1. Introduction
Plant nanovesicles are closely related to mammalian extracellular vesicles (EVs) but, compared to the latter, they have been relegated to the background. However, scientific interest has been increasingly paid to the plant equivalent of exosomes and microvesicles lately [
1,
2].
Structure and density of plant nanovesicles are comparable with exosomes from mammals [
3]. Also, multivesicular bodies (MVBs) [
4] and apoptotic bodies [
5] have been observed in plants and even though we found no reports of plasma membrane-derived microvesicles, we assume that the mechanisms of EV formation [
6] can be transferred from the animal kingdom to plants, or even to eukaryotes in general. However, not all of those “exosome-like nanoparticles”, that can be isolated from (homogenized) plant materials like juices, are certainly of extracellular origin. We therefore recommend using the term “nanovesicles” for nanometer-scaled (1–999 nm) membrane-delimited vesicles, as long as the selected raw material does not ensure that EVs are concentrated. For instance, isolated nanovesicles from apoplastic fluid can be assumed to be EVs. The scheme in
Figure 1 provides an overview on present key issues in plant nanovesicle research.
Current research regarding plant nanovesicles deals with general properties [
3,
12,
13,
14], host-versus-pathogen-interactions [
8,
11,
15,
16,
17] and health-beneficial effects [
18,
19,
20,
21,
22,
23,
24]. Another approach to use the advantageous properties of plant nanovesicles can be to foster the use of these small non-coding RNA (sRNA)-containing vehicles as bio-compatible and sustainable plant protection agents [
25]. In order to determine the effects on human health, current preclinical and clinical trials are aiming to prove whether nanovesicles from ginger and aloe improve the condition of polycystic ovary syndrome (NCT03493984), whether grape nanovesicles prevent chemoradiation treatment-associated oral mucositis (NCT01668849), and if plant nanovesicles can effectively deliver curcumin to normal colon tissue and colon tumors (NCT01294072).
Although we have already learned a lot about properties and effects of vesicle subtypes isolated from distinct plant materials, we still neither know for sure how EVs pass through the apoplastic space or cell walls, nor what their function truly is. Considering the inhibitory effects of plant nanovesicles against pathogenic microorganisms and increased vesicle amounts after fungal infections in plants [
8,
10,
11,
17,
25] together with the diversity of plant nanovesicle lipids [
17,
18,
24,
26], one can assume that plant cells produce EVs in order to control the growth of pathogenic microorganisms in case those try to invade the plant and also to address distant host cells, e.g., to improve their immune response. Taking into account that some pests mainly infect particular plants or plant families, it is plausible to imagine that not only the vesicle’s cargo is tuned by the host but also the envelope, to fight such pests as effectively as possible.
Lipid profiles have been shown to vary between different species and we therefore assume, so far, that lipid compositions are species-specific [
17], although the extent of variations within the membrane composition in one and the same species remains unknown. Recent results by Berger et al., 2020 indicate that there might also be a family specificity [
24]. Anyways, qualitative and quantitative differences of vesicle lipids can hypothetically be utilized to target particular cell types or organs. This assumption is based on the finding that nanovesicles from certain plant species are preferentially taken up by particular cell types [
27]. As such, grape exosome-like nanoparticles can target intestinal stem cells [
26] while ginger-derived nanovesicles were taken up by hepatocytes and inhibited alcohol-induced liver damage [
22]. Likewise, semi-synthetic nanoparticles that have been engineered from grapefruit nanovesicle lipids, were delivered to the brain after nasal application [
28] or to liver macrophages after intravenous administration [
29]. If research could link the described lipid profiles with specific uptake capabilities into certain cells, we would have a promising tool for targeted drug delivery at hand. In order to get one step closer to that aim, we herein present TLC lipid profiles of some selected plants.
Another aspect of plants’ defense against pathogenic microorganisms might be the packaging of secondary metabolites with anti-microbial activity into EVs. The association of secondary metabolites with plant nanovesicles is also of some relevance regarding the bioavailability of these molecules, as it might increase significantly when secondary metabolites are vesicle-encapsulated. Unfortunately, the few available data on this issue are ambiguous—on the one hand naringin, naringenin [
30] and shogaol [
22] were found in nanovesicles from grapefruit and ginger. On the other hand are the results of Berger et al., 2020, showing the absence of vitamin C and naringenin in orange nanovesicles [
24]. To study the scope of nanovesicle-associated secondary metabolites, we investigated the presence of distinct characteristic alkaline and acidic constituents in nanovesicle-isolates from several pharmaceutically and toxicologically relevant herbal drugs.
Apparently, plant nanovesicles do not exhibit any toxic or immunogenic effects [
31], but for the pharmaceutical use of the described nanostructures, a richer knowledge on their properties would help to ensure quality. Identifying characteristic proteins and enzymes that are typically associated with plant nanovesicles could provide both marker proteins for purposes of isolation and analytics, as well as better understanding of (sub-)cellular mechanisms regarding EV formation and distribution.
For the pharmaceutical application of plant nanovesicles, either by themselves or for drug delivery, we need adequate sources for the production of medicinal products with sufficient and constant quality and safety. The establishment of plant cell cultures and their bioprocess optimization could provide a well controllable foundation for this purpose.
In the present article, we show that EVs can be successfully isolated from plant cell culture media following a straightforward procedure that can be easily scaled up to higher volumes. We further provide information on the proteome, as well as on lipids and secondary metabolites of EVs from plant cell culture, as well as of nanovesicles from dried herbs.
2. Results
2.1. Electron Microscopy
Lacking knowledge on marker proteins of plant EVs or nanovesicles at the current state, we imaged the samples that were supposed to contain nanometer-scaled vesicles, using transmission electron microscopy (TEM) in order to confirm the success of the isolation. We observed vesicles with cup-shaped morphology, which has been reported repeatedly for EVs as a resulting artifact of TEM preparation [
32]. Morphological differences between EVs and nanovesicles have not been observed, as shown in representative TEM images in
Figure 2.
Besides the mainly used isolation technique of differential centrifugation, plant EVs have also been concentrated by tangential flow filtration (TFF, HansaBioMed Life Sciences, Tallinn, Estonia) from cell culture medium (
Figure 2d). Since we further noticed that ultrafiltration was not suitable for nanovesicle isolation from herbal drugs due to co-concentration of accompanying polymeric substances, TFF was not utilized for this kind of raw material.
2.2. Secondary Metabolites
A crucial factor in secondary metabolite analysis is pH. Consequently, we chose two buffers for our investigations of either acidic or alkaline analytes. When searching for acidic components like curcuminoids, flavonoids or caffeic acid derivatives, a Tris-buffered saline (TBS) pH 8.0 was used (100 mM Tris, 100 mM NaCl, 10 mM EDTA, HCl q.s.) intending to deprotonate acid groups. Due to the resulting charge and thus lipophobicity, these structures should not be able to pass the membrane barrier and would stay either inside or outside the vesicle, depending on their original location.
Conversely, if vesicle samples were investigated for alkaloids, a buffer with acidic pH was used for isolations. We therefore modified the vesicle isolation buffer (VIB) by Rutter and Innes 2017 [
13] and used it as follows: 20 mM 2-(
N-morpholino)ethanesulfonic acid (MES), 100 mM NaCl, 10 mM EDTA, NaOH q.s. (to adjust pH to 5.5).
The TLC-chromatogram of curcuminoids in nanovesicles from
Curcumae zanthorrhizae rhizoma,
Curcuma zanthorrhiza Roxb. (
Figure 3) shows the influence of pH on curcuminoid solubility in ethyl acetate. While curcuminoids dissolved in the organic solvent when nanovesicles were prepared in acidic VIB, the deprotonated form present in alkaline TBS had to be acidified prior to extraction.
Figure 3 further indicates that curcuminoids were enriched in Javanese turmeric nanovesicles compared to the aqueous supernatants and that the washing step efficiently removed soluble contaminants.
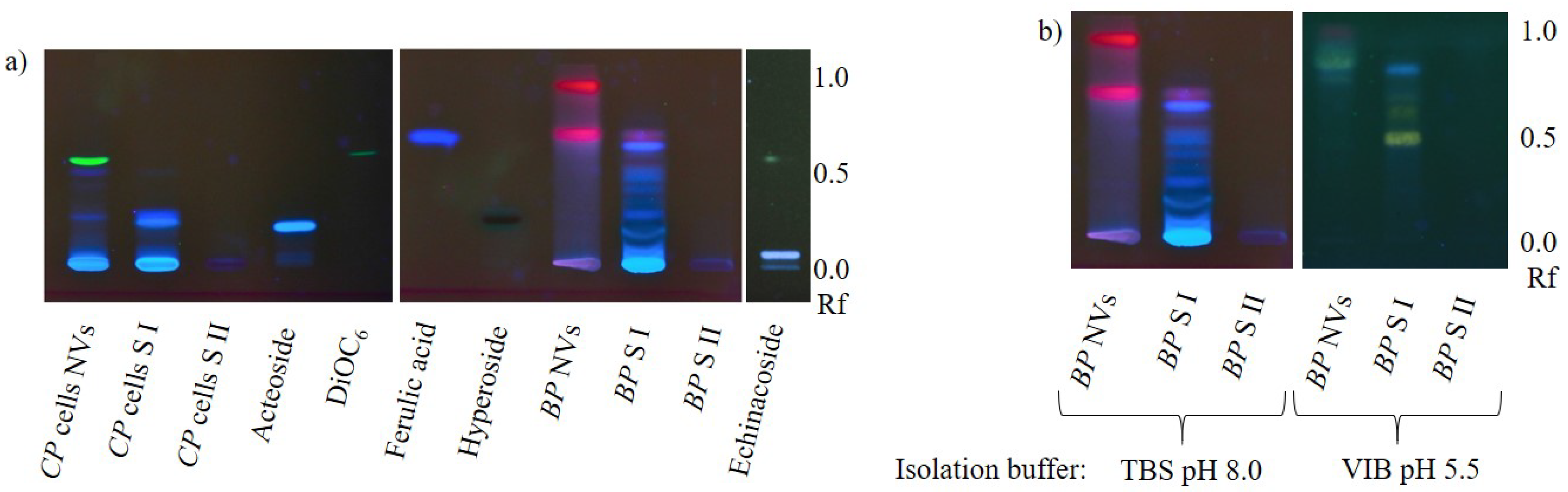
After this first indication that secondary metabolites could be found in plant nanovesicles, we also investigated nanovesicles from dried
Aconiti tuber, Aconitum napellus L. and
Uvae-ursi folium,
Arctostaphylos uva-ursi (L.) Spreng. as well as EVs from
Nicotiana tabacum L. leaves’ apoplastic fluid for the occurrence of characteristic compounds (alkaloids and phenolics). Chromatograms in
Figure 4 show that neither of the investigated vesicle sample contained detectable amounts of the respective analyte, while characteristic metabolites were determined in each aqueous supernatant of the first high speed centrifugation step (S I).
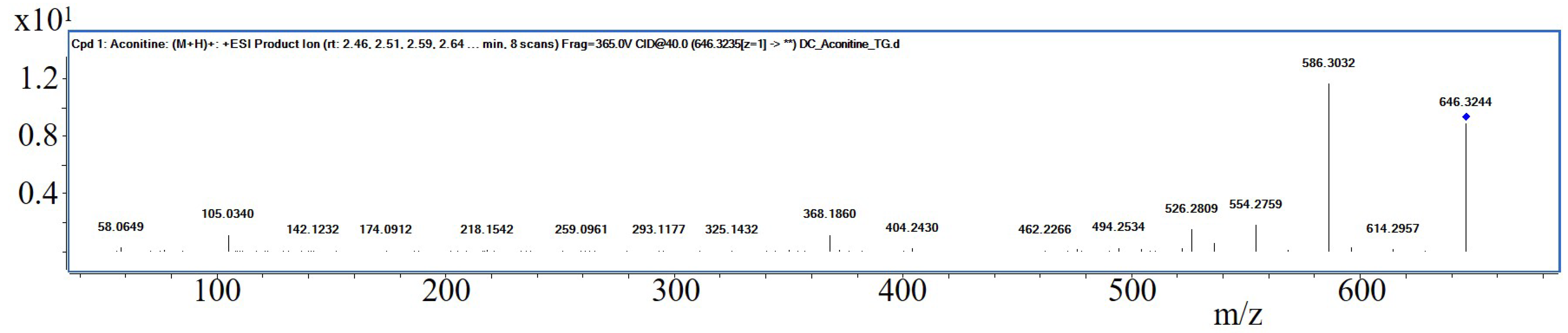
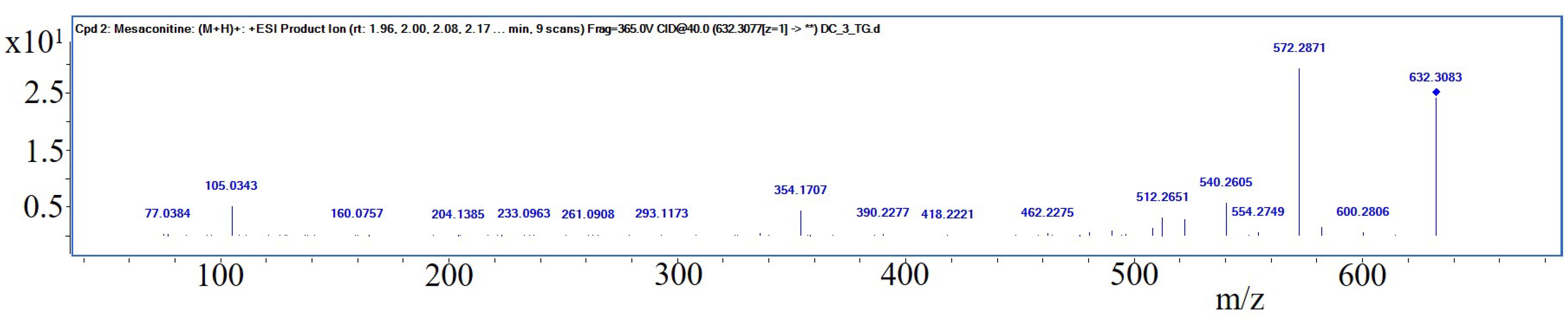
Interestingly, S I of
Aconiti tuber contained only traces of aconitine, while the two prominent bands in the relating track have been identified putatively being hypaconitine and mesaconitine by liquid chromatography—mass spectrometry (LC-MS, PharmaMS, Core Facility BioSupraMol, Freie Universität Berlin, Germany) after reextracting the excised bands from the HPTLC plate. Aconitine was used as positive control. Fragments (For product ion spectra see
Appendix A,
Figure A1,
Figure A2 and
Figure A3) appeared to confirm the identity of the norditerpene alkaloids (The absorbance spectra correlation between aconitine reference and the two derivatives was 97.6% and 98.9% and the substances showed positive reactions with Dragendorff’s reagent.) according to [
33]:
Phenylpropanoid analysis in nanovesicles from
Craterostigma plantagineum Hochst. cells and NVs from dried
Betulae folium,
Betula pubescens Ehrh. and/or
Betula pendula Roth showed results that appeared to be ambiguous at first sight (
Figure 5a). While the blue-fluorescing zones in
C. plantagineum nanovesicles indicated the occurrence of caffeic acid derivatives in this sample, similar bands were not detected in birch nanovesicles. Instead, nanovesicles from birch leaves contained red-fluorescing lipophilic substances, probably chlorophylls. Considering the fact that chlorophylls are physiologically mainly present in chloroplasts jointly with the above-reported results, we hypothesize that secondary metabolites (or comparable molecules of the primary metabolism) are not actively packaged into plant EVs. Further, lipophilic structures appear to be enriched in vesicle membranes, similarly to the membrane dye 3,3’-dihexyloxacarbocyanine iodide (DiOC
6). Chlorophylls were likely released due to chloroplast rupture during herb drying.
However, caffeic acid derivatives should not occur in
C. plantagineum nanovesicles if our theory was consistently valid, but taking the pH of the growth medium of the plant cell culture into account, the hypothesis still stands. The medium pH of 5.8 would allow caffeic acid derivatives to pass vesicle membranes. Regarding the pk
a of caffeic acid (pk
a = 4.62, referring to
https://pubchem.ncbi.nlm.nih.gov/compound/Caffeic-acid#section=pKa (accessed on 2 March 2021)), around 7% of the molecules would be present in the protonated neutral form, which is able to make its way into the vesicle by diffusion and establish equilibrium. This assumption is underlined by the findings presented in
Figure 5b. The chromatograms of birch nanovesicles and related supernatants from the isolation process, in context with the pH of buffer used for the vesicle isolation have been compared. As expected, isolating the vesicles in a slightly acidic environment resulted in the presence of phenylpropanoids in the nanovesicle sample. The high retention factor (Rf) of the yellow- and blue-fluorescing zones, which were likely caused by flavonoids and caffeic acid derivatives, demonstrate their hydrophobicity letting us assume, that mainly aglyca were enriched here, not glycosides.
2.3. Proteomics
Protein identification from
N. tabacum leaf apoplastic fluid-derived EVs was troubled by ribulose-1,5-bisphosphate carboxylase-oxygenase (RuBisCO). This extremely highly abundant enzyme—probably the most abundant protein in nature [
34]—appeared as a contamination throughout the whole lane of the sodium dodecyl sulfate—polyacrylamide gel electrophoresis (SDS-PAGE) of isolated EV samples, severely impairing the identification of additional proteins in excised bands via peptide mass fingerprint and matrix assisted laser desorption ionization—time of flight—mass spectrometry (MALDI-TOF-MS). Nonetheless, two proteins could be identified: 40S ribosomal protein S4 ≈ 30 kDa and 40S ribosomal protein S6 ≈ 22 kDa (both confirmed by MS-MS data). Related proteins have already been reported in plant nanovesicles [
10,
13,
14,
19] but they do not appear to be very specific for plant EVs.
As a consequence of the RuBisCO interference, another approach towards protein identification was made with EVs from N. tabacum in vitro cell culture medium, this time isolated by TFF. Here, the ≈85 kDa hydrolase -xylosidase/-L-arabinofuranosidase 2-like protein (confirmed by MS-MS, apparently a mixture of two or more isoforms) was reliably identified as an EV protein.
Following the thread of proteomics,
C. plantagineum EVs from in vitro cell culture medium and nanovesicles from homogenized cells were also investigated using a LC-MS-based approach. We identified proteins in two batches of EVs and nanovesicles isolated from
C. plantagineum cell suspension cultures. The two batches differ in the amount of input material for proteomics—the second batch allowed the identification of more proteins exclusively present in EVs or nanovesicles. The results of the first and second batch were compiled in
Supplementary Materials, respectively and selected results in
Table 1. The complete MS proteomics datasets are available via ProteomeXchange with the identifier PXD024203 via
http://www.ebi.ac.uk/pride/archive/projects/PXD024203 (accessed on 2 March 2021).
2.4. Phospholipid Profiles
In order to validate or refute our hypothesis that the phospholipid composition influences the uptake of nanovesicles into certain cell types or specific organs, two main aspects have to be taken into account: (i) To which extent do the lipid compositions of plant nanovesicles vary? (ii) Are these variations causally linked to any cellular uptake preferences?
To investigate the first aspect, we used an HPTLC method to get a first insight into plant phospholipid profiles. Preliminary investigations showed that sterols (-sitosterol und stigmasterol) were not retained by the stationary phase and retention factors resulted to be nearly 1. Further, the HPTLC of 1,2-dioleoyl-sn-glycero-3-phosphocholine and 2-oleoyl-1-palmitoyl-sn-glycero-3-phosphocholine has shown no differences in retention factors (data not shown). Thus, the phospholipid class apparently influences the migration distance of the lipid more than individual fatty acid residues.
Although the application of single phospholipids provided different retention factors (
Figure 6), phosphatidylethanolamine and phosphatidylinositol were not satisfactorily separated when the five phospholipid classes were applied together. Nonetheless, the method was used to get a first insight into the phospholipid profiles of a row of nanovesicle extracts and as the chromatogram in
Figure 6 shows, we were able to identify phospholipids in these samples as follows:
Uvae-ursi folium nanovesicles: Phosphatidic acid
C. plantagineum nanovesicles: Phosphatidylcholine, Phosphatidic acid, Phosphatidylethanolamine
Curcumae zanthorrhizae rhizoma nanovesicles: Phosphatidylinositol
Zingiberis rhizoma nanovesicles: Phosphatidic acid, Phosphatidylethanolamine, Phosphatidylinositol
Our investigations also revealed that relatively large amounts of vesicles were necessary to achieve analyzable chromatograms. 1–2 μg of each phospholipid were clearly detectable. Meanwhile, a minimum of 1 mg (calculated as total protein amount) of a vesicle sample had to be extracted. Therefore, other investigated isolates (e.g., nanovesicles from Aconiti tuber, Betulae folium and Uvae-ursi folium) remained without reportable results.
3. Discussion
Beyond doubt, the research on EVs from all domains of life has already revolutionized our understanding of intercellular information transmission, but, in particular with regard to plant-derived EVs, its potential is still waiting to be fully tapped. With our present investigation we intended to keep plant nanovesicles in the scientific focus and move a little closer towards the usage of these promising structures. Our finding that plant EVs can be isolated from plant in vitro culture media might pave the way towards that aim.
Compared to the vesicle isolation process from conventionally grown and harvested plant materials, the amount of accompanying substances would be minimized when cell culture media were used as starting point. Besides, the production could take place under controlled sterile environmental conditions, which ought to ensure product consistency and maybe plant cells could even be induced, by additives, to produce EVs with desired properties, or loaded with drugs by plant cells themselves. Differences between the metabolisms of humans and plants could even allow the application of drugs that cannot be packaged into vesicles in in vitro cultures of human cells (e.g., due to cytotoxic effects). However, before this perspective can be translated into first studies, more knowledge on plant nanovesicles must be compiled. We therefore gathered information on secondary metabolites, the proteome and lipids of plant EVs and nanovesicles in the investigation at hand.
3.1. Secondary Metabolites
Due to the complexity of nanovesicle samples containing nucleic acids, proteins, lipids and salts from isolation buffers, HPTLC appeared to be the chromatography technique of choice, since it is especially robust, yet sensitive and can be applied for a broad set of analytes. Secondary metabolite analysis in nanovesicles from several plant species revealed that especially lipophilic molecules were vesicle-associated. As such, we found curcuminoids and chlorophylls being enriched in the investigated vesicles, just like DiOC
6, if the membrane dye had been added. Meanwhile, neither alkaloids nor phenols or phenylpropanoids appeared to be typically associated with the isolated vesicles. Analogously, Stremersch et al., 2016 reported that cholesterol-modified siRNA bound to the vesicle membrane surface with the cholesterol residue acting as anchor [
35]. We therefore conclude that secondary metabolites are associated to nanovesicle membranes in a passive manner due to lipophilicity, rather than being actively packaged into the vehicles.
The above-mentioned ambiguous data about secondary metabolites in plant nanovesicles are not in contradiction with our theory, since shogaol, which has been found in ginger nanovesicles [
22], is chemically and biogenetically closely related to curcuminoids. The reports about flavonoids are not consistent since naringin and naringenin were found in grapefruit nanovesicles [
30] but ascorbic acid as well as naringenin were absent from nanovesicles from orange [
24]. Howsoever, in the acidic milieu of citrus fruits, we do not question the possibility of diffusion-driven flavonoid distribution through all compartments.
From an evolutionary point of view, thinking that secondary metabolites are not actively packaged into EVs is not odd. The high conservation of EV formation and the fact that practically every living cell secretes vesicles [
36], makes it appear likely that mechanisms of EV formation had been established in eukaryotic metabolism prior to the development of plant secondary metabolite production.
What still remains puzzling is the extent to which plant cells may vary EV compositions. Similar to what is known from the animal kingdom [
37], plants could tune EVs as a response to certain physiological or environmental conditions. As such, approaching stimuli might induce the formation of vesicles aligned to meet the requirements of the particular signal.
Among the objectives of further investigations could be a more comprehensive screening of a greater variety of species and analytes, together with a clustering of n-octanol-water partition coefficients of vesicle-associated substances and the evaluation of differences between aglyca and glycosides. A possible attempt to prove our hypothesis might also be to grow plant cell suspension cultures in media with different pH ranges and analyze EVs for secondary metabolites.
3.2. Proteomics
The high abundance of RuBisCO hindered the identification of lower-abundance proteins, underlining the importance of a careful evaluation of the results of proteomic investigations. Additionally, reconfirmation is required to clarify whether the proteins are indeed structural EV components and not just co-isolated. Considering this constraint, the list of proteins in
Table 1 was thoughtfully selected from the proteins identified in EVs or NVs but not in the supernatants S I and S II of high-speed centrifugation.
It is remarkable that the hydrolase
-xylosidase/
-L-arabinofuranosidase 2-like was found in the vesicles of both investigated species
C. plantagineum and
N. tabacum. This enzyme is involved in cell wall remodeling [
38,
39] and might be responsible for the passage of EVs through cell walls, which has not yet been conclusively elucidated [
13]. One aspect of the hypothetical passage mechanisms is based on the assumption that EVs are associated with enzymes loosening the cell wall structure. The identified hydrolase could be one of these enzymes. Guerra-Guimarães et al., 2014 found this hydrolase to be prominently abundant in apoplastic fluid from
Coffea arabica L. leaves [
38]. Regarding that in this work proteins were concentrated by ultrafiltration, apoplastic EVs might have been concentrated unintentionally.
Besides
-xylosidase/
-L-arabinofuranosidase 2-like, we identified a couple of other proteins involved in cell wall reorganization, many of them exclusively in the vesicle fraction (
Table 1). Interestingly, even an endochitinase EP3-like was detected, indicating the relevance of EVs as defensive agents against pathogenic fungi.
In order to estimate cellulase-/hemicellulase activity, zymograms could perspectively help to evaluate enzyme activity and thus the extent to which hydrolases are correlated to the vesicles themselves or rather surrounded by them. If the vesicle fraction appears to be active, we might have a marker protein at hand. Other potential marker proteins are listed in
Table 1 “Reported in plant NVs or EVs”.
Another interesting finding is the joint appearance of transmembrane 9 superfamily member 11, AP-complex subunits and membrane steroid-binding protein 2, as well as proteins that are related to ubiquitination. The presence of these proteins indicates that the isolated vesicles might originate not only from endosomal pathways with Golgi-apparatus involvement, but also directly from the plasma membrane, hinting at the correctness of our hypothesis that plant cells secrete exosomes, as well as microvesicles, in a similar manner as animal cells do.
3.3. Phospholipid Profiles
Plant biomembranes consist of glycerolipids, sphingolipids and sterols [
40]. Among glycerolipids, the phospholipid group appears to play a crucial role not only for nanovesicle stability but probably also for addressing target cells. Although the accuracy of our method can yet be improved, this technique is broadly applicable and high throughputs are possible. The observed need of substantial sample amounts has been reported similarly by other groups [
18,
41]. Despite these obstacles, we were able to get a first insight into the phospholipid profiles of nanovesicles from a panel of different plant species.
Creating such lipid profiles of a variety of nanovesicles from the plant kingdom might improve our understanding of EV and nanovesicle formation, addressing and uptake, as well as related synthetic nanocarriers. If we were once able to address specific target cells with biocompatible drug-loaded nanocarriers, side effects could be brought down to an absolute minimum. Comparative studies of nanovesicles and EVs from one and the same plant, like recently published by Liu et al., 2020 [
1], may also help improve our knowledge, probably even beyond the fields of lipids and cellular uptake.
4. Materials and Methods
4.1. Isolation of Nanovesicles and EVs
Plant EVs can be obtained from apoplastic fluid [
12,
13] but in the investigation at hand, we also isolated EVs from plant cell culture media. Moreover, nanovesicles were isolated from homogenized plant materials, such as dried powdered herbs or from in vitro cultured plant cells. For vesicle rehydration, 50 g dried herbs were incubated overnight in 500 mL of the respective buffer (see
Section 4.3) at 4 °C. In vitro cultured plant cells were ground with ice-cooled mortar and pestle and PBS added respectively. The suspensions of either cultured cells or herb powder were decanted and centrifuged differentially. All centrifugation steps were performed at 4 °C. The aqueous extracts were centrifuged twice at 4000×
g for 10 min to remove large particles (Beckman Allegra X 30 R centrifuge, SX 4400 rotor; Beckman Coulter, Brea, CA, USA). The supernatant was then centrifuged 15 min at 20,000×
g removing medium size particles (Avanti J-26 S XP centrifuge, JA 25.50 rotor; Beckman Coulter). The supernatant was filtered through a 0.85 μm syringe filter (Rotilabo
® CME, Carl Roth GmbH & Co. KG, Karlsruhe, Germany), membranes optionally stained using DiOC
6 and then nanovesicles pelleted at 50,000×
g; 90 min (Avanti J-26 S XP centrifuge, JA 25.50 rotor). The resulting pellet was washed using freshly 0.2 μm filtered buffer and the final centrifugation step repeated. The pellet, containing isolated nanovesicles, was suspended in 100–1000 μL filtered buffer, depending on the yield. After isolation, protein concentrations of the vesicle samples were determined by Roti
®Nanoquant Bradford assay (Carl Roth) according to the manufacturer’s manual.
For the article at hand, plant EVs were isolated from the apoplastic fluid of
Nicotiana tabacum L. (tobacco seeds were provided by the Botanical Garden Berlin, Germany; accession number 107-01-95-14, and grown in the greenhouse at the Institute of Pharmacy, Freie Universität Berlin, Germany). Apoplastic fluid was obtained as previously described [
42] according to Rutter and Innes 2016 [
13] using the vacuum-infiltration centrifugation technique.
EVs were further concentrated from suspension culture media of
Craterostigma plantagineum Hochst. and
N. tabacum. The
C. plantagineum calli were dedifferentiated from leaves of plants supplied by Prof. Dr. Dorothea Bartels (Rheinische Friedrich-Wilhelms-Universität Bonn, Germany) and cultured as described previously [
43]. Cell cultures were grown for 2 weeks under light in 2 L flasks at 100 rpm and 26 °C.
Nanovesicles were isolated from the above-mentioned cultured plant cells, as well as from dried herbs of: Aconitum napellus L., Arctostaphylos uva-ursi (L.) Spreng., Betula pubescens Ehrh. and/or Betula pendula Roth, Curcuma zanthorrhiza Roxb. and Zingiber officinale Roscoe (all purchased from Alfred Galke GmbH, Bad Grund, Germany).
Suspension culture media and cells of N. tabacum were kindly provided by the Department of Pharmacognosy and Herbal Medicines of Wrocław Medical University, Wrocław, Poland.
4.2. Electron Microscopy
Isolated nanovesicles were visualized by TEM, using the scanning electron microscope Hitachi SU 8030 in TEM mode (Hitachi Ltd., Tokyo, Japan). Samples were prepared as earlier described [
42]: 5 μL of each sample were placed on 300 mesh Formvar and carbon-coated copper grids and incubated 1–2 min. Grids were then rinsed and negatively stained by pipetting ≈80 μL Uranyless
® (Science Services GmbH, Munich, Germany) across the grid surface. Fluid excess was carefully blotted using Kimwipe before overnight drying in a desiccator. TEM imaging was performed using 30 kV acceleration voltage.
4.3. Secondary Metabolites
The buffer for vesicle isolations has been selected regarding the secondary metabolites that ought to be investigated. For the analytics of alkaline structures, VIB was used—a MES buffer of pH 5.5 (20 mM MES, 100 mM NaCl, 10 mM EDTA, 10 M NaOH to adjust pH to 5.5), while we chose TBS of pH 8.0 (100 mM Tris, 100 mM NaCl, 10 mM EDTA, 10% HCl to adjust pH to 8.0) when acidic metabolites were in the focus of investigation. This way, we intended to inhibit the membrane passage of secondary metabolites—alkaloids would be positively charged due to protonation in acidic environment and acidic compounds were negatively charged because of deprotonation under basic conditions.
To evaluate the content of secondary metabolites, 200 μL of each sample have been lyophilized (Alpha 1–2 LDplus, Martin Christ GmbH, Osterode am Harz, Germany) and afterwards extracted in 90 μL of an organic solvent (e.g., ethyl acetate or a mixture of equal parts methanol and chloroform). Due to the pH of buffers used for vesicle isolation, secondary metabolites had to be neutralized prior to extraction. Therefore, 10 μL of formic acid were added to protonate acidic molecules while 10 μL of 10 M NaOH were added deprotonating alkaloids. After thorough mixing and brief centrifugation in a benchtop centrifuge (Heraeus Biofuge pico, Heraeus GmbH, Hanau, Germany) to settle undissolved solids at the bottom of the reaction tube, 5–10 μL of supernatants were applied band-shaped onto HPTLC plates (Nano-SIL-20/UV
254, Macherey-Nagel GmbH & Co. KG, Düren, Germany) by Linomat IV (Camag AG, Muttenz, Switzerland). Alkaloid HPTLC development was performed automated in an AMD2 device (Camag) following the gradient in
Appendix A,
Table A1. Plates for acidic metabolite analysis were also developed automated, using ADC2 (Camag). Mobile phase compositions are listed in
Appendix A,
Table A2.
After development, plates were documented under UV light at 254 and 366 nm and scanned by TLC Scanner 4 (Camag) at 210, 250 and 280 nm at first. These first scans were used to create absorption spectra of selected bands and optimize the detection wavelength of further scans to the maxima of the analytes of interest. Identities of bands with similar Rf were confirmed or disproved by matching the absorption spectra. After the described densitometric analysis, plates were eventually derivatized with Dragendorff’s reagent or 5% 2-aminoethyl diphenyl borate in methanol.
4.4. Proteomics
Proteins of plant extracellular- and nanovesicles have been identified from leaves’ apoplastic fluid or suspension cultured cells of the two plants N. tabacum and C. plantagineum. The vesicle samples were usually isolated by differential centrifugation as described above, while tobacco EVs from cell culture medium were isolated using a tangential flow filter (TFF-Easy, HansaBioMed Life Sciences Ltd., Tallinn, Estonia) according to the manufacturer’s instructions. For proteomic investigations, 0.5 mM phenylmethylsulfonyl fluoride (PMSF) were freshly added to the isolation buffer.
To separate proteins by SDS-PAGE, one volume of isolated tobacco EVs was mixed with 4 volumes of reducing 4x Laemmli-buffer (Bio-Rad Laboratories Inc., Hercules, CA, USA) and proteins denatured by boiling at 95 °C for 10 min. The separation itself was performed by discontinuous SDS-PAGE, conducted following the instructions of Jansohn and Rothaemel 2012 [
44] using 12.5% (
w/
v) polyacrylamide resolving gel and 5% (
w/
v) polyacrylamide stacking gel on top. 15–20 μL of each sample were added to gel pockets and electrophoresis performed at 200 V for approximately 45 min, until bromophenol blue reached the bottom of the gel. After electrophoresis, gels were immediately transferred into the staining solution. According to Neuhoff et al., 1988 [
45], gels were stained overnight in 0.1% (
w/
v) Coomassie Brilliant Blue G-250, 2% (
v/
v) H
3PO
4, 10% (
w/
v) (NH
4)
2SO
4, and 20% (
v/
v) methanol. The next day, gel matrices were destained using 25% (
v/
v) methanol.
Out of these gels, visible protein bands were excised and proteins in-gel digested with trypsin (for MS from porcine pancreas, SERVA Electrophoresis GmbH, Heidelberg, Germany) according to Shevchenko et al., 1996 [
46]. Prior to trypsin digestion, proteins were destained, reduced and carbamidomethylated. Digestion supernatants were then investigated by MALDI-TOF-MS (Ultraflex-II TOF/TOF, Bruker Daltonics, Bremen, Germany) with
α-cyano-4-hydroxycinnamic acid as matrix. Database searches for protein identification of the peptide mass fingerprints were performed using Mascot (Matrix Science Ltd., London, UK,
http://www.matrixscience.com (accessed on 2 March 2021)) against the SwissProt database (all entries). The mass tolerance was set at ±75 ppm and we allowed for one missed cleavage. Carbamidomethylation (CAM) of cysteines was set as a fixed modification, and oxidation of methionines as variable modification. The identifications were confirmed by MS/MS sequencing of selected peptides acquired in the LIFT mode [
47]. Here fragment peptide mass tolerance was set at ±100 ppm and fragment mass tolerance at ±0.7 Da.
Following up on these preliminary proteomic investigations, the methodology has been changed to a more comprehensive protocol using nano LC-MS/MS, investigating C. plantagineum samples from cell culture as follows:
Lyophilized
C. plantagineum samples were directly denatured in fresh lysis buffer (urea 7 M, thiourea 2 M, 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate 0.5% (
w/
v)) and subsequently loaded on a precast gel (Criterion™ XT precast 1D gel 12% Bis-Tris, Bio-Rad) for a short migration. Proteins were stained with Instant Blue (Gentaur BVBA, Kampenhout, Belgium), reduced and alkylated. Thereafter, proteins were digested with trypsin 5 ng/µL (sequencing mass grade, Promega) o/n at 37 °C. The extracted peptides were solubilized to perform protein identification by nano LC-MS/MS, using a NanoLC-425 Eksigent system coupled to TripleTOF
® 6600+ MS (SCIEX, Darmstadt, Germany). The protocol is essentially the one described previously [
48].
The MS data were processed with Mascot (version 2.4.2) using Mascot Daemon interface (version 2.4.2, Matrix Science) by searching against an in-house annotated database of C. plantagineum (288,270 sequences). The parameters were set as follows: peptide tolerance of 20 ppm, fragment mass tolerance of 0.5 Da, maximum two missed cleavages, CAM of cysteine as fixed modification and oxidation of methionine, N-terminal protein acetylation, N-terminal glutamine to pyroglutamate and tryptophan to kynurenine as variable modifications. Only the proteins identified with a significance Mascot-calculated threshold corresponding to a p-value < 0.05 and at least two sequences per protein and one unique sequence per protein were accepted.
4.5. Phospholipid Profiles
According to the methods of Mu et al., 2014 [
18] and Deranieh et al., 2013 [
49], we established an instrumental HPTLC method to generate profiles of vesicle phospholipids: after vesicle isolation, distinct protein amounts of vesicle samples were lyophilized in glass tubes, avoiding any possible adsorption of lipids to plastic tubes. After adding adequate volumes of a mixture of equal parts methanol and chloroform (≈100–150 μL), samples were thoroughly mixed and then centrifuged for 5 min at 4000×
g (Beckman Allegra X 30 R centrifuge, SX 4400 rotor). 5 and 10 μL of the supernatant solvent (Combining secondary metabolite analysis and lipid profiling was also possible.) were applied band-shaped to HPTLC plates (Nano-SIL-20/UV
254) using Linomat IV and developed in the AMD2 device. The mobile phase gradient is given in
Appendix A,
Table A3. Detection of phospholipids was not possible by densitometric analysis, though HPTLC plates were bathed in 10% (
m/
v) CuSO
4 in 8% (
v/
v) H
3PO
4 and afterwards heated to 140–145 °C, charring analytes [
50].


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





