
Antithrombin and Its Role in Host Defense and Inflammation

{kind=link}
Abstract
:1. Overview
2. The Role of AT in Host Response to Infection
3. AT and Inflammation
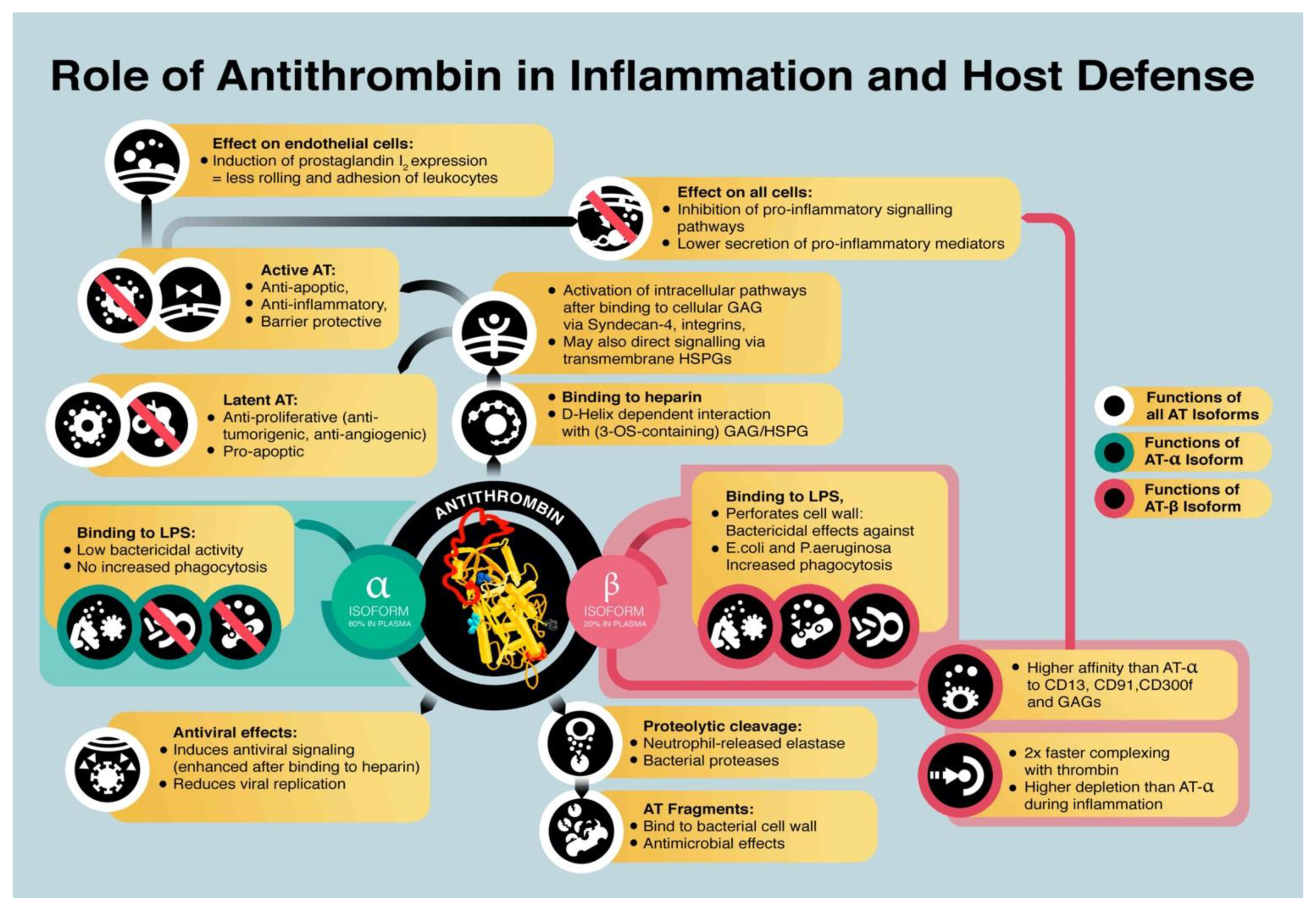
- Binding of AT to heparan-sulfate proteoglycans transmits anti-inflammatory effects. Binding to endothelial heparin-like GAGs induced the release of endothelial prostaglandin I2 [61], which inhibits platelet activation and suppresses adhesion and rolling of leukocytes on endothelial cells [62]. AT interacts with the glycosaminoglycan syndecan-4 via its heparin-binding domain [63] and interacts with receptors of the integrin family [64] on the cell surface [65].
- AT directly binds to specific cellular receptors that inhibit a pro-inflammatory response. AT was shown to interact with LRP-1, CD13, and CD300f, which reduces the pro-inflammatory response by suppressing the expression of IL-6, TNF-α, and tissue factor, e.g., by inhibiting formation of the nuclear factor кB (NF-кB) complex [29,30,38,66].
4. Role of Antithrombin in Tissue Damage
5. Antithrombin as a Key Target for Sepsis Treatment
6. Immunologic Implications of Therapeutic Antithrombin Supplementation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Agostini, A.I.; Watkins, S.C.; Slayter, H.S.; Youssoufian, H.; Rosenberg, R.D. Localization of anticoagulantly active heparan sulfate proteoglycans in vascular endothelium: Antithrombin binding on cultured endothelial cells and perfused rat aorta. J. Cell Biol. 1990, 111, 1293–1304. [Google Scholar] [CrossRef]
- Guo, C.; Fan, X.; Qiu, H.; Xiao, W.; Wang, L.; Xu, B. High-resolution probing heparan sulfate-antithrombin interaction on a single endothelial cell surface: Single-molecule AFM studies. Phys. Chem. Chem. Phys. 2015, 17, 13301–13306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohdan, N.; Espin, S.; Aguila, S.; Teruel-Montoya, R.; Vicente, V.; Corral, J.; Martinez-Martinez, I. Heparanase activates antithrombin through the binding to its heparin binding site. PLoS ONE 2016, 11, e0157834. [Google Scholar] [CrossRef] [PubMed]
- Marcum, J.A.; McKenney, J.B.; Rosenberg, R.D. Acceleration of thrombin-antithrombin complex formation in rat hindquarters via heparinlike molecules bound to the endothelium. J. Clin. Investig. 1984, 74, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Westmark, P.R.; Tanratana, P.; Sheehan, J.P. Selective disruption of heparin and antithrombin-mediated regulation of human factor IX. J. Thromb. Haemost. 2015, 13, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Joist, J.H.; Cowan, J.F.; Khan, M. Rapid loss of factor XII and XI activity in ellagic acid-activated normal plasma: Role of plasma inhibitors and implications for automated activated partial thromboplastin time recording. J. Lab. Clin. Med. 1977, 90, 1054–1065. [Google Scholar]
- Derkx, F.H.; Bouma, B.N.; Schalekamp, M.A. Prorenin-renin conversion by the contact activation system in human plasma: Role of plasma protease inhibitors. J. Lab. Clin. Med. 1984, 103, 560–573. [Google Scholar] [PubMed]
- Vatsyayan, R.; Kothari, H.; Mackman, N.; Pendurthi, U.R.; Rao, L.V. Inactivation of factor VIIa by antithrombin in vitro, ex vivo and in vivo: Role of tissue factor and endothelial cell protein C receptor. PLoS ONE 2014, 9, e103505. [Google Scholar] [CrossRef]
- Rezaie, A.R.; Giri, H. Anticoagulant and signaling functions of antithrombin. J. Thromb. Haemost. 2020, 18, 3142–3153. [Google Scholar] [CrossRef]
- Reda, S.; Müller, J.; Pavlova, A.; Pezeshkpoor, B.; Oldenburg, J.; Pötzsch, B.; Rühl, H. Functional Characterization of Antithrombin Mutations by Monitoring of Thrombin Inhibition Kinetics. Int. J. Mol. Sci. 2021, 22, 2119. [Google Scholar] [CrossRef]
- Gindele, R.; Selmeczi, A.; Oláh, Z.; Ilonczai, P.; Pfliegler, G.; Marján, E.; Nemes, L.; Nagy, Á.; Losonczy, H.; Mitic, G.; et al. Clinical and laboratory characteristics of antithrombin deficiencies: A large cohort study from a single diagnostic center. Thromb. Res. 2017, 160, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Bereczky, Z.; Gindele, R.; Fiatal, S.; Speker, M.; Miklós, T.; Balogh, L.; Mezei, Z.; Szabó, Z.; Ádány, R. Age and origin of the founder antithrombin Budapest 3 (p.Leu131Phe) mutation; its high prevalence in the Roma population and its association with cardiovascular diseases. Front. Cardiovasc. Med. 2021, 7, 617711. [Google Scholar] [CrossRef] [PubMed]
- Niederwanger, C.; Hell, T.; Hofer, S.; Salvador, C.; Michel, M.; Schenk, B.; Treml, B.; Bachler, M. Antithrombin deficiency is associated with mortality and impaired organ function in septic pediatric patients: A retrospective study. PeerJ 2018, 6, e5538. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhang, Y.; Tian, R.; Jin, W.; Du, J.; Zhou, Z.; Wang, R. A prediction model of sepsis-associated acute kidney injury based on antithrombin III. Clin. Exp. Med. 2020, 21. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Klok, F.A.; Kruip, M.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef]
- Lodigiani, C.; Iapichino, G.; Carenzo, L.; Cecconi, M.; Ferrazzi, P.; Sebastian, T.; Kucher, N.; Studt, J.D.; Sacco, C.; Bertuzzi, A.; et al. Venous and arterial thromboembolic complications in COVID-19 patients admitted to an academic hospital in Milan, Italy. Thromb. Res. 2020, 191, 9–14. [Google Scholar] [CrossRef]
- Carsana, L.; Sonzogni, A.; Nasr, A.; Rossi, R.S.; Pellegrinelli, A.; Zerbi, P.; Rech, R.; Colombo, R.; Antinori, S.; Corbellino, M.; et al. Pulmonary post-mortem findings in a series of COVID-19 cases from northern Italy: A two-centre descriptive study. Lancet Infect. Dis. 2020, 20, 1135–1140. [Google Scholar] [CrossRef]
- Panigada, M.; Bottino, N.; Tagliabue, P.; Grasselli, G.; Novembrino, C.; Chantarangkul, V.; Pesenti, A.; Peyvandi, F.; Tripodi, A. Hypercoagulability of COVID-19 patients in intensive care unit: A report of thromboelastography findings and other parameters of hemostasis. J. Thromb. Haemost. 2020, 18, 1738–1742. [Google Scholar] [CrossRef]
- Gazzaruso, C.; Paolozzi, E.; Valenti, C.; Brocchetta, M.; Naldani, D.; Grignani, C.; Salvucci, F.; Marino, F.; Coppola, A.; Gallotti, P. Association between antithrombin and mortality in patients with COVID-19. A possible link with obesity. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1914–1919. [Google Scholar] [CrossRef]
- Briguglio, M.; Crespi, T.; Pino, F.; Mazzocchi, M.; Porta, M.; De Vecchi, E.; Banfi, G.; Perazzo, P. Clinical characteristics of severe COVID-19 patients admitted to an Intensive Care Unit in Lombardy during the Italian pandemic. Front. Med. 2021. [Google Scholar] [CrossRef]
- Levy, J.H.; Sniecinski, R.M.; Welsby, I.J.; Levi, M. Antithrombin: Anti-inflammatory properties and clinical applications. Throm. Haemost. 2016, 115, 712–728. [Google Scholar] [CrossRef]
- Hjorleifsson, E.; Sigurdsson, M.I.; Gudmundsdottir, B.R.; Sigurdsson, G.H.; Onundarson, P.T. Prediction of survival in patients suspected of disseminated intravascular coagulation. Acta Anaesthesiol. Scand. 2015, 59, 870–880. [Google Scholar] [CrossRef]
- Levi, M.; van der Poll, T.; Büller, H.R. Bidirectional relation between inflammation and coagulation. Circulation 2004, 109, 2698–2704. [Google Scholar] [CrossRef] [Green Version]
- Rezaie, A.R.; Giri, H. Antithrombin: An anticoagulant, anti-inflammatory and antibacterial serpin. J. Thromb. Haemost. 2020, 18, 528–533. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Pei, X.Y.; Skinner, R.; Abrahams, J.P.; Carrell, R.W. Structure of beta-antithrombin and the effect of glycosylation on antithrombin’s heparin affinity and activity. J. Mol. Biol. 2003, 326, 823–833. [Google Scholar] [CrossRef]
- Kamp, P.; Strathmann, A.; Ragg, H. Heparin cofactor II, antithrombin-beta and their complexes with thrombin in human tissues. Thromb. Res. 2001, 101, 483–491. [Google Scholar] [CrossRef]
- Chan, A.K.; Berry, L.R.; Paredes, N.; Parmar, N. Isoform composition of antithrombin in a covalent antithrombin-heparin complex. Biochem. Biophys. Res. Commun. 2003, 309, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Romisch, J.; Donges, R.; Stauss, H.; Inthorn, D.; Muhlbayer, D.; Jochum, M.; Hoffmann, J. Quantification of antithrombin isoform proportions in plasma samples of healthy subjects, sepsis patients, and in antithrombin concentrates. Pathophysiol. Haemost. Thromb. 2002, 32, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Oelschlager, C.; Romisch, J.; Staubitz, A.; Stauss, H.; Leithauser, B.; Tillmanns, H.; Holschermann, H. Antithrombin III inhibits nuclear factor kappaB activation in human monocytes and vascular endothelial cells. Blood 2002, 99, 4015–4020. [Google Scholar] [CrossRef] [Green Version]
- Duensing, T.D.; Wing, J.S.; van Putten, J.P. Sulfated polysaccharide-directed recruitment of mammalian host proteins: A novel strategy in microbial pathogenesis. Infect. Immun. 1999, 67, 4463–4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehio, M.; Gomez-Duarte, O.G.; Dehio, C.; Meyer, T.F. Vitronectin-dependent invasion of epithelial cells by Neisseria gonorrhoeae involves alpha(v) integrin receptors. FEBS Lett. 1998, 424, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Duensing, T.D.; Putten, J.P. Vitronectin binds to the gonococcal adhesin OpaA through a glycosaminoglycan molecular bridge. Biochem. J. 1998, 334, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallstrom, T.; Singh, B.; Kraiczy, P.; Hammerschmidt, S.; Skerka, C.; Zipfel, P.F.; Riesbeck, K. Conserved Patterns of Microbial Immune Escape: Pathogenic Microbes of Diverse Origin Target the Human Terminal Complement Inhibitor Vitronectin via a Single Common Motif. PLoS ONE 2016, 11, e0147709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widjaja, M.; Berry, I.J.; Jarocki, V.M.; Padula, M.P.; Dumke, R.; Djordjevic, S.P. Cell surface processing of the P1 adhesin of Mycoplasma pneumoniae identifies novel domains that bind host molecules. Sci. Rep. 2020, 10, 6384. [Google Scholar] [CrossRef]
- Lerch, T.F.; Chapman, M.S. Identification of the heparin binding site on adeno-associated virus serotype 3B (AAV-3B). Virology 2012, 423, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Minnema, M.C.; Chang, A.C.; Jansen, P.M.; Lubbers, Y.T.; Pratt, B.M.; Whittaker, B.G.; Taylor, F.B.; Hack, C.E.; Friedman, B. Recombinant human antithrombin III improves survival and attenuates inflammatory responses in baboons lethally challenged with Escherichia coli. Blood 2000, 95, 1117–1123.043A. [Google Scholar] [CrossRef]
- Papareddy, P.; Rossnagel, M.; Doreen Hollwedel, F.; Kilic, G.; Veerla, S.; Naudin, C.; Smeds, E.; Westman, J.; Martinez-Martinez, I.; Egesten, A.; et al. A human antithrombin isoform dampens inflammatory responses and protects from organ damage during bacterial infection. Nat. Microbiol. 2019, 4, 2442–2455. [Google Scholar] [CrossRef]
- Lawrence, D.A.; Olson, S.T.; Muhammad, S.; Day, D.E.; Kvassman, J.O.; Ginsburg, D.; Shore, J.D. Partitioning of serpin-proteinase reactions between stable inhibition and substrate cleavage is regulated by the rate of serpin reactive center loop insertion into beta-sheet A. J. Biol. Chem. 2000, 275, 5839–5844. [Google Scholar] [CrossRef] [Green Version]
- Papareddy, P.; Kalle, M.; Bhongir, R.K.; Mörgelin, M.; Malmsten, M.; Schmidtchen, A. Antimicrobial effects of helix D-derived peptides of human antithrombin III. J. Biol. Chem. 2014, 289, 29790–29800. [Google Scholar] [CrossRef] [Green Version]
- Hofstra, J.J.; Cornet, A.D.; de Rooy, B.F.; Vlaar, A.P.; van der Poll, T.; Levi, M.; Zaat, S.A.; Schultz, M.J. Nebulized antithrombin limits bacterial outgrowth and lung injury in Streptococcus pneumoniae pneumonia in rats. Crit. Care 2009, 13, R145. [Google Scholar] [CrossRef] [Green Version]
- Dinarvand, P.; Yang, L.; Biswas, I.; Giri, H.; Rezaie, A.R. Plasmodium falciparum histidine rich protein HRPII inhibits the anti-inflammatory function of antithrombin. J. Thromb. Haemost. 2020, 18, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Quenelle, D.C.; Hartman, T.L.; Buckheit, R.W.; Prichard, M.N.; Lynn, R.G. Anti-HSV activity of serpin antithrombin III. Int. Trends Immun. 2014, 2, 87–92. [Google Scholar]
- Geiben-Lynn, R.; Brown, N.; Walker, B.D.; Luster, A.D. Purification of a modified form of bovine antithrombin III as an HIV-1 CD8+ T-cell antiviral factor. J. Biol. Chem. 2002, 277, 42352–42357. [Google Scholar] [CrossRef] [Green Version]
- Smee, D.F.; Hurst, B.L.; Day, C.W.; Geiben-Lynn, R. Influenza Virus H1N1 inhibition by serine protease inhibitor (serpin) antithrombin III. Int. Trends Immun. 2014, 2, 83–86. [Google Scholar]
- Asmal, M.; Seaman, M.; Lin, W.; Chung, R.T.; Letvin, N.L.; Geiben-Lynn, R. Inhibition of HCV by the serpin antithrombin III. Virol. J. 2012, 9, 226. [Google Scholar] [CrossRef] [Green Version]
- Whitney, J.B.; Asmal, M.; Geiben-Lynn, R. Serpin induced antiviral activity of prostaglandin synthetase-2 against HIV-1 replication. PLoS ONE 2011, 6, e18589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmaleh, D.R.; Brown, N.V.; Geiben-Lynn, R. Anti-viral activity of human antithrombin III. Int. J. Mol. Med. 2005, 16, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Asmal, M.; Whitney, J.B.; Luedemann, C.; Carville, A.; Steen, R.; Letvin, N.L.; Geiben-Lynn, R. In vivo anti-HIV activity of the heparin-activated serine protease inhibitor antithrombin III encapsulated in lymph-targeting immunoliposomes. PLoS ONE 2012, 7, e48234. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, E.M.; Linhardt, R.J. Heparin-binding domains in vascular biology. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1549–1557. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, H.; Masuda, Y. Comparison of Protective Effects of Recombinant Antithrombin Gamma and Plasma-Derived Antithrombin on Sepsis-Induced Disseminated Intravascular Coagulation and Multiple Organ Failure. Clin. Appl. Thromb. 2020, 26. [Google Scholar] [CrossRef]
- Inthorn, D.; Hoffmann, J.N.; Hartl, W.H.; Muhlbayer, D.; Jochum, M. Effect of antithrombin III supplementation on inflammatory response in patients with severe sepsis. Shock 1998, 10, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Kellner, P.; Nestler, F.; Leimert, A.; Bucher, M.; Czeslick, E.; Sablotzki, A.; Raspe, C. Antithrombin III, but not C1 esterase inhibitor reduces inflammatory response in lipopolysaccharide-stimulated human monocytes in an ex-vivo whole blood setting. Cytokine 2014, 70, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Komura, H.; Uchiba, M.; Mizuochi, Y.; Arai, M.; Harada, N.; Katsuya, H.; Okajima, K. Antithrombin inhibits lipopolysaccharide-induced tumor necrosis factor-alpha production by monocytes in vitro through inhibition of Egr-1 expression. J. Thromb. Haemost. 2008, 6, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Gritti, D.; Malinverno, A.; Gasparetto, C.; Wiedermann, C.J.; Ricevuti, R. Attenuation of leukocyte beta 2-integrin expression by antithrombin-III. Int. J. Immunopathol. Pharmacol. 2004, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Dinarvand, P.; Qureshi, S.H.; Rezaie, A.R. Engineering D-helix of antithrombin in alpha-1-proteinase inhibitor confers antiinflammatory properties on the chimeric serpin. Thromb. Haemost. 2014, 112, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Wildhagen, K.C.; Wiewel, M.A.; Schultz, M.J.; Horn, J.; Schrijver, R.; Reutelingsperger, C.P.; van der Poll, T.; Nicolaes, G.A. Extracellular histone H3 levels are inversely correlated with antithrombin levels and platelet counts and are associated with mortality in sepsis patients. Thromb. Res. 2015, 136, 542–547. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Caudrillier, A.; Kessenbrock, K.; Gilliss, B.M.; Nguyen, J.X.; Marques, M.B.; Monestier, M.; Toy, P.; Werb, Z.; Looney, M.R. Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury. J. Clin. Investig. 2012, 122, 2661–2671. [Google Scholar] [CrossRef]
- Taylor, F.B., Jr.; Emerson, T.E., Jr.; Jordan, R.; Chang, A.K.; Blick, K.E. Antithrombin-III prevents the lethal effects of Escherichia coli infusion in baboons. Circ. Shock 1988, 26, 227–235. [Google Scholar]
- Horie, S.; Ishii, H.; Kazama, M. Heparin-like glycosaminoglycan is a receptor for antithrombin III-dependent but not for thrombin-dependent prostacyclin production in human endothelial cells. Thromb. Res. 1990, 59, 895–904. [Google Scholar] [CrossRef]
- Hoffmann, J.N.; Vollmar, B.; Inthorn, D.; Schildberg, F.W.; Menger, M.D. Antithrombin reduces leukocyte adhesion during chronic endotoxemia by modulation of the cyclooxygenase pathway. Am. J. Physiol. Physiol. 2000, 279. [Google Scholar] [CrossRef]
- Kaneider, N.C.; Egger, P.; Dunzendorfer, S.; Wiedermann, C.J. Syndecan-4 as antithrombin receptor of human neutrophils. Biochem. Biophys. Res. Commun. 2001, 287, 42–46. [Google Scholar] [CrossRef]
- Morgan, M.R.; Humphries, M.J.; Bass, M.D. Synergistic control of cell adhesion by integrins and syndecans. Nat. Rev. Mol. Cell Biol. 2007, 8, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Dunzendorfer, S.; Kaneider, N.; Rabensteiner, A.; Meierhofer, C.; Reinisch, C.; Römisch, J.; Wiedermann, C.J. Cell-surface heparan sulfate proteoglycan-mediated regulation of human neutrophil migration by the serpin antithrombin III. Blood 2001, 97, 1079–1085. [Google Scholar] [CrossRef]
- Kaneider, N.C.; Reinisch, C.M.; Dunzendorfer, S.; Romisch, J.; Wiedermann, C.J. Syndecan-4 mediates antithrombin-induced chemotaxis of human peripheral blood lymphocytes and monocytes. J. Cell Sci. 2002, 115, 227–236. [Google Scholar] [PubMed]
- Schreuder, H.A.; de Boer, B.; Dijkema, R.; Mulders, J.; Theunissen, H.J.; Grootenhuis, P.D.; Hol, W.G. The intact and cleaved human antithrombin III complex as a model for serpin-proteinase interactions. Nat. Struct. Biol. 1994, 1, 48–54. [Google Scholar] [CrossRef]
- Olson, S.T.; Richard, B.; Izaguirre, G.; Schedin-Weiss, S.; Gettins, P.G. Molecular mechanisms of antithrombin-heparin regulation of blood clotting proteinases. A paradigm for understanding proteinase regulation by serpin family protein proteinase inhibitors. Biochimie 2010, 92, 1587–1596. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Sakai, T.; Fassler, R.; Ruoslahti, E. Antiangiogenic proteins require plasma fibronectin or vitronectin for in vivo activity. Proc. Natl. Acad. Sci. USA 2003, 100, 11435–11438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, M.S.; Pirie-Shepherd, S.; Lane, W.S.; Folkman, J. Antiangiogenic activity of the cleaved conformation of the serpin antithrombin. Science 1999, 285, 1926–1928. [Google Scholar] [CrossRef]
- Guerrero, J.A.; Teruel, R.; Martinez, C.; Arcas, I.; Martinez-Martinez, I.; de la Morena-Barrio, M.E.; Vicente, V.; Corral, J. Protective role of antithrombin in mouse models of liver injury. J. Hepatol. 2012, 57, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Wiedermann, C.J. Clinical review: Molecular mechanisms underlying the role of antithrombin in sepsis. Crit. Care 2006, 10, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, K.; Kojima, T.; Kadomatsu, K.; Nakayama, Y.; Takagi, A.; Suzuki, M.; Takeda, N.; Ito, M.; Yamamoto, K.; Matsushita, T.; et al. Complete antithrombin deficiency in mice results in embryonic lethality. J. Clin. Investig. 2000, 106, 873–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, Y.; Wang, J.; Gao, J.; Tong, C.; Manithody, C.; Li, J.; Rezaie, A.R. Antithrombin is protective against myocardial ischemia and reperfusion injury. J. Thromb. Haemost. 2013, 11, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Okajima, K.; Kushimoto, S.; Isobe, H.; Tanaka, K. Antithrombin reduces ischemia/reperfusion injury of rat liver by increasing the hepatic level of prostacyclin. Blood 1999, 93, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Chappell, D.; Jacob, M.; Hofmann-Kiefer, K.; Rehm, M.; Welsch, U.; Conzen, P.; Becker, B.F. Antithrombin reduces shedding of the endothelial glycocalyx following ischaemia/reperfusion. Cardiovasc. Res. 2009, 83, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.F.; Jacob, M.; Leipert, S.; Salmon, A.H.; Chappell, D. Degradation of the endothelial glycocalyx in clinical settings: Searching for the sheddases. Br. J. Clin. Pharmacol. 2015, 80, 389–402. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Aihara, K.; Kadota, K.; Tanaka, H.; Sato, K.; Nagaoka, I. Newly Developed Recombinant Antithrombin Protects the Endothelial Glycocalyx in an Endotoxin-Induced Rat Model of Sepsis. Int. J. Mol. Sci. 2020, 176. [Google Scholar] [CrossRef]
- Camprubi-Rimblas, M.; Tantinya, N.; Guillamat-Prats, R.; Bringue, J.; Puig, F.; Gomez, M.N.; Blanch, L.; Artigas, A. Effects of nebulized antithrombin and heparin on inflammatory and coagulation alterations in an acute lung injury model in rats. J. Thromb. Haemost. 2020, 18, 571–583. [Google Scholar] [CrossRef]
- Matsubara, T.; Yamakawa, K.; Umemura, Y.; Gando, S.; Ogura, H.; Shiraishi, A.; Kushimoto, S.; Abe, T.; Tarui, T.; Hagiwara, A.; et al. Significance of plasma fibrinogen level and antithrombin activity in sepsis: A multicenter cohort study using a cubic spline model. Thromb. Res. 2019, 181, 17–23. [Google Scholar] [CrossRef]
- Lisman, T.; Arefaine, B.; Adelmeijer, J.; Zamalloa, A.; Corcoran, E.; Smith, J.G.; Bernal, W.; Patel, V.C. Global hemostatic status in patients with acute-on-chronic liver failure and septics without underlying liver disease. J. Thromb. Haemost. 2021, 19, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Yamashita, H.; Oka, N.; Ueda, T.; Kohama, K.; Nakao, A.; Kotani, J. Antithrombin III improved neutrophil extracellular traps in lung after the onset of endotoxemia. J. Surg. Res. 2017, 208, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, N.; Goto, K.; Ohchi, Y.; Abe, T.; Koga, H.; Kitano, T. The efficacy and safety of antithrombin and recombinant human thrombomodulin combination therapy in patients with severe sepsis and disseminated intravascular coagulation. J. Crit. Care 2016, 36, 29–34. [Google Scholar] [CrossRef]
- Gando, S.; Saitoh, D.; Ishikura, H.; Ueyama, M.; Otomo, Y.; Oda, S.; Kushimoto, S.; Tanjoh, K.; Mayumi, T.; Ikeda, T.; et al. A randomized, controlled, multicenter trial of the effects of antithrombin on disseminated intravascular coagulation in patients with sepsis. Crit. Care 2013, 17, R297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, T.; Gando, S.; Saitoh, D.; Wada, H.; Di Nisio, M.; Thachil, J. Antithrombin supplementation and risk of bleeding in patients with sepsis-associated disseminated intravascular coagulation. Thromb. Res. 2016, 145, 46–50. [Google Scholar] [CrossRef]
- Tagami, T.; Matsui, H.; Fushimi, K.; Yasunaga, H. Supplemental dose of antithrombin use in disseminated intravascular coagulation patients after abdominal sepsis. Thromb. Haemost. 2015, 114, 537–545. [Google Scholar] [CrossRef]
- Iba, T.; Saitoh, D.; Gando, S.; Thachil, J. The usefulness of antithrombin activity monitoring during antithrombin supplementation in patients with sepsis-associated disseminated intravascular coagulation. Thromb. Res. 2015, 135, 897–901. [Google Scholar] [CrossRef]
- Warren, B.L.; Eid, A.; Singer, P.; Pillay, S.S.; Carl, P.; Novak, I.; Chalupa, P.; Atherstone, A.; Penzes, I.; Kubler, A.; et al. Caring for the critically ill patient. High-dose antithrombin III in severe sepsis: A randomized controlled trial. JAMA 2001, 286, 1869–1878. [Google Scholar] [CrossRef]
- Kienast, J.; Juers, M.; Wiedermann, C.J.; Hoffmann, J.N.; Ostermann, H.; Strauss, R.; Keinecke, H.O.; Warren, B.L.; Opal, S.M. Treatment effects of high-dose antithrombin without concomitant heparin in patients with severe sepsis with or without disseminated intravascular coagulation. J. Thromb. Haemost. 2006, 4, 90–97. [Google Scholar] [CrossRef]
- Allingstrup, M.; Wetterslev, J.; Ravn, F.B.; Moller, A.M.; Afshari, A. Antithrombin III for critically ill patients: A systematic review with meta-analysis and trial sequential analysis. Intensive Care Med. 2016, 42, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Wiedermann, C.J. Antithrombin concentrate use in disseminated intravascular coagulation of sepsis: Meta-analyses revisited. J. Thromb. Haemost. 2018, 16, 455–457. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.C.; Elamin, E.M.; Suffredini, A.F. Inhaled anticoagulation regimens for the treatment of smoke inhalation-associated acute lung injury: A systematic review. Crit. Care Med. 2014, 42, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fourrier, F.; Chopin, C.; Huart, J.J.; Runge, I.; Caron, C.; Goudemand, J. Double-blind, placebo-controlled trial of antithrombin III concentrates in septic shock with disseminated intravascular coagulation. Chest 1993, 104, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Iba, T.; Saito, D.; Wada, H.; Asakura, H. Efficacy and bleeding risk of antithrombin supplementation in septic disseminated intravascular coagulation: A prospective multicenter survey. Thromb. Res. 2012, 130, e129133. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ko, B.S.; Park, S.Y.; Oh, D.K.; Hong, S.B.; Jang, S.; Kim, W.Y. Effect of High-dose Antithrombin Supplementation in Patients with Septic Shock and Disseminated Intravascular Coagulation. Sci. Rep. 2019, 9, 16626. [Google Scholar] [CrossRef]
- Nishida, O.; Ogura, H.; Egi, M.; Fujishima, S.; Hayashi, Y.; Iba, T.; Imaizumi, H.; Inoue, S.; Kakihana, Y.; Kotani, J.; et al. The Japanese Clinical Practice Guidelines for Management of Sepsis and Septic Shock 2016 (J-SSCG 2016). J. Intensive Care 2018, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Furuie, H.; Kanda, H. Randomized Comparison Study of Novel Recombinant Human Antithrombin Gamma and Plasma-Derived Antithrombin in Healthy Volunteers. Clin. Drug Investig. 2019, 39, 1185–1194. [Google Scholar] [CrossRef] [Green Version]
- Tagami, T. Antithrombin concentrate use in sepsis-associated disseminated intravascular coagulation: Re-evaluation of a ’pendulum effect’ drug using a nationwide database. J. Thromb. Haemost. 2018, 16, 458–461. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.N.; Vollmar, B.; Laschke, M.W.; Inthorn, D.; Kaneider, N.C.; Dunzendorfer, S.; Wiedermann, C.J.; Romisch, J.; Schildberg, F.W.; Menger, M.D. Adverse effect of heparin on antithrombin action during endotoxemia: Microhemodynamic and cellular mechanisms. Thromb. Haemost. 2002, 88, 242–252. [Google Scholar]
- Lewis, B.E.; Wallis, D.E.; Berkowitz, S.D.; Matthai, W.H.; Fareed, J.; Walenga, J.M.; Bartholomew, J.; Sham, R.; Lerner, R.G.; Zeigler, Z.R.; et al. Argatroban anticoagulant therapy in patients with heparin-induced thrombocytopenia. Circulation 2001, 103, 1838–1843. [Google Scholar] [CrossRef] [Green Version]
- Hursting, M.J.; Alford, K.L.; Becker, J.C.; Brooks, R.L.; Joffrion, J.L.; Knappenberger, G.D.; Kogan, P.W.; Kogan, T.P.; McKinney, A.A.; Schwarz, R.P., Jr. Novastan (brand of argatroban): A small-molecule, direct thrombin inhibitor. Semin. Thromb. Hemost. 1997, 23, 503–516. [Google Scholar] [CrossRef]
- Beyer, J.; Halbritter, K.; Weise, M.; Schellong, S. Influence of antithrombin and argatroban on disseminated intravascular coagulation parameters in a patient with septic shock. Thromb. Res. 2009, 124, 383–386. [Google Scholar] [CrossRef]
- Jang, I.K.; Brown, D.F.; Giugliano, R.P.; Anderson, H.V.; Losordo, D.; Nicolau, J.C.; Dutra, O.P.; Bazzino, O.; Viamonte, V.M.; Norbady, R.; et al. A multicenter, randomized study of argatroban versus heparin as adjunct to tissue plasminogen activator (TPA) in acute myocardial infarction: Myocardial infarction with novastan and TPA (MINT) study. J. Am. Coll. Cardiol. 1999, 33, 1879–1885. [Google Scholar] [CrossRef] [Green Version]
- Kumon, K.; Tanaka, K.; Nakajima, N.; Naito, Y.; Fujita, T. Anticoagulation with a synthetic thrombin inhibitor after cardiovascular surgery and for treatment of disseminated intravascular coagulation. Crit. Care Med. 1984, 12, 1039–1043. [Google Scholar] [CrossRef]
- Mukundan, S.; Zeigler, Z.R. Direct antithrombin agents ameliorate disseminated intravascular coagulation in suspected heparin-induced thrombocytopenia thrombosis syndrome. Clinical. Appl. Thromb. 2002, 8, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Elg, M.; Gustafsson, D. A combination of a thrombin inhibitor and dexamethasone prevents the development of experimental disseminated intravascular coagulation in rats. Thromb. Res. 2006, 117, 429–437. [Google Scholar] [CrossRef]
- Bachler, M.; Hell, T.; Bosch, J.; Treml, B.; Schenk, B.; Treichl, B.; Friesenecker, B.; Lorenz, I.; Stengg, D.; Hruby, S.; et al. A Prospective Pilot Trial to Assess the Efficacy of Argatroban (Argatra((R))) in Critically Ill Patients with Heparin Resistance. J. Clin. Med. 2020, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nougier, C.; Benoit, R.; Simon, M.; Desmurs-Clavel, H.; Marcotte, G.; Argaud, L.; David, J.S.; Bonnet, A.; Negrier, C.; Dargaud, Y. Hypofibrinolytic state and high thrombin generation may play a major role in SARS-COV2 associated thrombosis. J. Thromb. Haemost. 2020, 18, 2215–2219. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlömmer, C.; Brandtner, A.; Bachler, M. Antithrombin and Its Role in Host Defense and Inflammation. Int. J. Mol. Sci. 2021, 22, 4283. https://doi.org/10.3390/ijms22084283
Schlömmer C, Brandtner A, Bachler M. Antithrombin and Its Role in Host Defense and Inflammation. International Journal of Molecular Sciences. 2021; 22(8):4283. https://doi.org/10.3390/ijms22084283
Chicago/Turabian StyleSchlömmer, Christine, Anna Brandtner, and Mirjam Bachler. 2021. "Antithrombin and Its Role in Host Defense and Inflammation" International Journal of Molecular Sciences 22, no. 8: 4283. https://doi.org/10.3390/ijms22084283