
An Improved Protocol for Agrobacterium-Mediated Transformation in Subterranean Clover (Trifolium subterraneum L.)




Abstract
:1. Introduction
2. Results
2.1. In Vitro Culture Optimisation: Selection of Shooting and Rooting Media
2.2. Antibiotic Sensitivity Testing
2.2.1. Kanamycin Concentration Test
2.2.2. Hygromycin Concentration Test
2.2.3. Cefotaxime Concentration Test
2.2.4. Combined Antibiotics Test
2.3. Cefotaxime Positive Effect on Explant Regeneration
2.4. GUS Plasmid
2.5. GUS Transformation and Genetic Confirmation
3. Discussion
Cefotaxime Positive Effect on Shoot Regeneration
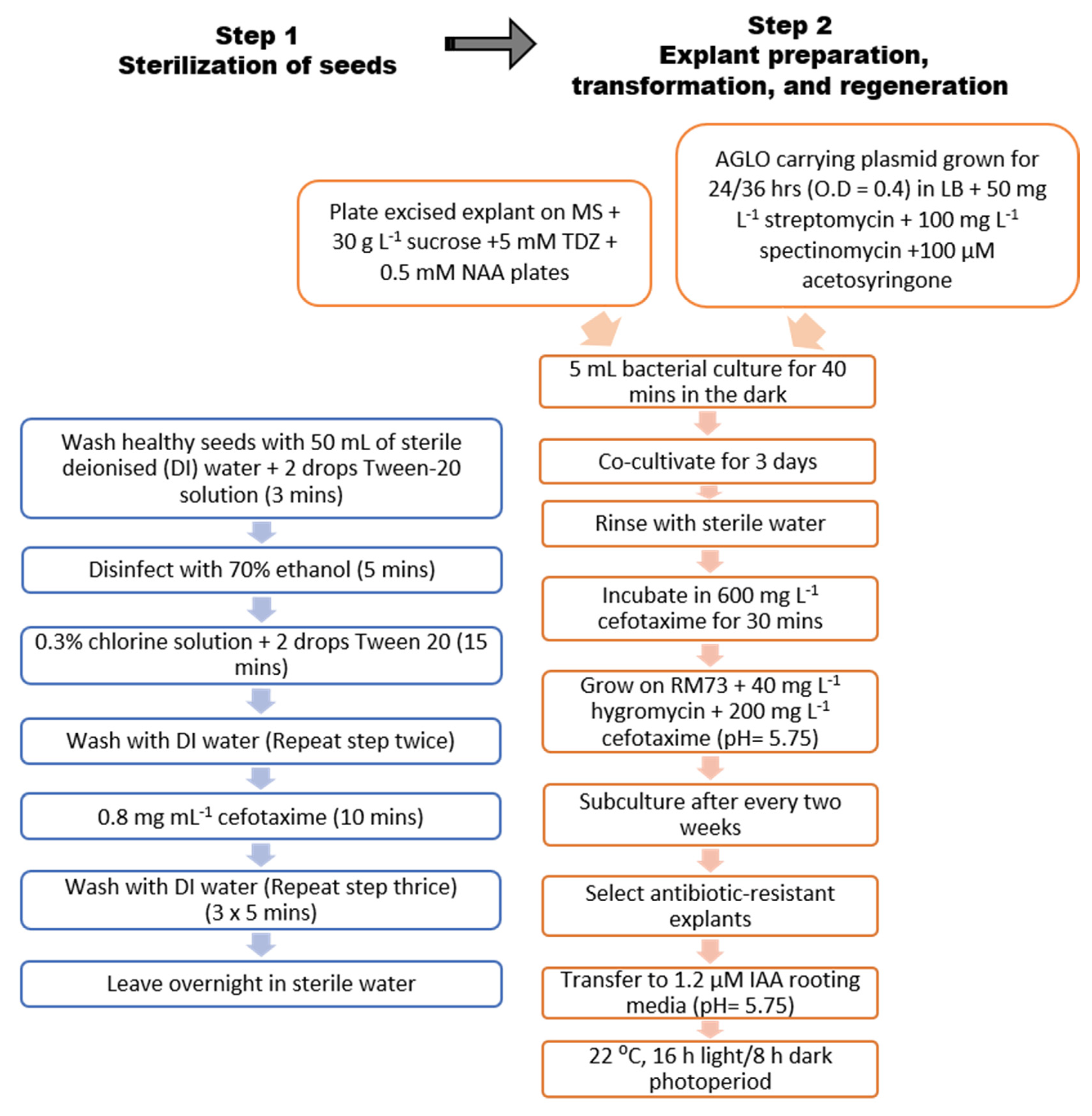
4. Materials and Methods
4.1. Seed Sterilisation
4.2. Explant Preparation and In Vitro Culture
4.3. Antibiotic Concentration Test
4.4. Cefotaxime Effect on Shoot Regeneration
4.5. Plasmids and Bacterial Strains
4.6. In Vitro Plant Transformation and Regeneration
4.7. Genetic Confirmation Test
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nichols, P.G.H.; Revell, C.K.; Humphries, A.W.; Howie, J.H.; Hall, E.J.; Sandral, G.A.; Ghamkhar, K.; Harris, C.A. Temperate pasture legumes in Australia—their history, current use, and future prospects. Crop. Pasture Sci. 2012, 63, 691–725. [Google Scholar] [CrossRef]
- Hirakawa, H.; Kaur, P.; Shirasawa, K.; Nichols, P.; Nagano, S.; Appels, R.; Erskine, W.; Isobe, S.N. Draft genome sequence of subterranean clover, a reference for genus Trifolium. Sci. Rep. 2016, 6, 30358. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Bayer, P.E.; Milec, Z.; Vrána, J.; Yuan, Y.; Appels, R.; Edwards, D.; Batley, J.; Nichols, P.; Erskine, W.; et al. An advanced reference genome of Trifolium subterraneum L. reveals genes related to agronomic performance. Plant Biotechnol. J. 2017, 15, 1034–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, P.G.H.; Foster, K.J.; Piano, E.; Pecetti, L.; Kaur, P.; Ghamkhar, K.; Collins, W.J. Genetic improvement of subterranean clover (Trifolium subterraneum L.). 1. Germplasm, traits and future prospects. Crop. Pasture Sci. 2013, 64, 312–346. [Google Scholar] [CrossRef]
- Peoples, M.B.; Baldock, J.A. Nitrogen dynamics of pastures: Nitrogen fixation inputs, the impact of legumes on soil nitrogen fertility, and the contributions of fixed nitrogen to Australian farming systems. Aust. J. Exp. Agric. 2001, 41, 327. [Google Scholar] [CrossRef]
- Ru, Y.J.; Fortune, J.A. Seed yield and nutritive value of dry, mature subterranean clover (Trifolium subterraneum L.). Aust. J. Exp. Agric. 2001, 41, 169. [Google Scholar] [CrossRef]
- Schut, A.G.T.; Gherardi, S.G.; Wood, D.A. Empirical models to quantify the nutritive characteristics of annual pastures in south-west Western Australia. Crop. Pasture Sci. 2010, 61, 32–43. [Google Scholar] [CrossRef]
- Haling, R.E.; Yang, Z.; Shadwell, N.; Culvenor, R.A.; Stefanski, A.; Ryan, M.H.; Sandral, G.A.; Kidd, D.R.; Lambers, H.; Simpson, R.J. Growth and root dry matter allocation by pasture legumes and a grass with contrasting external critical phosphorus requirements. Plant Soil 2016, 407, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Chu, P.W.G.; Anderson, B.J.; Khan, M.R.I.; Shukla, D.; Higgins, T.J.V. Production of Bean yellow mosaic virus resistant subterranean clover (Trifolium subterraneum) plants by transformation with the virus coat protein gene. Ann. Appl. Biol. 1999, 135, 469–480. [Google Scholar] [CrossRef]
- Ding, Y.-L.; Aldao-Humble, G.; Ludlow, E.; Drayton, M.; Lin, Y.-H.; Nagel, J.; Dupal, M.; Zhao, G.; Pallaghy, C.; Kalla, R.; et al. Efficient plant regeneration and Agrobacterium-mediated transformation in Medicago and Trifolium species. Plant Sci. 2003, 165, 1419–1427. [Google Scholar] [CrossRef]
- Khan, M.; Tabe, L.M.; Heath, L.C.; Spencer, D.; Higgins, T. Agrobacterium-Mediated Transformation of Subterranean Clover (Trifolium subterraneum L.). Plant Physiol. 1994, 105, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Chen, Y.; Schnabel, E.; Crook, A.; Frugoli, J. Comparison of efficiency and time to regeneration of Agrobacterium-mediated transformation methods in Medicago truncatula. Plant Methods 2019, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.F.; Townsend, C.O. A plant-tumor of bacterial origin. Science 1907, 25, 671–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nester, E.W. Agrobacterium: Nature’s genetic engineer. Front. Plant Sci. 2015, 5, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. Rev. 2003, 67, 16–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santana, J.R.F.; Paiva, R.; Aloufa, M.A.I.; De Lemos, E.E.P. Efficiency of ampicillin and benomyl at controlling contamination of Annonaceae leaf segments cultured in vitro. Fruits 2003, 58, 357–361. [Google Scholar] [CrossRef]
- Nguyen, A.H.; Wijayanto, T.; Erskine, W.; Barker, S.J. Using green fluorescent protein sheds light on Lupinus angustifolius L. transgenic shoot development. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 127, 665–674. [Google Scholar] [CrossRef]
- Joersbo, M.; Brunstedt, J.; Marcussen, J.; Okkels, F.T. Transformation of the endospermous legume guar (Cyamopsis tetragonoloba L.) and analysis of transgene transmission. Mol. Breed. 1999, 5, 521–529. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.-B.; Schneeberger, K. The impact of third generation genomic technologies on plant genome assembly. Curr. Opin. Plant Biol. 2017, 36, 64–70. [Google Scholar] [CrossRef]
- Mathias, R.J.; Boyd, L.A. Cefotaxime stimulates callus growth, embryogenesis and regeneration in hexaploid bread wheat (Triticum aestivum L em. Thell). Plant Sci. 1986, 46, 217–223. [Google Scholar] [CrossRef]
- Borrelli, G.; Di Fonzo, N.; Lupotto, E. Effect of Cefotaxime on Callus Culture and Plant Regeneration in Durum Wheat. J. Plant Physiol. 1992, 140, 372–374. [Google Scholar] [CrossRef]
- Danilova, S.A.; Dolgikh, Y.I. The Stimulatory Effect of the Antibiotic Cefotaxime on Plant Regeneration in Maize Tissue Culture. Russ. J. Plant Physiol. 2004, 51, 559–562. [Google Scholar] [CrossRef]
- Heath, L.C.; Chin, S.-F.; Spencer, D.; Higgins, T.J.V. In vitro regeneration of commercial cultivars of subterranean clover. Plant Cell Tissue Organ Cult. (PCTOC) 1993, 35, 43–48. [Google Scholar] [CrossRef]
- Nauerby, B.; Billing, K.; Wyndaele, R. Influence of the antibiotic timentin on plant regeneration compared to carbenicillin and cefotaxime in concentrations suitable for elimination of Agrobacterium tumefaciens. Plant Sci. 1997, 123, 169–177. [Google Scholar] [CrossRef]
- Kaur, P.; Appels, R.; Bayer, P.E.; Keeble-Gagnere, G.; Wang, J.; Hirakawa, H.; Shirasawa, K.; Vercoe, P.; Stefanova, K.; Durmic, Z.; et al. Climate Clever Clovers: New Paradigm to Reduce the Environmental Footprint of Ruminants by Breeding Low Methanogenic Forages Utilizing Haplotype Variation. Front. Plant Sci. 2017, 8, 1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo, F.P.; Nyman, R.K.M.; Johnson, A.A.T.; Navarro, M.P.; Ryan, M.H.; Erskine, W.; Kaur, P. CRISPR-Cas systems: Ushering in the new genome editing era. Bioengineered 2018, 9, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Beattie, L.D.; Garrett, R.G. Adventitious shoot production from immature embryos of white clover. Plant Cell Tissue Organ Cult. (PCTOC) 1995, 42, 67–72. [Google Scholar] [CrossRef]
- Fratini, R.; Ruiz, M.L. A rooting procedure for lentil (Lens culinaris Medik.) and other hypogeous legumes (pea, chickpea and Lathyrus) based on explant polarity. Plant Cell Rep. 2003, 21, 726–732. [Google Scholar] [CrossRef]
| 1 | 0 mg L−1 (Control) |
| 2 | 25 mg L−1 |
| 3 | 100 mg L−1 |
| 4 | 0 mg L−1 (Control) |
| 5 | 10 mg L−1 |
| 6 | 25 mg L−1 |
| 7 | 50 mg L−1 |
| 8 | 0 mg L−1 (Control) |
| 9 | 10 mg L−1 |
| 10 | 25 mg L−1 |
| 11 | 50 mg L−1 |
| 12 | Control (no antibiotics added) |
| 13 | 200 mg L−1 Cef |
| 14 | 100 mg L−1 Kan + 200 mg L−1 Cef |
| 15 | 40 mg L−1 Hyg + 200 mg L−1 Cef |
| 16 | 1 |
| 17 | 128 |
| 18 | 2 |
| 19 | 126 |




{kind=link}
{kind=link}
{kind=link}
| Percentage of Survival | ||||
|---|---|---|---|---|
| Treatment | Number of Explants | 15 Days | 30 Days | 45 Days |
| Kanamycin concentration test | ||||
| 0 mg L−1 (Control) | 45 | 95.5 | 86.6 | 82.2 |
| 25 mg L−1 | 27 | 96.4 | 66.6 | 48.1 |
| 50 mg L−1 | 38 | 76.3 | 60.5 | 44.7 |
| 100 mg L−1 | 30 | 42.8 a | 23.3 a | 16.6 a |
| Hygromycin concentration test | ||||
| 0 mg L−1 (Control) | 30 | 100 | 96.6 | 93.3 |
| 10 mg L−1 | 30 | 90 | 56.4 a | 36.6 a |
| 25 mg L−1 | 30 | 46.6 a | 30 a | 16.6 a |
| 50 mg L−1 | 30 | 16.6 a | 10 a | 0 a |
| Cefotaxime concentration test | ||||
| 0 mg L−1 (Control) | 36 | 81.8 | 72.7 | NA |
| 10 mg L−1 | 33 | 86.1 | 80.5 | NA |
| 25 mg L−1 | 31 | 80.6 | 74.3 | NA |
| 50 mg L−1 | 29 | 82.7 | 79.3 | NA |
| Treatment | Number of Explants | Percentage of Survival after 30 Days |
|---|---|---|
| Control (no antibiotics added) | 29 | 89.6 a |
| 200 mg L−1 Cef | 30 | 80 a |
| 100 mg L−1 Kan | 30 | 13.3 b |
| 100 mg L−1 Kan + 200 mg L−1 Cef | 29 | 27.5 b |
| 40 mg L−1 Hyg + 200 mg L−1 Cef | 45 | 15.5 b |
| Experiment | Total Number of Plants | No. Resistant Explants after 30 Days in Selection Medium (% in Parentheses) | PCR Configuration (% in Parentheses) |
|---|---|---|---|
| 1 | 128 | 23 (17.9%) | 9 (7.0%) |
| 2 | 126 | 22 (17.4%) | 7 (5.5%) |
| 3 | 125 | 26 (20.8%) | 4 (3.2%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojo, F.P.; Seth, S.; Erskine, W.; Kaur, P. An Improved Protocol for Agrobacterium-Mediated Transformation in Subterranean Clover (Trifolium subterraneum L.). Int. J. Mol. Sci. 2021, 22, 4181. https://doi.org/10.3390/ijms22084181
Rojo FP, Seth S, Erskine W, Kaur P. An Improved Protocol for Agrobacterium-Mediated Transformation in Subterranean Clover (Trifolium subterraneum L.). International Journal of Molecular Sciences. 2021; 22(8):4181. https://doi.org/10.3390/ijms22084181
Chicago/Turabian StyleRojo, Fernando Perez, Sumedha Seth, William Erskine, and Parwinder Kaur. 2021. "An Improved Protocol for Agrobacterium-Mediated Transformation in Subterranean Clover (Trifolium subterraneum L.)" International Journal of Molecular Sciences 22, no. 8: 4181. https://doi.org/10.3390/ijms22084181