.P.; Kavanagh, K. The Aspergillus fumigatus toxin fumagillin suppresses the immune response of Galleria mellonella larvae by inhibiting the action of haemocytes. Microbiology 2011, 157, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- Guruceaga, X.; Ezpeleta, G.; Mayayo, E.; Sueiro-Olivares, M.; Abad-Diaz-De-Cerio, A.; Aguirre Urizar, J.M.; Liu, H.G.; Wiemann, P.; Bok, J.W.; Filler, S.G.; et al. A possible role for fumagillin in cellular damage during host infection by Aspergillus fumigatus. Virulence 2018, 9, 1548–1561. [Google Scholar] [CrossRef] [Green Version]
- Davies, G.; Rolle, A.M.; Maurer, A.; Spycher, P.R.; Schillinger, C.; Solouk-Saran, D.; Hasenberg, M.; Weski, J.; Fonslet, J.; Dubois, A.; et al. Towards translational immunoPET/MR imaging of invasive pulmonary Aspergillosis: The humanised monoclonal antibody JF5 detects Aspergillus lung infections in vivo. Theranostics 2017, 7, 3398–3414. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Chen, S.C.; Kauffman, C.A.; Steinbach, W.J.; Baddley, J.W.; Verweij, P.E.; Clancy, C.J.; Wingard, J.R.; Lockhart, S.R.; Groll, A.H.; et al. Revision and update of the consensus definitions of invasive fungal disease from the European organization for research and treatment of cancer and the mycoses study group education and research consortium. Clin. Infect. Dis. 2020, 71, 1367–1376. [Google Scholar] [CrossRef] [Green Version]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Florl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24 (Suppl. 1), e1–e38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, C.R. Breaking the mould—Novel diagnostic and therapeutic strategies for invasive pulmonary aspergillosis in the immune deficient patient. Expert Rev. Clin. Immunol. 2014, 10, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Marr, K.A.; Anaissie, E.J.; Azie, N.; Quan, S.P.; Meier-Kriesche, H.U.; Apewokin, S.; Horn, D.L. Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J. Infect. 2012, 65, 453–464. [Google Scholar] [CrossRef]
- Sarwar, M.; Gardezi, S.A.H.; Zaman, G.; Ikram, A.; Satti, L.; Khadim, M.T. Evaluation of galactomannan and beta-d-glucan assays for the diagnosis of invasive aspergillosis in clinically suspected cases. J. Pak. Med. Assoc. 2020, 70, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, K.; Seybold, U.; Ormanns, S.; Horns, H.; Wagener, J. Evaluation of a novel Aspergillus antigen enzyme-linked immunosorbent assay. J. Clin. Microbiol. 2019, 57, e00136-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, M.; Penziner, S.; Dichtl, K.; Gornicec, M.; Kriegl, L.; Krause, R.; Khong, E.; Mehta, S.; Vargas, M.; Gianella, S.; et al. Performance of the euroimmun Aspergillus antigen ELISA for the diagnosis of invasive pulmonary Aspergillosis in bronchoalveolar lavage fluid. J. Clin. Microbiol. 2022, 60, e00215-22. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Singh, O.; Prattes, J.; Hoenigl, M.; Sheppard, P.W.; Thornton, C.R. asp f I: Old enemies but new opportunities for urine-based detection of invasive pulmonary Aspergillosis using lateral-flow technology. J. Fungi 2020, 7, 19. [Google Scholar] [CrossRef]
- Jenks, J.D.; Mehta, S.R.; Taplitz, R.; Aslam, S.; Reed, S.L.; Hoenigl, M. Point-of-care diagnosis of invasive aspergillosis in non-neutropenic patients: Aspergillus galactomannan lateral flow assay versus Aspergillus-specific lateral flow device test in bronchoalveolar lavage. Mycoses 2019, 62, 230–236. [Google Scholar] [CrossRef]
- Jenks, J.D.; Hoenigl, M. Point-of-care diagnostics for invasive aspergillosis: Nearing the finish line. Expert Rev. Mol. Diagn. 2020, 20, 1009–1017. [Google Scholar] [CrossRef]
- Fisher, B.T.; Westling, T.; Boge, C.L.K.; Zaoutis, T.E.; Dvorak, C.C.; Nieder, M.; Zerr, D.M.; Wingard, J.R.; Villaluna, D.; Esbenshade, A.J.; et al. Prospective evaluation of galactomannan and (1-->3) beta-d-glucan assays as diagnostic tools for invasive fungal disease in children, adolescents, and young adults with acute myeloid leukemia receiving fungal prophylaxis. J. Pediatric Infect. Dis Soc. 2021, 10, 864–871. [Google Scholar] [CrossRef]
- Calero, A.L.; Alonso, R.; Gadea, I.; Vega, M.D.M.; Garcia, M.M.; Munoz, P.; Machado, M.; Bouza, E.; Garcia-Rodriguez, J. Comparison of the performance of two galactomannan detection tests: Platelia Aspergillus Ag and Aspergillus galactomannan ag virclia monotest. Microbiol. Spectr. 2022, 10, e02626-21. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.; Husain, S. Current state of the diagnosis of invasive pulmonary Aspergillosis in lung transplantation. Front. Microbiol. 2018, 9, 3273. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, H.; Zhang, Y.; Huang, M.; He, Q.; Li, P.; Zhang, F.; Shi, Y.; Su, X. Diagnostic value of galactomannan antigen test in serum and bronchoalveolar lavage fluid samples from patients with nonneutropenic invasive pulmonary Aspergillosis. J. Clin. Microbiol. 2017, 55, 2153–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamoth, F. Galactomannan and 1,3-beta-d-glucan testing for the diagnosis of invasive Aspergillosis. J. Fungi 2016, 2, 22. [Google Scholar] [CrossRef] [PubMed]
- Leeflang, M.M.; Debets-Ossenkopp, Y.J.; Wang, J.; Visser, C.E.; Scholten, R.J.; Hooft, L.; Bijlmer, H.A.; Reitsma, J.B.; Zhang, M.; Bossuyt, P.M.; et al. Galactomannan detection for invasive aspergillosis in immunocompromised patients. Cochrane Database Syst. Rev. 2015, 2015, CD007394. [Google Scholar] [CrossRef]
- Hao, W.; Pan, Y.X.; Ding, Y.Q.; Xiao, S.; Yin, K.; Wang, Y.D.; Qiu, L.W.; Zhang, Q.L.; Woo, P.C.; Lau, S.K.; et al. Well-characterized monoclonal antibodies against cell wall antigen of Aspergillus species improve immunoassay specificity and sensitivity. Clin. Vaccine Immunol. 2008, 15, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Y.; Cai, J.P.; Qiu, L.W.; Hao, W.; Pan, Y.X.; Tung, E.T.; Lau, C.C.; Woo, P.C.; Lau, S.K.; Yuen, K.Y.; et al. Development of monoclonal antibody-based galactomannoprotein antigen-capture ELISAs to detect Aspergillus fumigatus infection in the invasive aspergillosis rabbit models. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2943–2950. [Google Scholar] [CrossRef]
- Ansari, S.; Mousavi, A.; Safarnejad, M.R.; Farrokhi, N.; Alavi, S.M.; Schillberg, S.; Nolke, G. Selection and characterization of two monoclonal antibodies specific for the Aspergillus flavus major antigenic cell wall protein Aflmp1. Fungal Biol. 2021, 125, 621–629. [Google Scholar] [CrossRef]
- Lian, X.; Chambers, S.; Lewis, J.G.; Scott-Thomas, A.; Bhatia, M. Two Monoclonal antibodies that specifically recognize Aspergillus cell wall antigens and can detect circulating antigens in infected mice. Int. J. Mol. Sci. 2022, 23, 252. [Google Scholar] [CrossRef]
- Prattes, J.; Flick, H.; Pruller, F.; Koidl, C.; Raggam, R.B.; Palfner, M.; Eigl, S.; Buzina, W.; Zollner-Schwetz, I.; Thornton, C.R.; et al. Novel tests for diagnosis of invasive aspergillosis in patients with underlying respiratory diseases. Am. J. Respir. Crit. Care Med. 2014, 190, 922–929. [Google Scholar] [CrossRef]
- Thornton, C.; Johnson, G.; Agrawal, S. Detection of invasive pulmonary aspergillosis in haematological malignancy patients by using lateral-flow technology. J. Vis. Exp. 2012, 61, e3721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenks, J.D.; Mehta, S.R.; Taplitz, R.; Law, N.; Reed, S.L.; Hoenigl, M. Bronchoalveolar lavage Aspergillus Galactomannan lateral flow assay versus Aspergillus-specific lateral flow device test for diagnosis of invasive pulmonary Aspergillosis in patients with hematological malignancies. J. Infect. 2019, 78, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Heldta, S.; Prattes, J.; Eigl, S.; Spiess, B.; Flick, H.; Rabensteiner, J.; Johnson, G.; Pruller, F.; Wolfler, A.; Niedrist, T.; et al. Diagnosis of invasive aspergillosis in hematological malignancy patients: Performance of cytokines, Asp LFD, and Aspergillus PCR in same day blood and bronchoalveolar lavage samples. J. Infect. 2018, 77, 235–241. [Google Scholar] [CrossRef]
- Mercier, T.; Schauwvlieghe, A.; De Kort, E.; Dunbar, A.; Reynders, M.; Guldentops, E.; Rijnders, B.; Verweij, P.E.; Lagrou, K.; Maertens, J. Diagnosing invasive pulmonary aspergillosis in hematology patients: A retrospective multicenter evaluation of a novel lateral flow device. J. Clin. Microbiol. 2019, 57, e01913-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, K.A.; Kauffman, C.A.; Zhou, S.; Miceli, M.H. Clinical application of Aspergillus lateral flow device in bronchoalveolar lavage fluid of patients with classic risk factors for invasive pulmonary aspergillosis. Mycoses 2019, 62, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Datta, K.; Mehta, S.; Ostrander, D.B.; Rock, M.; Francis, J.; Feldmesser, M. Urine antigen detection as an aid to diagnose invasive Aspergillosis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 1705–1711. [Google Scholar] [CrossRef] [Green Version]
- Hoenigl, M.; Eigl, S.; Heldt, S.; Duettmann, W.; Thornton, C.; Prattes, J. Clinical evaluation of the newly formatted lateral-flow device for invasive pulmonary aspergillosis. Mycoses 2018, 61, 40–43. [Google Scholar] [CrossRef] [Green Version]
- Held, J.; Schmidt, T.; Thornton, C.R.; Kotter, E.; Bertz, H. Comparison of a novel Aspergillus lateral-flow device and the Platelia (R) galactomannan assay for the diagnosis of invasive aspergillosis following haematopoietic stem cell transplantation. Infection 2013, 41, 1163–1169. [Google Scholar] [CrossRef]
- White, P.L.; Parr, C.; Thornton, C.; Barnes, R.A. Evaluation of real-time PCR, galactomannan enzyme-linked immunosorbent assay (ELISA), and a novel lateral-flow device for diagnosis of invasive aspergillosis. J. Clin. Microbiol. 2013, 51, 1510–1516. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.K.; Mauger, J.; McGowan, K.L. Immunohistochemical detection of Aspergillus species in pediatric tissue samples. Am. J. Clin. Pathol. 2004, 121, 18–25. [Google Scholar] [CrossRef]
- Phillips, P.; Weiner, M.H. Invasive aspergillosis diagnosed by immunohistochemistry with monoclonal and polyclonal reagents. Hum. Pathol. 1987, 18, 1015–1024. [Google Scholar] [CrossRef]
- Pierard, G.E.; Arrese Estrada, J.; Pierard-Franchimont, C.; Thiry, A.; Stynen, D. Immunohistochemical expression of galactomannan in the cytoplasm of phagocytic cells during invasive aspergillosis. Am. J. Clin. Pathol. 1991, 96, 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, P.E.; Smedts, F.; Poot, T.; Bult, P.; Hoogkamp-Korstanje, J.A.; Meis, J.F. Immunoperoxidase staining for identification of Aspergillus species in routinely processed tissue sections. J. Clin. Pathol. 1996, 49, 798–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amich, J.; Mokhtari, Z.; Strobel, M.; Vialetto, E.; Sheta, D.; Yu, Y.; Hartweg, J.; Kalleda, N.; Jarick, K.J.; Brede, C.; et al. Three-dimensional light sheet fluorescence microscopy of lungs to dissect local host immune-Aspergillus fumigatus interactions. mBio 2020, 11, e02752-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henneberg, S.; Hasenberg, A.; Maurer, A.; Neumann, F.; Bornemann, L.; Gonzalez-Menendez, I.; Kraus, A.; Hasenberg, M.; Thornton, C.R.; Pichler, B.J.; et al. Antibody-guided in vivo imaging of Aspergillus fumigatus lung infections during antifungal azole treatment. Nat. Commun. 2021, 12, 1707. [Google Scholar] [CrossRef] [PubMed]
- Rolle, A.M.; Hasenberg, M.; Thornton, C.R.; Solouk-Saran, D.; Mann, L.; Weski, J.; Maurer, A.; Fischer, E.; Spycher, P.R.; Schibli, R.; et al. ImmunoPET/MR imaging allows specific detection of Aspergillus fumigatus lung infection in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E1026–E1033. [Google Scholar] [CrossRef] [Green Version]
- Gunzer, M.; Thornton, C.R.; Beziere, N. Advances in the in vivo molecular imaging of invasive Aspergillosis. J. Fungi 2020, 6, 338. [Google Scholar] [CrossRef]
- Gammon, S.T.; Foje, N.; Brewer, E.M.; Owers, E.; Downs, C.A.; Budde, M.D.; Leevy, W.M.; Helms, M.N. Preclinical anatomical, molecular, and functional imaging of the lung with multiple modalities. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 306, L897–L914. [Google Scholar] [CrossRef] [Green Version]
- Andreu, N.; Zelmer, A.; Wiles, S. Noninvasive biophotonic imaging for studies of infectious disease. FEMS Microbiol. Rev. 2011, 35, 360–394. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Álvarez, E.; Stamatakis, K.; Punzón, C.; Álvarez-Velilla, R.; Tejería, A.; Escudero-Martínez, J.M.; Pérez-Pertejo, Y.; Fresno, M.; Balaña-Fouce, R.; Reguera, R.M. Infrared fluorescent imaging as a potent tool for in vitro, ex vivo and in vivo models of visceral leishmaniasis. PLoS Negl. Trop. Dis. 2015, 9, e0003666. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhang, Z.; Chen, P.; Long, N.; Lu, L.; Sang, H. In vitro and in vivo efficacy of a synergistic combination of itraconazole and verapamil against Aspergillus fumigatus. Front. Microbiol. 2019, 10, 1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, E.; Packard, A.B. Radioactive transition metals for imaging and therapy. Chem. Rev. 2019, 119, 870–901. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.; Cardinale, J.; Aulsebrook, M.L.; Gasser, G.; Mindt, T.L. An overview of PET radiochemistry, part 2: Radiometals. J. Nucl. Med. 2018, 59, 1500–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.J.; Lewis, J.S. Current status and future challenges for molecular imaging. Philos. Trans. A Math. Phys. Eng. Sci. 2017, 375, 20170023. [Google Scholar] [CrossRef] [PubMed]
- Badr, C.E.; Tannous, B.A. Bioluminescence imaging: Progress and applications. Trends Biotechnol. 2011, 29, 624–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, M.; Jouvion, G.; Droin-Bergere, S.; Dussurget, O.; Nicola, M.A.; Ibrahim-Granet, O. Bioluminescent Aspergillus fumigatus, a new tool for drug efficiency testing and in vivo monitoring of invasive aspergillosis. Appl. Environ. Microbiol. 2008, 74, 7023–7035. [Google Scholar] [CrossRef] [Green Version]
- Velde, G.V.; Wiehr, S. Fungal imaging. In Imaging Infections; Jain, S.K., Ed.; Springer International Publishing: Baltimore, MD, USA, 2017; pp. 173–183. [Google Scholar]
- Ibrahim-Granet, O.; Jouvion, G.; Hohl, T.M.; Droin-Bergere, S.; Philippart, F.; Kim, O.Y.; Adib-Conquy, M.; Schwendener, R.; Cavaillon, J.M.; Brock, M. In vivo bioluminescence imaging and histopathopathologic analysis reveal distinct roles for resident and recruited immune effector cells in defense against invasive aspergillosis. BMC Microbiol. 2010, 10, 105. [Google Scholar] [CrossRef]
- Galiger, C.; Brock, M.; Jouvion, G.; Savers, A.; Parlato, M.; Ibrahim-Granet, O. Assessment of efficacy of antifungals against Aspergillus fumigatus: Value of real-time bioluminescence imaging. Antimicrob. Agents Chemother. 2013, 57, 3046–3059. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Rosenkrans, Z.T.; Liu, J.; Huang, G.; Luo, Q.Y.; Cai, W. ImmunoPET: Concept, design, and applications. Chem. Rev. 2020, 120, 3787–3851. [Google Scholar] [CrossRef]
- Lechermann, L.M.; Lau, D.; Attili, B.; Aloj, L.; Gallagher, F.A. In vivo cell tracking using PET: Opportunities and challenges for clinical translation in oncology. Cancers 2021, 13, 4042. [Google Scholar] [CrossRef]
- Thornton, C.R. Development of an immunochromatographic lateral-flow device for rapid serodiagnosis of invasive aspergillosis. Clin. Vaccine Immunol. 2008, 15, 1095–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poelman, J.; Himmelreich, U.; Vanherp, L.; Zhai, L.C.; Hillen, A.; Holvoet, B.; Belderbos, S.; Brock, M.; Maertens, J.; Vande Velde, G.; et al. A multimodal imaging approach enables in vivo assessment of antifungal treatment in a mouse model of invasive pulmonary Aspergillosis. Antimicrob. Agents Chemother. 2018, 62, e00240-18. [Google Scholar] [CrossRef] [Green Version]
- Leroux, S.; Ullmann, A.J. Management and diagnostic guidelines for fungal diseases in infectious diseases and clinical microbiology: Critical appraisal. Clin. Microbiol. Infect. 2013, 19, 1115–1121. [Google Scholar] [CrossRef] [Green Version]
- Shi, N.; Zheng, Q.; Zhang, H. Molecular dynamics investigations of binding mechanism for triazoles inhibitors to CYP51. Front. Mol. Biosci. 2020, 7, 586540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Lu, J.; Duan, X.; Chen, J.; Jin, X.; Lin, Z.; Pang, Y.; Wang, X.; Lou, H.; Chang, W. Rimonabant potentiates the antifungal activity of amphotericin B by increasing cellular oxidative stress and cell membrane permeability. FEMS Yeast Res. 2021, 21, foab016. [Google Scholar] [CrossRef] [PubMed]
- Aruanno, M.; Glampedakis, E.; Lamoth, F. Echinocandins for the Treatment of Invasive Aspergillosis: From Laboratory to Bedside. Antimicrob. Agents Chemother. 2019, 63, e00399-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamaris, G.A.; Lewis, R.E.; Chamilos, G.; May, G.S.; Safdar, A.; Walsh, T.J.; Raad, I.I.; Kontoyiannis, D.P. Caspofungin-mediated beta-glucan unmasking and enhancement of human polymorphonuclear neutrophil activity against Aspergillus and non-Aspergillus hyphae. J. Infect. Dis. 2008, 198, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Torosantucci, A.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Palma, A.S.; Liu, Y.; Mignogna, G.; Maras, B.; Colone, M.; Stringaro, A.; et al. Protection by anti-beta-glucan antibodies is associated with restricted beta-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE 2009, 4, e5392. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, A.K.; Kumar, R.; Kumar, A.; Shukla, P.K. A monoclonal IgM directed against immunodominant catalase B of cell wall of Aspergillus fumigatus exerts anti-A. fumigatus activities. Mycoses 2009, 52, 524–533. [Google Scholar] [CrossRef]
- Kumar, A.; Shukla, P.K. A monoclonal antibody against glycoproteins of Aspergillus fumigatus shows anti-adhesive potential. Microb. Pathog. 2015, 79, 24–30. [Google Scholar] [CrossRef]
- Matveev, A.L.; Krylov, V.B.; Khlusevich, Y.A.; Baykov, I.K.; Yashunsky, D.V.; Emelyanova, L.A.; Tsvetkov, Y.E.; Karelin, A.A.; Bardashova, A.V.; Wong, S.S.W.; et al. Novel mouse monoclonal antibodies specifically recognizing β-1,3-D-glucann antigen. PLoS ONE 2019, 14, e0215535. [Google Scholar] [CrossRef]
- Chauvin, D.; Hust, M.; Schutte, M.; Chesnay, A.; Parent, C.; Moreira, G.; Arroyo, J.; Sanz, A.B.; Pugniere, M.; Martineau, P.; et al. Targeting Aspergillus fumigatus crf transglycosylases with neutralizing antibody is relevant but not sufficient to erase fungal burden in a neutropenic rat model. Front. Microbiol. 2019, 10, 600. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, S.; Ebel, F. Monoclonal antibodies as tools to combat fungal infections. J. Fungi 2020, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.K.; Shukla, P.K. A novel monoclonal antibody against enolase antigen of Aspergillus fumigatus protects experimental aspergillosis in mice. FEMS Microbiol. Lett. 2019, 366, fnz015. [Google Scholar] [CrossRef]
- Xu, X.Y.; Chen, F.; Sun, H.; Chen, C.; Zhao, B.L. Important factors mediates the adhesion of Aspergillus fumigatus to alveolar epithelial cells with E-cadherin. Am. J. Transl. Res. 2016, 8, 2419–2425. [Google Scholar] [PubMed]
- Xu, X.Y.; Shi, Y.; Zhang, P.P.; Zhang, F.; Shen, Y.Y.; Su, X.; Zhao, B.L. E-cadherin mediates adhesion and endocytosis of Aspergillus fumigatus blastospores in human epithelial cells. Chin. Med. J. 2012, 125, 617–621. [Google Scholar] [PubMed]
- Bowen, A.; Wear, M.P.; Cordero, R.J.; Oscarson, S.; Casadevall, A. A monoclonal antibody to cryptococcus neoformans glucuronoxylomannan manifests hydrolytic activity for both peptides and polysaccharides. J. Biol. Chem. 2017, 292, 417–434. [Google Scholar] [CrossRef] [Green Version]
- Frosco, M.B.; Chase, T., Jr.; Macmillan, J.D. The effect of elastase-specific monoclonal and polyclonal antibodies on the virulence of Aspergillus fumigatus in immunocompromised mice. Mycopathologia 1994, 125, 65–76. [Google Scholar] [CrossRef]
- Cenci, E.; Mencacci, A.; Spreca, A.; Montagnoli, C.; Bacci, A.; Perruccio, K.; Velardi, A.; Magliani, W.; Conti, S.; Polonelli, L.; et al. Protection of killer antiidiotypic antibodies against early invasive aspergillosis in a murine model of allogeneic T-cell-depleted bone marrow transplantation. Infect. Immun. 2002, 70, 2375–2382. [Google Scholar] [CrossRef] [Green Version]
- Moragues, M.D.; Omaetxebarria, M.J.; Elguezabal, N.; Sevilla, M.J.; Conti, S.; Polonelli, L.; Ponton, J. A monoclonal antibody directed against a Candida albicans cell wall mannoprotein exerts three anti-C. albicans activities. Infect. Immun. 2003, 71, 5273–5279. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, A.K.; Kavishwar, A.; Shiva Keshava, G.B.; Shukla, P.K. Monoclonal immunoglobulin G1 directed against Aspergillus fumigatus cell wall glycoprotein protects against experimental murine aspergillosis. Clin. Diagn. Lab. Immunol. 2005, 12, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Wharton, R.E.; Stefanov, E.K.; King, R.G.; Kearney, J.F. Antibodies generated against Streptococci protect in a mouse model of disseminated aspergillosis. J. Immunol. 2015, 194, 4387–4396. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Habiel, D.M.; Coelho, A.L.; Hogaboam, C.M. Axl receptor blockade protects from invasive pulmonary aspergillosis in mice. J. Immunol. 2014, 193, 3559–3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; De Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P.; et al. A novel glyco-conjugate vaccine against fungal pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Nosanchuk, J.D.; Dadachova, E. Radioimmunotherapy of fungal diseases: The therapeutic potential of cytocidal radiation delivered by antibody targeting fungal cell surface antigens. Front. Microbiol. 2011, 2, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appel, E.; Vallon-Eberhard, A.; Rabinkov, A.; Brenner, O.; Shin, I.; Sasson, K.; Shadkchan, Y.; Osherov, N.; Jung, S.; Mirelman, D. Therapy of murine pulmonary aspergillosis with antibody-alliinase conjugates and alliin. Antimicrob. Agents Chemother. 2010, 54, 898–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagshawe, K.D.; Sharma, S.K.; Springer, C.J.; Rogers, G.T. Antibody directed enzyme prodrug therapy (ADEPT). A review of some theoretical, experimental and clinical aspects. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 1994, 5, 879–891. [Google Scholar] [CrossRef]
- Karam, S.D.; Raben, D. Radioimmunotherapy for the treatment of head and neck cancer. Lancet Oncol. 2019, 20, E404–E416. [Google Scholar] [CrossRef]
- Dadachova, E.; Nakouzi, A.; Bryan, R.A.; Casadevall, A. Ionizing radiation delivered by specific antibody is therapeutic against a fungal infection. Proc. Natl. Acad. Sci. USA 2003, 100, 10942–10947. [Google Scholar] [CrossRef] [Green Version]
- Segal, B.H.; Herbrecht, R.; Stevens, D.A.; Ostrosky-Zeichner, L.; Sobel, J.; Viscoli, C.; Walsh, T.J.; Maertens, J.; Patterson, T.F.; Perfect, J.R.; et al. Defining responses to therapy and study outcomes in clinical trials of invasive fungal diseases: Mycoses Study Group and European Organization for Research and Treatment of Cancer consensus criteria. Clin. Infect. Dis. 2008, 47, 674–683. [Google Scholar] [CrossRef]
- Food and Drug Administration. Development Considerations of Antifungal Drugs to Address Unmet Medical Need. Available online: https://www.fda.gov/media/141400/download (accessed on 11 May 2022).
- Rex, J.H. Antifungal Drugs to Address Unmet Medical Need. Available online: https://www.fda.gov/media/141074/download (accessed on 11 May 2022).



{kind=link}
{kind=link}
| Diagnostic Methods | Advantages | Disadvantages |
|---|---|---|
| Histopathology and microbiology | Gold standard Pathologic changes of the tissue Morphology of the Aspergillus | Invasive operation High requirements for specimen quality Technology dependent on the technician Time-consuming False-negative |
| Fungal biomarker assay | Early detection Non-invasive Various sample resources Platform widely available Rapid turnaround time | False-positive False-negative Unknown pathogen species Unknown infection site |
| Aspergillus polymerase chain reaction (PCR) test | Specific species Various sample resources Rapid turnaround time | Lack of standardization Contamination can be problematic |
| CT scanning | Fast and non-invasive Location of infection site Lesion size and number | Non-specific Pathogen not identified Viability of pathogen not indicated |
| Serological antibody test | Easily performed on readily accessible samples | False-negative in immunocompromised host |
| Therapeutic Effects | MAb | Subclass | References |
|---|---|---|---|
| Fungal growth inhibition/fungicidal activity (in vitro) | C7, K10, A9, Mab-7, SMB19, R-5, MS112-IIB1, YW327.6S2, 3G11 and 5H5 | IgM, IgG, IgG1, IgG3 | [110,112,113,115,120,121,122,123,124] |
| Fungal growth inhibition/fungicidal activity (in vivo) | K10, A9, 2G8, R-5, 3G11 and 5H5 | IgM, IgG1, IgG2b, IgG3 | [112,115,118,123,125,126] |
| Germination suppression (in vitro) | K10, A9, 2G8, R-5, 3G11 and 5H5 | IgM, IgG1, IgG2b, IgG3 | [112,115,118,123,125,126] |
| Attachment inhibition (in vitro) | 2G8, Mab-7, AK-14 | IgG2b, IgM | [109,110,111,125] |
| Protease inhibition (in vivo) | BB11, MB8, KD5, GD11, and CCIII 19 | IgG1 and IgG2a | [119] |
| Immunological enhancement (in vitro and in vivo) | A9, SMB19, MS112-IIB1, 3G11, 5H5, YW327.6S2 | IgG1, IgM, IgG1, IgG3, IgG | [112,113,122,123,124] |
| Drug mediator (in vitro and in vivo) | MPS5.44 | IgM | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, X.; Scott-Thomas, A.; Lewis, J.G.; Bhatia, M.; MacPherson, S.A.; Zeng, Y.; Chambers, S.T. Monoclonal Antibodies and Invasive Aspergillosis: Diagnostic and Therapeutic Perspectives. Int. J. Mol. Sci. 2022, 23, 5563. https://doi.org/10.3390/ijms23105563
Lian X, Scott-Thomas A, Lewis JG, Bhatia M, MacPherson SA, Zeng Y, Chambers ST. Monoclonal Antibodies and Invasive Aspergillosis: Diagnostic and Therapeutic Perspectives. International Journal of Molecular Sciences. 2022; 23(10):5563. https://doi.org/10.3390/ijms23105563
Chicago/Turabian StyleLian, Xihua, Amy Scott-Thomas, John G. Lewis, Madhav Bhatia, Sean A. MacPherson, Yiming Zeng, and Stephen T. Chambers. 2022. "Monoclonal Antibodies and Invasive Aspergillosis: Diagnostic and Therapeutic Perspectives" International Journal of Molecular Sciences 23, no. 10: 5563. https://doi.org/10.3390/ijms23105563