
Glycosylation of Receptor Binding Domain of SARS-CoV-2 S-Protein Influences on Binding to Immobilized DNA Aptamers

 , , , , and
, , , , and
Abstract
:
1. Introduction
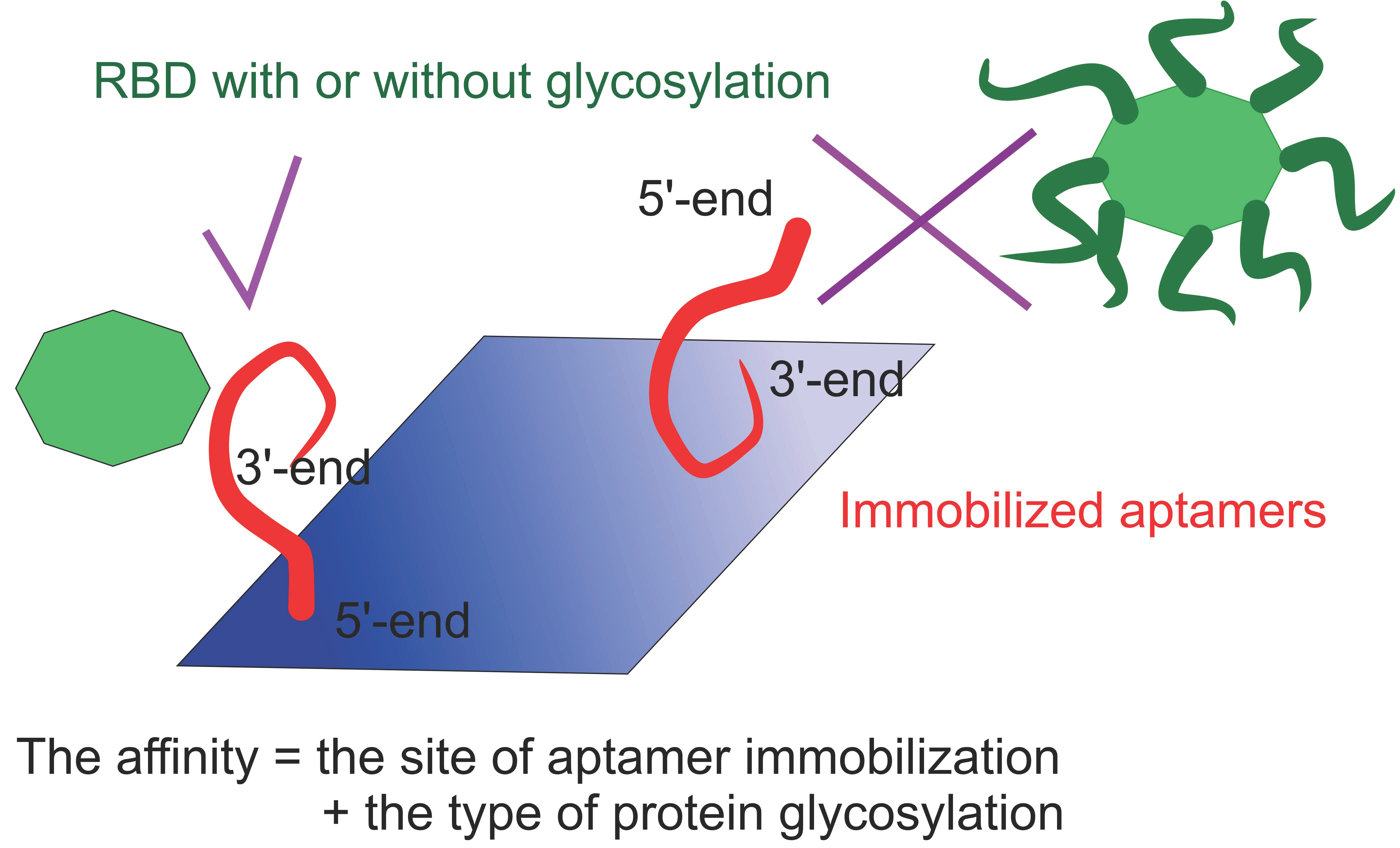
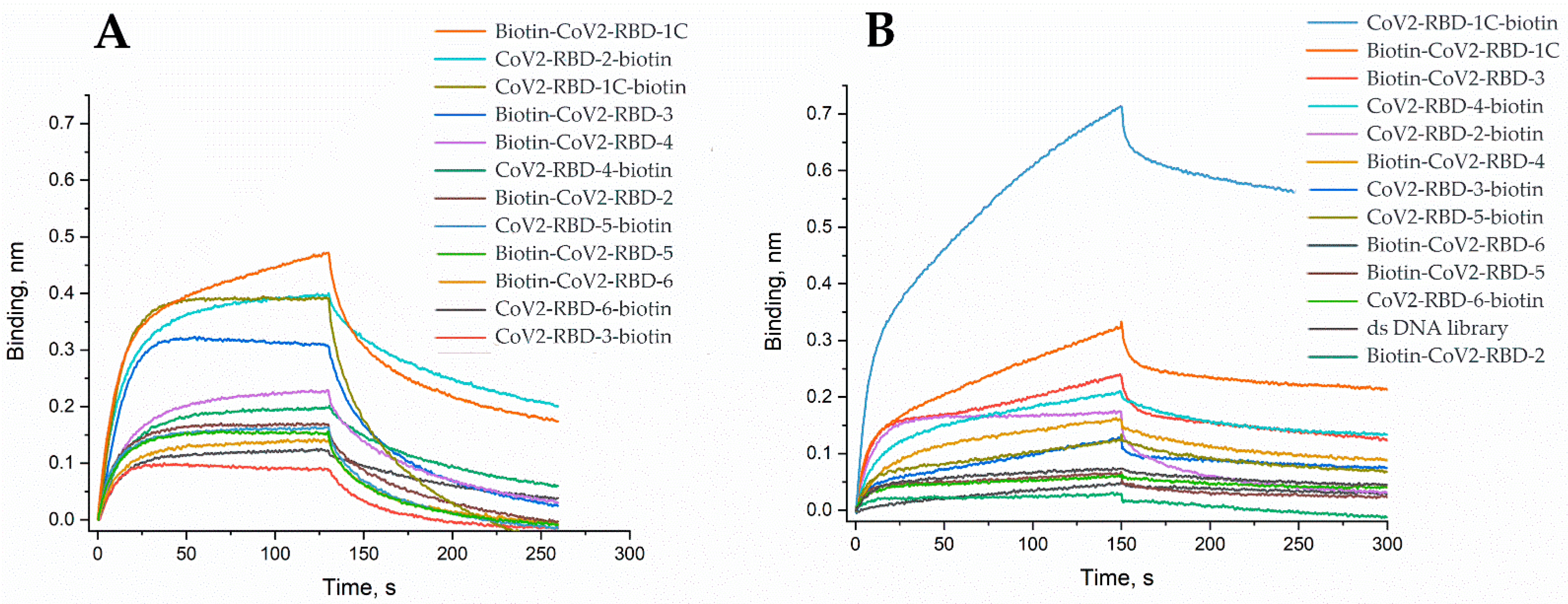
2. Results and Discussion
3. Materials and Methods
3.1. Production of a Recombinant RBD in P. pastoris and Purification
3.2. SDS-PAGE and Western Blot Analysis
3.3. Oligonucleotides
3.4. Binding Experiments
- Initial baseline carried in binding buffer for 90 s;
- Loading of aptamer for 210 s;
- The second baseline in binding buffer for 30 s;
- Association for 150 s (50, 100 or 200 nM protein solution in binding buffer);
- Dissociation in binding buffer for 150 s;
- Washing step-1 carried in 1 M ethanolamine (pH 8.3) for 120 s;
- Washing step-2 carried in binding buffer for 120 s.
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ranjan, P.; Singha, A.; Yadav, S.; Kumar, N.; Murali, S.; Sanghi, S.K.; Khan, R. Rapid diagnosis of SARS-CoV-2 using potential point-of-care electrochemical immunosensor: Toward the future prospects. Int. Rev. Immunol. 2021, 40, 126–142. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.; Ranjan, P.; Sanghi, S.K.; Srivastava, A.K.; Khan, R. Point-of-care biosensor-based diagnosis of COVID-19 holds promise to combat current and future pandemics. ACS Appl. Bio Mater. 2020, 3, 7326–7343. [Google Scholar] [CrossRef]
- Yadav, S.; Sadique, M.A.; Ranjan, P.; Kumar, N.; Singhal, A.; Srivastava, A.K.; Khan, R. SERS based lateral flow immunoassay for point-of-care detection of SARS-CoV-2 in clinical samples. ACS Appl. Bio Mater. 2021, 4, 2974–2995. [Google Scholar] [CrossRef]
- Arya, R.; Kumari, S.; Pandey, B.; Mistry, H.; Bihani, S.C.; Das, A.; Prashar, V.; Gupta, G.D.; Panicker, L.; Kumar, M. Structural insights into SARS-CoV-2 proteins. J. Mol. Biol. 2021, 433, 166725. [Google Scholar] [CrossRef] [PubMed]
- PANGO Lineages: Latest Epidemiological Lineages of SARS-CoV-2. Available online: Cov-lineages.org (accessed on 20 November 2021).
- WHO. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants (accessed on 20 November 2021).
- Liu, X.; Wang, Y.L.; Wu, J.; Qi, J.; Zeng, Z.; Wan, Q.; Chen, Z.; Manandhar, P.; Cavener, V.S.; Boyle, N.R.; et al. Neutralizing aptamers block S/RBD-ACE2 interactions and prevent host cell infection. Angew. Chem. Int. Ed. Engl. 2021, 60, 10273–10278. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.; Weber, A.; Bayin, M.; Breuers, S.; Fieberg, V.; Famulok, M.; Mayer, G.A. SARS-CoV-2 spike binding DNA aptamer that inhibits pseudovirus infection by an RBD-independent mechanism. Angew. Chem. Int. Ed. 2021, 60, 10279–10285. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Z.; Gu, J.; Stacey, H.D.; Ang, J.C.; Capretta, A.; Filipe, C.D.M.; Mossman, K.L.; Balion, C.; Salena, B.J.; et al. Diverse high-affinity DNA aptamers for wild-type and B.1.1.7 SARS-CoV-2 spike proteins from a pre-structured DNA library. Nucleic Acid Res. 2021, 49, 7267–7279. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, Z.; Mohammed, I.; Zhao, L.; Wei, W.; Xiao, H.; Guo, W.; Zhao, Y.; Qu, F.; Huanget, Y. Identification of SARS-CoV-2-against aptamer with high neutralization activity by blocking the RBD domain of spike protein 1. Sig. Transduct. Target Ther. 2021, 6, 227. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Anand, A.; Jain, N.; Goswami, S.; Anantharaj, A.; Patil, S.; Singh, R.; Kumar, A.; Shrivastava, T.; Bhatnagar, S.; et al. A novel G-quadruplex aptamer-based spike trimeric antigen test for the detection of SARS-CoV-2. Mol. Ther. Nucleic Acids. 2021, 26, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Kacherovsky, N.; Yang, L.F.; Dang, H.V.; Cheng, E.L.; Cardle, I.I.; Walls, A.C.; McCallum, M.; Sellers, D.L.; DiMaio, F.; Salipante, S.J.; et al. Discovery and characterization of spike N-terminal domain-binding aptamers for rapid SARS-CoV-2 detection. Angew. Chem. Int. Ed. Engl. 2021, 60, 21211–21215. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Song, J.; Wei, X.; Huang, M.; Sun, M.; Zhu, L.; Lin, B.; Shen, H.; Zhu, Z.; Yang, C. Discovery of aptamers targeting the receptor-binding domain of the SARS-CoV-2 spike glycoprotein. Anal. Chem. 2020, 92, 9895–9900. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, S.; Wei, X.; Wan, S.; Huang, M.; Song, T.; Lu, Y.; Weng, X.; Lin, Z.; Chen, H.; et al. Aptamer blocking strategy inhibits SARS-CoV-2 virus infection. Angew. Chem. Int. Ed. 2021, 60, 10266. [Google Scholar] [CrossRef] [PubMed]
- Alves Ferreira-Bravo, I.; DeStefano, J.J. Xeno-nucleic acid (XNA) 2’-fluoro-arabino nucleic acid (FANA) aptamers to the receptor-binding domain of SARS-CoV-2 S protein block ACE2 binding. Viruses 2021, 13, 1983. [Google Scholar] [CrossRef] [PubMed]
- Stanborough, T.; Given, F.M.; Koch, B.; Sheen, C.R.; Stowers-Hull, A.B.; Waterland, M.R.; Crittenden, D.L. Optical detection of CoV-SARS-2 viral proteins to sub-picomolar concentrations. ACS Omega 2021, 6, 6404–6413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Pandey, R.; Li, J.; Gu, J.; White, D.; Stacey, H.D.; Ang, J.C.; Steinberg, C.J.; Capretta, A.; Filipe, C.D.M.; et al. High-affinity dimeric aptamers enable the rapid electrochemical detection of wild-type and B.1.1.7 SARS-CoV-2 in unprocessed saliva. Angew. Chem. Int. Ed. Engl. 2021, 60, 24266–24274. [Google Scholar] [CrossRef] [PubMed]
- Zavyalova, E.; Ambartsumyan, O.; Zhdanov, G.; Gribanyov, D.; Gushchin, V.; Tkachuk, A.; Rudakova, E.; Nikiforova, M.; Kuznetsova, N.; Popova, L.; et al. SERS-based aptasensor for rapid quantitative detection of SARS-CoV-2. Nanomaterials 2021, 11, 1394. [Google Scholar] [CrossRef] [PubMed]
- Wildt, S.; Gerngross, T.U. The humanization of N-glycosylation pathways in yeast. Nat. Rev. Microbiol. 2005, 3, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Sinegubova, M.V.; Orlova, N.A.; Kovnir, S.V.; Dayanova, L.K.; Vorobiev, I.I. High-level expression of the monomeric SARS-CoV-2 S protein RBD 320-537 in stably transfected CHO cells by the EEF1A1-based plasmid vector. PLoS ONE 2021, 16, e0242890. [Google Scholar] [CrossRef] [PubMed]
- Cleri, F.; Lensink, M.F.; Blossey, R. DNA aptamers block the receptor binding domain at the spike protein of SARS-CoV-2. Front. Mol. Biosci. 2021, 8, 713003. [Google Scholar] [CrossRef] [PubMed]
- SPR Pages. Available online: www.sprpages.nl (accessed on 28 November 2021).


{kind=link}
{kind=link}
| Aptamer | RBD Expressed in CHO Cell Line | RBD Expressed in P. pastoris | ||
|---|---|---|---|---|
| KD, nM | Max Signal | KD, nM | Max Signal | |
| Biotin-CoV2-RBD-1C | 5.8 ± 1.4 | 0.47 | 0.80 ± 0.16 | 0.32 |
| CoV2-RBD-2-Biotin | 4.3 ± 1.4 | 0.40 | 10.0 ± 1.8 | 0.17 |
| CoV2-RBD-1C-Biotin | 27 ± 5 | 0.39 | 1.5 ± 0.3 | 0.71 |
| Biotin-CoV2-RBD-3 | 40 ± 6 | 0.32 | 1.4 ± 0.4 | 0.23 |
| Biotin-CoV2-RBD-4 | 18 ± 2 | 0.22 | 14 ± 4 | 0.16 |
| CoV2-RBD-4-Biotin | 21 ± 4 | 0.20 | 5.5 ± 0.9 | 0.21 |
| Biotin-CoV2-RBD-2 | 17 ± 5 | 0.17 | n.d. | 0.03 |
| CoV2-RBD-5-Biotin | 26 ± 5 | 0.16 | 1.20 ± 0.11 | 0.12 |
| Biotin-CoV2-RBD-5 | 22 ± 5 | 0.15 | n.d. | 0.06 |
| Biotin-CoV2-RBD-6 | 29 ± 8 | 0.14 | 3.5 ± 0.9 | 0.07 |
| CoV2-RBD-6-Biotin | 10 ± 2 | 0.12 | 1.0 ± 0.3 | 0.06 |
| CoV2-RBD-3-Biotin | 24 ± 3 | 0.10 | 2.0 ± 0.7 | 0.12 |
| Aptamer | RBD Expressed in CHO Cell Line | RBD Expressed in P. pastoris | ||
|---|---|---|---|---|
| kon, µM−1s−1 | koff, ms−1 | kon, µM−1s−1 | koff, ms−1 | |
| Biotin-CoV2-RBD-1C | 1.1 ± 0.3 | 6.3 ± 1.4 | 1.5 ± 0.3 | 1.2 ± 0.4 |
| CoV2-RBD-2-Biotin | 0.8 ± 0.2 | 3.6 ± 1.4 | 1.0 ± 0.2 | 10.0 ± 1.2 |
| CoV2-RBD-1C-Biotin | 0.73 ± 0.14 | 19 ± 3 | 0.9 ± 0.2 | 1.4 ± 0.3 |
| Biotin-CoV2-RBD-3 | 0.51 ± 0.08 | 21 ± 3 | 1.2 ± 0.3 | 1.6 ± 0.5 |
| Biotin-CoV2-RBD-4 | 0.75 ± 0.12 | 13.9 ± 0.9 | 0.59 ± 0.15 | 8 ± 3 |
| CoV2-RBD-4-Biotin | 0.53 ± 0.14 | 11.0 ± 1.6 | 0.80 ± 0.16 | 4.3 ± 0.6 |
| Biotin-CoV2-RBD-2 | 0.8 ± 0.3 | 14.4 ± 0.8 | n.d. | n.d. |
| CoV2-RBD-5-Biotin | 0.8 ± 0.2 | 20.8 ± 0.9 | 1.62 ± 0.09 | 1.9 ± 0.2 |
| Biotin-CoV2-RBD-5 | 1.1 ± 0.3 | 24 ± 3 | n.d. | n.d. |
| Biotin-CoV2-RBD-6 | 0.8 ± 0.2 | 23.4 ± 0.8 | 1.7 ± 0.4 | 5.8 ± 1.6 |
| CoV2-RBD-6-Biotin | 0.74 ± 0.13 | 7.7 ± 1.7 | 2.0 ± 0.5 | 2.0 ± 0.8 |
| CoV2-RBD-3-Biotin | 0.98 ± 0.12 | 24 ± 4 | 0.7 ± 0.3 | 1.3 ± 0.3 |
| Aptamer | Sequence |
|---|---|
| Biotin-CoV2-RBD-1C | biotin-5′-T10-CAGCACCGACCTTGTGCTTTGGGAGTGCTGGTCC-AAGGGCGTTAATGGACA-3′ |
| CoV2-RBD-1C-Biotin | 5′-CAGCACCGACCTTGTGCTTTGGGAGTGCTGGTCC-AAGGGCGTTAATGGACA-T10-3′-biotin |
| Biotin-CoV2-RBD-2 | biotin-5′-T10-ATCCAGAGTGACGCAGCATCGAGTGGTGGGCTGGTC-GGGTTTGGATTCCCTTAGATGCTGGACACGGTGGCTTAGT-3′ |
| CoV2-RBD-2-Biotin | 5′-ATCCAGAGTGACGCAGCATCGAGTGGTGGGCTGGTC-GGGTTTGGATTCCCTTAGATGCTGGACACGGTGGCTTAGT-T10-3′-biotin |
| Biotin-CoV2-RBD-3 | biotin-5′-T10-ATCCAGAGTGACGCAGCACTGCGTAGGCGCGGCCAAT-GTGTAGGATTGCTCAGGTCTGCTGGACACGGTGGCTTAGT-3′ |
| CoV2-RBD-3-Biotin | 5′-ATCCAGAGTGACGCAGCACTGCGTAGGCGCGGCCAAT-GTGTAGGATTGCTCAGGTCTGCTGGACACGGTGGCTTAGT-T10-3′-biotin |
| Biotin-CoV2-RBD-4 | biotin-5′-T10-ATCCAGAGTGACGCAGCATTTCATCGGGTCCAAAA-GGGGCTGCTCGGGATTGCGGATATGGACACGT-3′ |
| CoV2-RBD-4-Biotin | 5′-ATCCAGAGTGACGCAGCATTTCATCGGGTCCAAAA-GGGGCTGCTCGGGATTGCGGATATGGACACGT-T10-3′-biotin |
| Biotin-CoV2-RBD-5 | biotin-5′-T10-ATCCAGAGTGACGCAGCAGGACTGCTTAGGATTGCGAAGCTGAGGAGCTCCCCCGCCTTGGACACGGTGGCTTAGT-3′ |
| CoV2-RBD-5-Biotin | 5′-ATCCAGAGTGACGCAGCAGGACTGCTTAGGATTGCGAA-GCTGAGGAGCTCCCCCGCCTTGGACACGGTGGCTTAGT-T10-3′-biotin |
| Biotin-CoV2-RBD-6 | biotin-5′-T10-ATCCAGAGTGACGCAGCAGTAGGGGGATTGGCTCCAGGG-CCTGGCTGACGGTTGCACGTGGACACGGTGGCTTAGT-3′ |
| CoV2-RBD-6-Biotin | 5′-ATCCAGAGTGACGCAGCAGTAGGGGGATTGGCTCCAGGG-CCTGGCTGACGGTTGCACGTGGACACGGTGGCTTAGT-T10-3′-biotin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabovenko, F.; Nikiforova, L.; Yanenko, B.; Ulitin, A.; Loktyushov, E.; Zatsepin, T.; Zavyalova, E.; Zvereva, M. Glycosylation of Receptor Binding Domain of SARS-CoV-2 S-Protein Influences on Binding to Immobilized DNA Aptamers. Int. J. Mol. Sci. 2022, 23, 557. https://doi.org/10.3390/ijms23010557
Grabovenko F, Nikiforova L, Yanenko B, Ulitin A, Loktyushov E, Zatsepin T, Zavyalova E, Zvereva M. Glycosylation of Receptor Binding Domain of SARS-CoV-2 S-Protein Influences on Binding to Immobilized DNA Aptamers. International Journal of Molecular Sciences. 2022; 23(1):557. https://doi.org/10.3390/ijms23010557
Chicago/Turabian StyleGrabovenko, Fedor, Liudmila Nikiforova, Bogdan Yanenko, Andrey Ulitin, Eugene Loktyushov, Timofei Zatsepin, Elena Zavyalova, and Maria Zvereva. 2022. "Glycosylation of Receptor Binding Domain of SARS-CoV-2 S-Protein Influences on Binding to Immobilized DNA Aptamers" International Journal of Molecular Sciences 23, no. 1: 557. https://doi.org/10.3390/ijms23010557