
Genome-Wide Identification and Characterization of Wheat 14-3-3 Genes Unravels the Role of TaGRF6-A in Salt Stress Tolerance by Binding MYB Transcription Factor

Abstract
:1. Introduction
2. Results
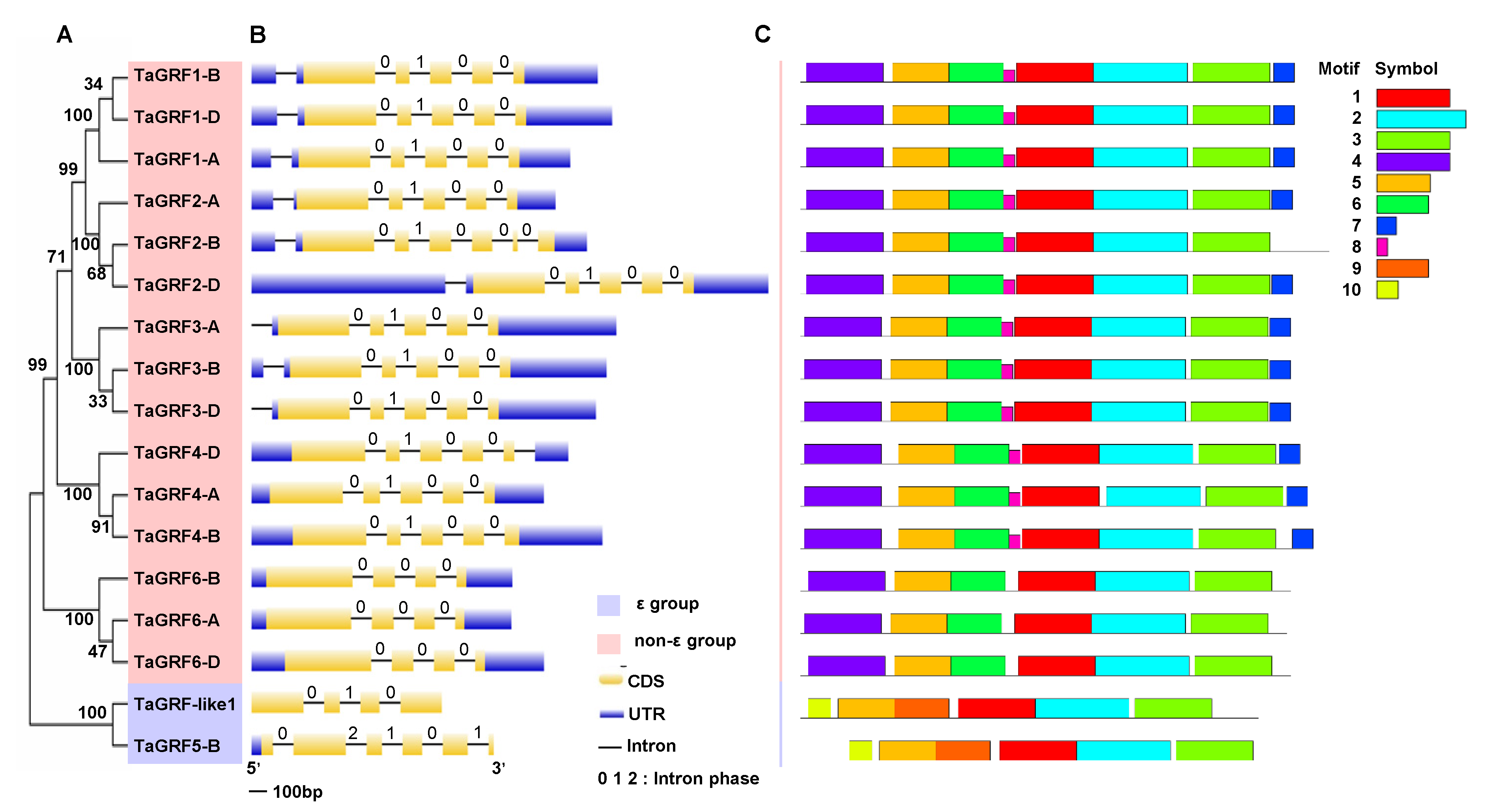
2.1. Genome-Wide Identification and Characterization of 14-3-3 Family Members in Wheat
2.2. Gene Structure and Conserved Motif Analysis
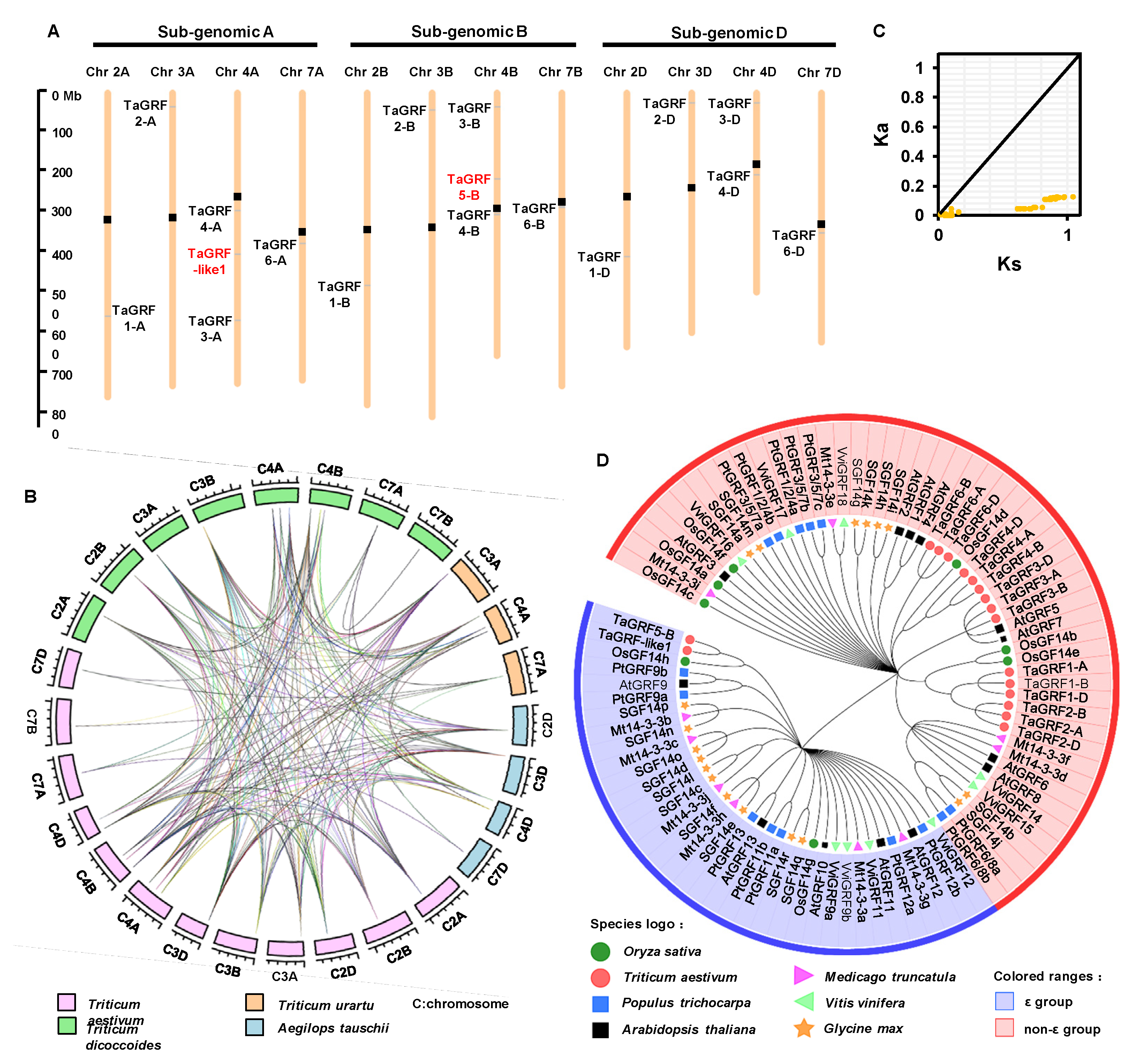
2.3. Chromosomal Distribution and Evolutionary Analysis of TaGRF Genes
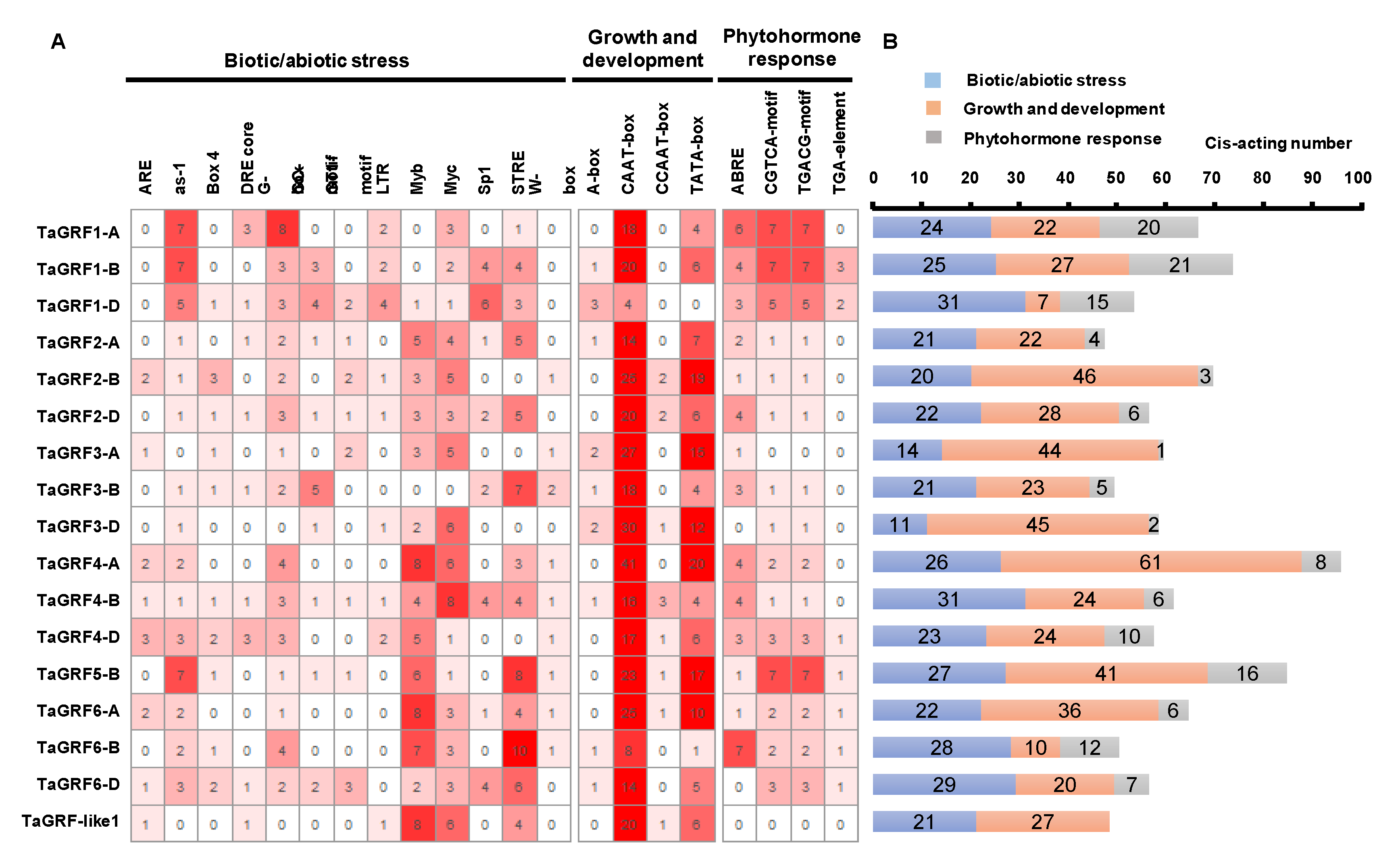
2.4. Cis-Element Analysis and Functional Annotation of TaGRF Genes
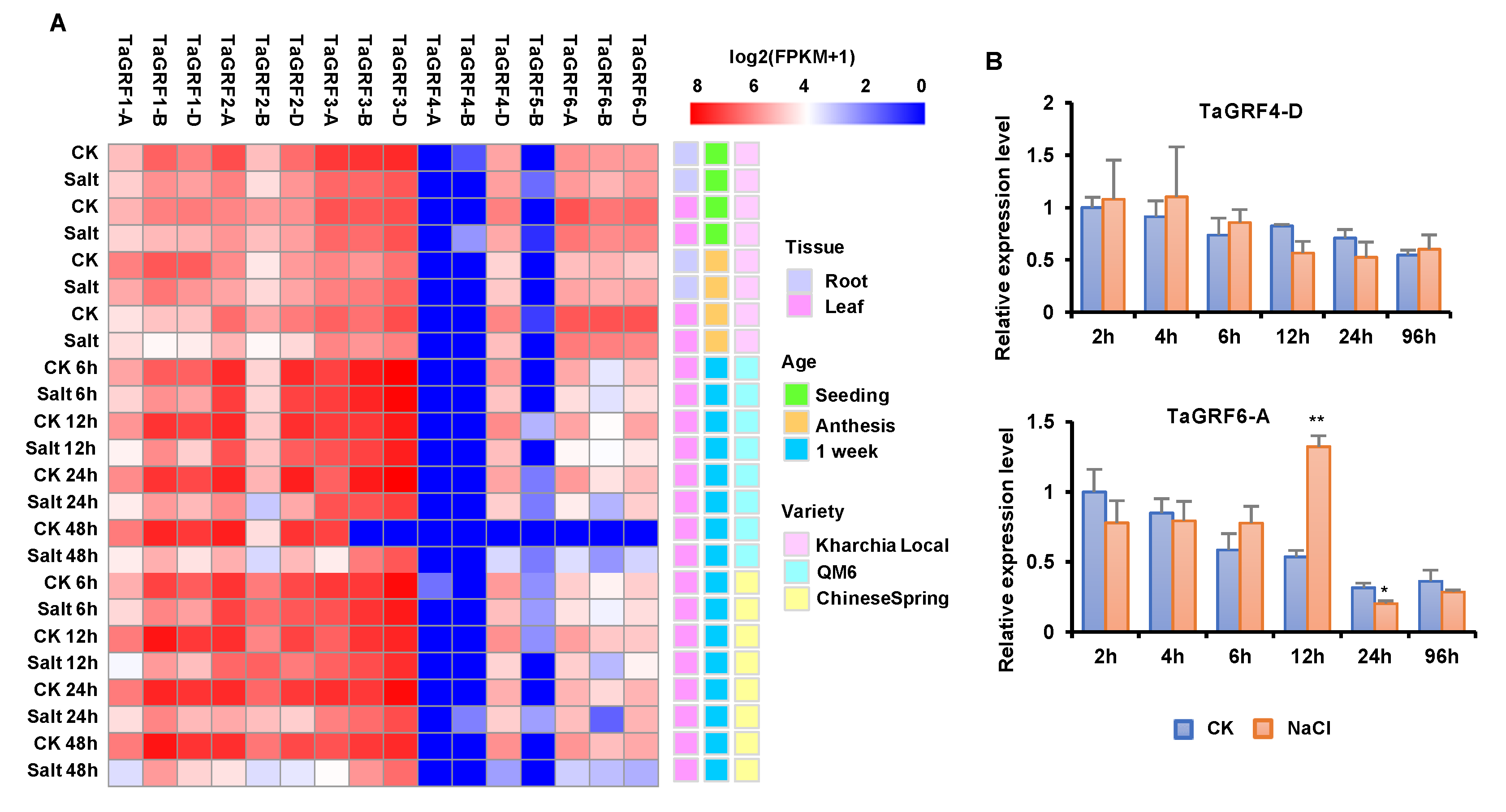
2.5. Expression Profiling of TaGRF Genes
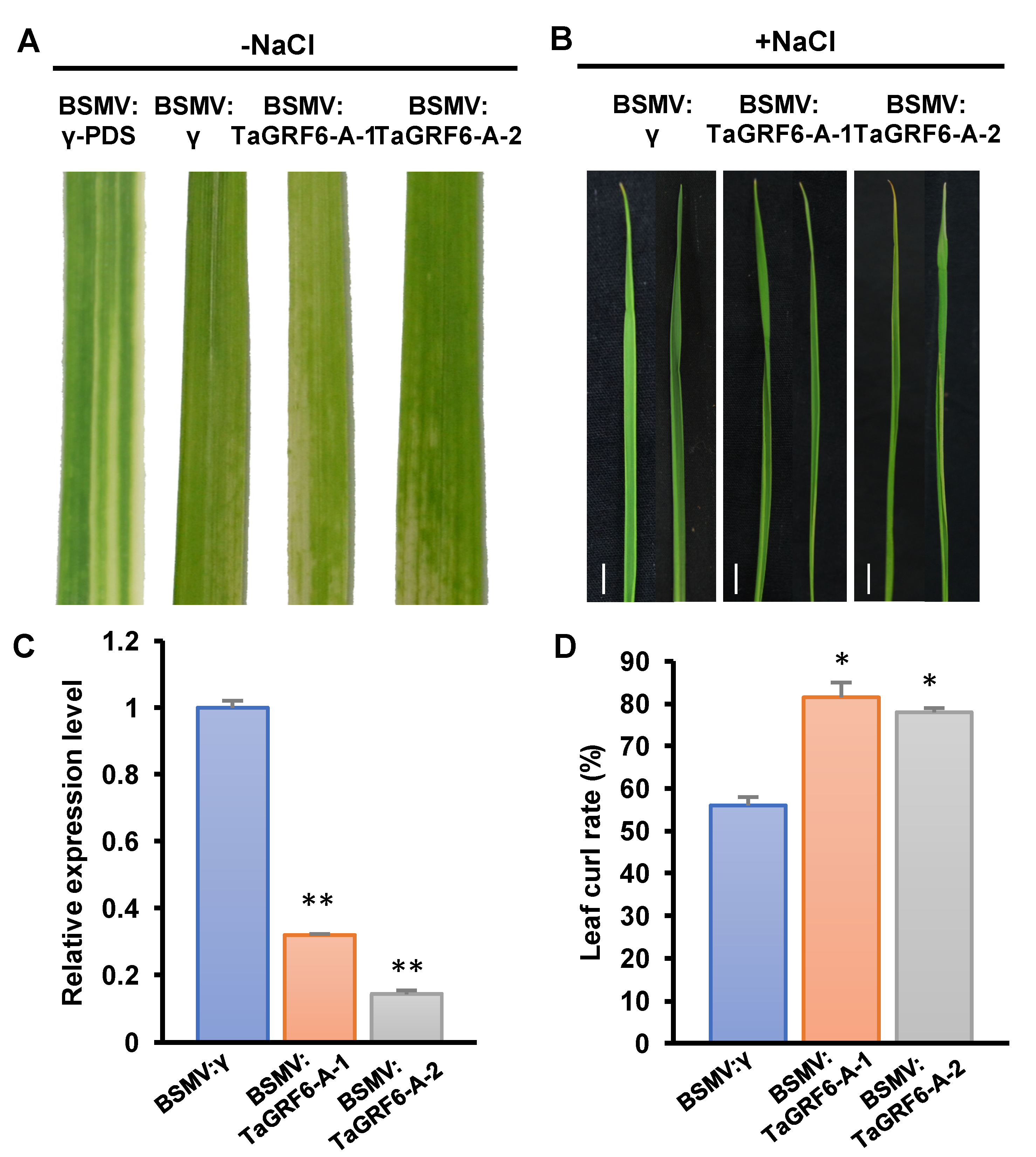
2.6. TaGRF6-A Positively Regulates Salt Tolerance in Wheat
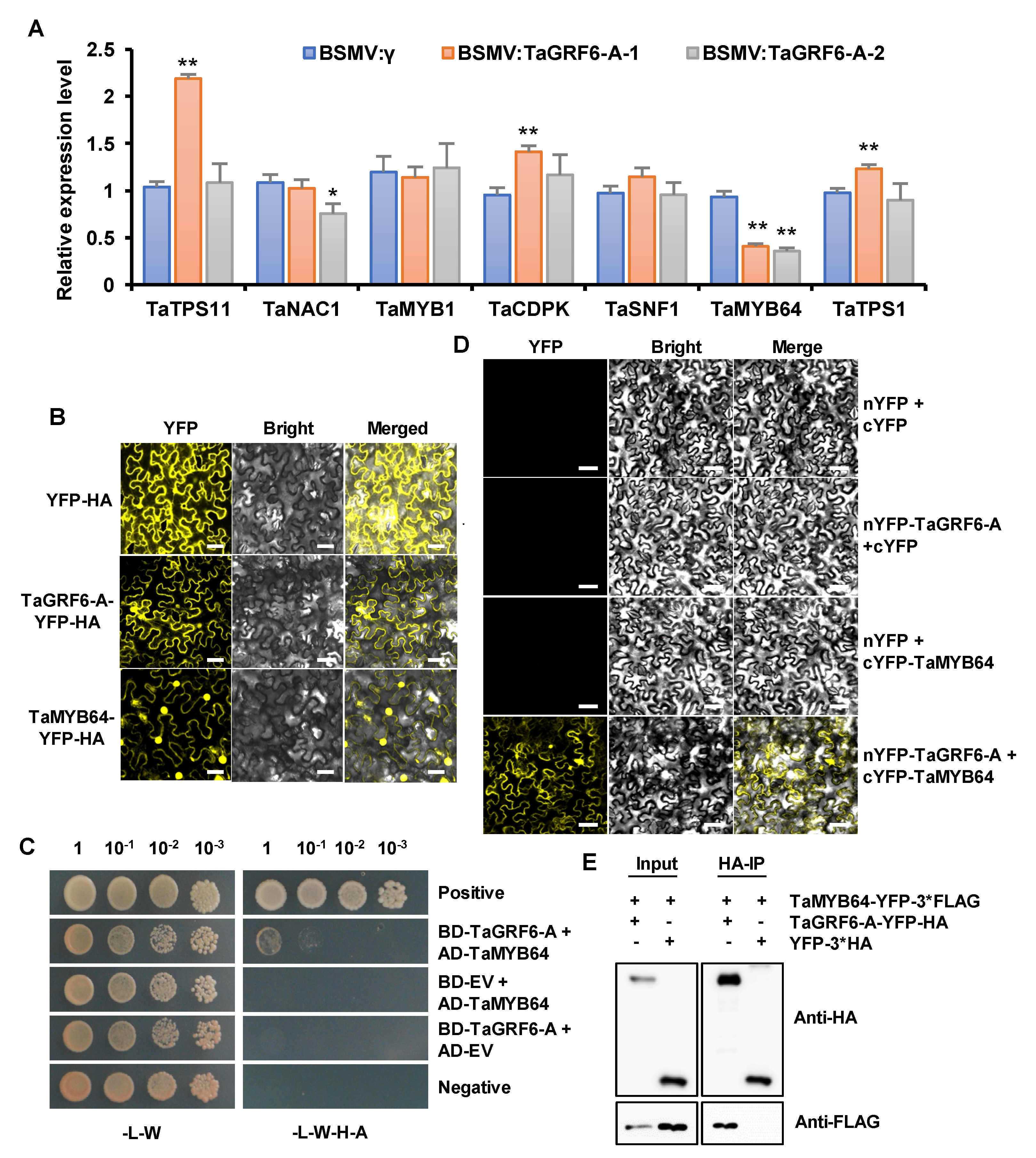
2.7. TaGRF6-A Interacts with TaMYB64 In Vitro and In Vivo
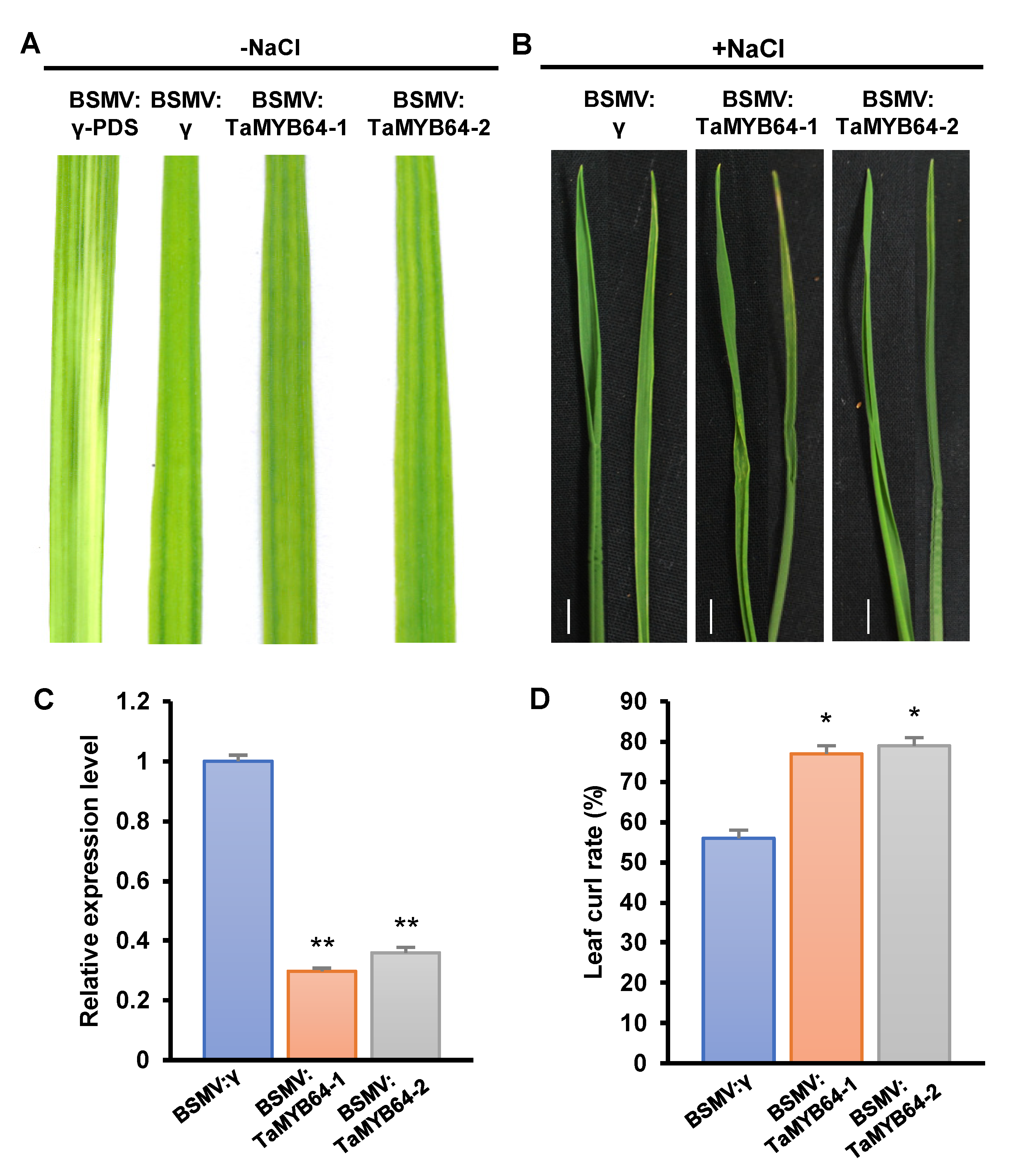
2.8. TaGRF6-A and TaMYB64 Work Together to Cope with Salt Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Salt Treatment
4.2. Mining and Phylogenetic Analysis of Wheat 14-3-3 Genes
4.3. Amino Acid Sequence Alignment and Characterization of TaGRF Proteins
4.4. Gene Structure Analysis and Conserved Motif Prediction
4.5. Chromosome Distribution, Synteny, Ka/Ks, and Phylogenetic Analysis of TaGRFs
4.6. Cis-Acting Element Analyses
4.7. Expression Analysis of TaGRF Genes under Abiotic Stress Conditions
4.8. RNA Isolation and qRT-PCR Analysis
4.9. Virus-Induced Gene Silencing (VIGS) Assay in Wheat
4.10. Subcellular Localization of the TaGRF6-A Protein
4.11. Y2H, BiFC, and CoIP Assays
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferl, R.J. 14-3-3 Proteins and signal transduction. Annu. Rev. Plant Biol. 1996, 47, 49–73. [Google Scholar] [CrossRef]
- Moore, B.W.; Perez, V.J. Specific Acidic Proteins of the Nervous System. Physiol. Biochem. Asp. Neverous Integr. 1967, 343–359. [Google Scholar]
- Ichimura, T.; Isobe, T.; Okuyama, T.; Yamauchi, T.; Fujisawa, H. Brain 14-3-3 protein is an activator protein that activates tryptophan 5-monooxygenase and tyrosine 3-monooxygenase in the presence of Ca2+, calmodulin-dependent protein kinase II. FEBS Lett. 1987, 219, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.; Jones, D.; Patel, Y.; Martin, H.; Madrazo, J.; Martin, S.; Howell, S.; Elmore, M.; Finnen, M.J.; Aitken, A. Mechanism of inhibition of protein kinase C by 14-3-3 isoforms. 14-3-3 isoforms do not have phospholipase A2 activity. Biochem. J. 1994, 299, 853–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braselmann, S.; McCormick, F. Bcr and Raf form a complex in vivo via 14-3-3 proteins. EMBO J. 1995, 14, 4839–4848. [Google Scholar] [CrossRef]
- Yaffe, M.B. How do 14-3-3 proteins work?–Gatekeeper phosphorylation and the molecular anvil hypothesis. FEBS Lett. 2002, 513, 53–57. [Google Scholar] [CrossRef]
- Aitken, A. 14-3-3 Proteins: A historic overview. Semin. Cancer Biol. 2006, 16, 162–172. [Google Scholar] [CrossRef]
- Darling, D.L.; Ling, Y.J.; Boris, W.A. Role of 14-3-3 proteins in eukaryotic signaling and development. Curr. Top. Dev. Biol. 2005, 68, 281–315. [Google Scholar]
- Fu, H.; Subramanian, R.R.; Masters, S.C. 14-3-3 PROTEINS: Structure, Function, and Regulation. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 617–647. [Google Scholar] [CrossRef]
- Hermeking, H.; Benzinger, A. 14-3-3 proteins in cell cycle regulation. Semin. Cancer Biol. 2006, 16, 183–192. [Google Scholar] [CrossRef]
- Muslin, A.J.; Tanner, J.W.; Allen, P.M.; Shaw, A.S. Interaction of 14-3-3 with signaling proteins is mediated by the recognition of phosphoserine. Cell 1996, 84, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Obsilová, V.; Silhan, J.; Boura, E.; Teisinger, J.; Obsil, T. 14-3-3 Proteins: A Family of Versatile Molecular Regulators. Physiol. Res. 2008, 57, S11–S21. [Google Scholar] [PubMed]
- Yaffe, M.B.; Volinia, S.; Leffers, H.; Rittinger, K.; Caron, P.R.; Aitken, A.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The structural basis for 14-3-3: Phosphopeptide binding specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lee, W.H.; Sobott, F.; Papagrigoriou, E.; Robinson, C.V.; Grossmann, J.G.; Sundström, M.; Doyle, D.A.; Elkins, J.M. Structural basis for protein-protein interactions in the 14-3-3 protein family. Proc. Natl. Acad. Sci. USA 2006, 103, 17237–17242. [Google Scholar] [CrossRef] [Green Version]
- Bian, Y.; Lv, D.; Cheng, Z.; Gu, A.; Cao, H.; Yan, Y. Integrative proteome analysis of Brachypodium distachyon roots and leaves reveals a synergetic responsive network under H2O2 stress. J. Proteom. 2015, 128, 388–402. [Google Scholar] [CrossRef]
- Lu, G.; Delislet, A.J.; Vetten, N.C.D.; Ferlo, R.J. Brain proteins in plants: An Arabidopsis homolog to neurotransmitter pathway activators is part of a DNA binding complex. Proc. Natl. Acad. Sci. USA 1992, 89, 11490–11494. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Wang, T.; Xie, Y.; Zhang, J.; Hu, J. Genome-wide identification, classification, and expression analysis of 14-3-3 gene family in Populus. PLoS ONE 2015, 10, 3225. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Wang, Y.; Chai, F.; Li, S.; Xin, H.; Liang, Z. Genome-wide identification and characterization of the 14–3-3 family in Vitis vinifera L. during berry development and cold- and heat-stress response. BMC Genom. 2018, 19, 579. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Fang, Z.; Sun, C.; Zhang, P.; Zhang, X.; Lu, C.; Wang, S.; Ma, D.F.; Zhu, Y. Rapid identification of a stripe rust resistant gene in a space-induced wheat mutant using specific locus amplified fragment (SLAF) sequencing. Sci. Rep. 2018, 8, 3086. [Google Scholar] [CrossRef]
- Tester, M.; Bacic, A. Abiotic stress tolerance in grasses. From model plants to crop plants. Plant Physiol. 2005, 137, 791–793. [Google Scholar] [CrossRef] [Green Version]
- Oecking, C.; Jaspert, N. Plant 14-3-3 proteins catch up with their mammalian orthologs. Curr. Opin. Plant Biol. 2009, 12, 760–765. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, X.; Chang, S.; Chu, Z.; Wang, H.; Han, S.; Wang, Y. Calcium-dependent protein kinase 21 phosphorylates 14-3-3 proteins in response to ABA signaling and salt stress in rice. Biochem. Biophys. Res. Commun. 2017, 493, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, H.; Huang, F.; Long, J.; Song, G.; Lin, W. The 14-3-3 protein GF14f negatively affects grain filling of inferior spikelets of rice (Oryza sativa L.). Plant J. 2019, 99, 344–358. [Google Scholar] [PubMed]
- Manosalva, P.M.; Bruce, M.; Leach, J.E. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance. Plant J. 2011, 68, 777–787. [Google Scholar] [CrossRef]
- Xu, W.; Shi, W. Expression profiling of the 14-3-3 gene family in response to salt stress and potassium and iron deficiencies in young tomato (Solanum lycopersicum) roots: Analysis by real-time RT-PCR. Ann. Bot. 2006, 98, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, H.; Zhou, S.; He, Y.; Luo, Q.; Zhang, F.; Qiu, D.; Feng, J.; Wei, Q.; Chen, L. Expression of TaGF14b, a 14-3-3 adaptor protein gene from wheat, enhances drought and salt tolerance in transgenic tobacco. Planta 2018, 248, 117–137. [Google Scholar] [CrossRef]
- Sun, X.; Luo, X.; Sun, M.; Chen, C.; Ding, X.; Wang, X.; Yang, S.; Yu, Q.; Jia, B.; Ji, W. A Glycine soja 14-3-3 protein GsGF14o participates in stomatal and root hair development and drought tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Dai, S.; Li, H.; Liu, A.; Liu, C.; Cheng, D.; Cao, X.; Chu, X.; Zhai, S.; Liu, J. Identification and Expression Analysis of Wheat TaGF14 Genes. Front. Genet. 2018, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- González, R.R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; Ex, F.V.; Pasha, A. The transcriptional landscape of polyploid wheat. Science 2018, 361, 6089. [Google Scholar] [CrossRef] [Green Version]
- De Lille, J.M.; Sehnke, P.C.; Ferl, R.J. The Arabidopsis 14-3-3 Family of Signaling Regulators. Plant Physiol. 2001, 126, 35–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.M.; Mysore, K.S. New dimensions for VIGS in plant functional genomics. Trends Plant Sci. 2011, 16, 656–665. [Google Scholar] [CrossRef]
- Chen, Y.; Ho, T.D.; Liu, L.; Lee, D.H.; Lee, C.; Chen, Y.; Lin, S.; Lu, C.; Yu, S. Sugar starvation-regulated MYBS2 and 14-3-3 protein interactions enhance plant growth, stress tolerance, and grain weight in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Dhaubhadel, S. 14-3-3 proteins act as scaffolds for GmMYB62 and GmMYB176 and regulate their intracellular localization in soybean. Plant Signal Behav. 2012, 7, 965–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Wang, S.; Xiang, W.; Yang, H.; Tahir, M.M.; Zheng, S.; An, N.; Han, M.; Zhao, C.; Zhang, D. Genome-wide identification of the 14-3-3 gene family and its participation in floral transition by interacting with TFL1/FT in apple. BMC Genom. 2021, 22, 41. [Google Scholar] [CrossRef]
- Ren, Y.R.; Zhao, Q.; Yang, Y.Y.; Zhang, T.E.; Wang, X.F.; You, C.X.; Hao, Y.J. The apple 14-3-3 protein MdGRF11 interacts with the BTB protein MdBT2 to regulate nitrate deficiency-induced anthocyanin accumulation. Hortic. Res. 2021, 8, 22. [Google Scholar] [CrossRef]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. 14-3-3 Proteins in Plant Hormone Signaling: Doing Several Things at Once. Front. Plant. Sci. 2018, 9, 297. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kong, Q.; Mantyla, J.J.; Yang, Y.; Ohlrogge, J.B.; Benning, C. 14-3-3 protein mediates plant seed oil biosynthesis through interaction with AtWRI1. Plant J. 2016, 88, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Lin, H.; Chen, S.; Becker, K.; Yang, Y.; Zhao, J.; Kudla, J.; Schumaker, K.S.; Guo, Y. Inhibition of the Arabidopsis salt overly sensitive pathway by 14-3-3 proteins. Plant Cell 2014, 26, 1166–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzinger, A.; Popowicz, G.M.; Joy, J.K.; Majumdar, S.; Holak, T.A.; Hermeking, H. The crystal structure of the non-liganded 14-3-3σ protein: Insights into determinants of isoform specific ligand binding and dimerization. Cell Res. 2005, 15, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Truong, A.B.; Masters, S.C.; Yang, H.; Fu, H. Role of the 14-3-3 C-terminal loop in ligand interaction. Proteins 2002, 49, 321–325. [Google Scholar] [CrossRef]
- Börnke, F. The variable C-terminus of 14-3-3 proteins mediates isoform-specific interaction with sucrose-phosphate synthase in the yeast two-hybrid system. J. Plant Physiol. 2005, 162, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Du, Y.; Jiang, L.; Liu, J. Molecular analysis and expression patterns of the 14-3-3 gene family from Oryza sativa. J. Biochem. Mol. Biol. 2007, 40, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, Q.; Sun, L.; He, Z. The rice 14-3-3 gene family and its involvement in responses to biotic and abiotic stress. DNA Res. 2006, 13, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.R.; Salinas, J.; Collinge, D.B. 14-3-3 proteins and the response to abiotic and biotic stress. Plant Mol. Biol. 2002, 50, 1031–1039. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Y.; Chen, L.; Wu, C.; Luo, Q.; Zhang, F.; Wei, Q.; Li, K.; Chang, J.; Yang, G. A Member of the 14-3-3 Gene Family in Brachypodium distachyon, BdGF14d, Confers Salt Tolerance in Transgenic Tobacco Plants. Front. Plant Sci. 2017, 8, 340. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Chen, L.; Dhaubhadel, S. 14-3-3 proteins regulate the intracellular localization of the transcriptional activator GmMYB176 and affect isoflavonoid synthesis in soybean. Plant J. 2012, 71, 239–250. [Google Scholar] [CrossRef]
- Finn, R.D.; Mistry, J.; SchusterBockler, B.; GriffithsJones, S.; Hollich, V.; Lassmann, T.; Moxon, S.; Marshall, M.; Khanna, A.; Durbin, R. Pfam: Clans, web tools and services. Nucleic Acids Res. 2006, D247–D251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, T.J.; Eddy, S.R. nhmmer: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Derbyshire, M.K.; Yamashita, R.A.; Bauer, M.A. NCBI’s Conserved Domain Database and Tools for Protein Domain Analysis. Curr. Protoc. Bioinform. 2020, 69. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 4i6, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modelling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools in the ExPASy Server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Ding, S.; Cai, Z.; Du, H.; Wang, H. Genome-Wide Analysis of TCP Family Genes in Zea mays L. Identified a Role for ZmTCP42 in Drought Tolerance. Int. J. Mol. Sci. 2019, 20, 2762. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, W202–W208. [Google Scholar] [CrossRef]
- Wang, G.; Wang, T.; Jia, Z.; Xuan, J.; Pan, D.; Guo, Z.; Zhang, J. Genome-Wide Bioinformatics Analysis of MAPK Gene Family in Kiwifruit (Actinidia Chinensis). Int. J. Mol. Sci. 2018, 19, 2510. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Yang, L.; He, Y.; Zhang, H.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and transcriptional expression analysis of superoxide dismutase (SOD) family in wheat (Triticum aestivum). PeerJ 2019, 7, 8062. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhu, X.; Shao, W.; Song, J.; Jiang, W.; He, Y.; Yin, J.; Ma, D.; Qiao, Y. Genome-Wide Mining of Wheat DUF966 Gene Family Provides New Insights into Salt Stress Responses. Front. Plant Sci. 2020, 11, 9838. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [Green Version]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.M.D.; Hand, T.W.; Han, S.; Gerner, M.Y.; Zaretsky, A.G.; Byrd, A.L.; Harrison, O.J.; Ortiz, A.M.; Quinones, M.; Trinchieri, G. Microbiota-Dependent Sequelae of Acute Infection Compromise Tissue-Specific Immunity. Cell 2015, 163, 354–366. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Geng, Y.; Liu, Y.; Chen, S.; Cao, S.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and characterization of SRO gene family in wheat: Molecular evolution and expression profiles during different stresses. Plant Physiol. Biochem. 2020, 154, 590–611. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Qi, T.; Yang, Q.; He, F.; Tan, C.; Ma, W.; Voegele, R.T.; Kang, Z.; Guo, J. Host-Induced Gene Silencing of the MAPKK Gene PsFUZ7 Confers Stable Resistance to Wheat Stripe Rust. Plant Physiol. 2017, 175, 1853–1863. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Jia, Y.; Shi, J.; Chen, C.; Ye, W.; Wang, Y.; Ma, W.; Qiao, Y. The WY domain in the Phytophthora effector PSR1 is required for infection and RNA silencing suppression activity. N. Phytol. 2019, 223, 839–852. [Google Scholar] [CrossRef]
- Chen, C.; He, B.; Liu, X.; Ma, X.; Liu, Y.; Yao, H.Y.; Zhang, P.; Yin, J.; Wei, X.; Koh, H.J. Pyrophosphate-fructose 6-phosphate 1-phosphotransferase (PFP1) regulates starch biosynthesis and seed development via heterotetramer formation in rice (Oryza sativa L). Plant Biotechnol. J. 2020, 18, 83–95. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence ID | ASa | Genomic Position | CDSb | Proteinc | MWd | pIe | Information |
|---|---|---|---|---|---|---|---|---|
| TaGRF1-A | TraesCS2A02G337300.1 | 2 | Chrf2:570782629–57078596 (−)g | 792 | 263 | 29.95 | 4.69 | TaGF14a# |
| TaGRF1-B | TraesCS2B02G344600.1 | 2 | Chr2:491180649–491184008 (+)h | 792 | 263 | 29.97 | 4.73 | TaGF14e# |
| TaGRF1-D | TraesCS2D02G325600.1 | 2 | Chr2:419039525–419043028 (+) | 792 | 263 | 29.95 | 4.69 | TaGF14k#, TaGF14r# |
| TaGRF2-A | TraesCS3A02G055600.1 | 2 | Chr3:32150045–32153332 (−) | 789 | 262 | 29.69 | 4.67 | TaGF14b# |
| TaGRF2-B | TraesCS3B02G068000.1 | 3 | Chr3:40243799–40247161 (−) | 849 | 282 | 31.93 | 4.98 | Identified in this study |
| TaGRF2-D | TraesCS3D02G055500.1 | 1 | Chr3:23061692–23065089 (−) | 789 | 262 | 29.75 | 4.71 | TaGF14m#, TaGF14s# |
| TaGRF3-A | TraesCS4A02G268700.1 | 2 | Chr4:580905848–580908549 (−) | 786 | 261 | 29.26 | 4.83 | TaGF14d# |
| TaGRF3-B | TraesCS4B02G045500.2 | 2 | Chr4:32805467–32808180 (+) | 786 | 261 | 29.26 | 4.83 | TaGF14h# |
| TaGRF3-D | TraesCS4D02G046400.2 | 2 | Chr4:21886194–21888840 (−) | 786 | 261 | 29.26 | 4.83 | TaGF14j# |
| TaGRF4-A | TraesCS4A02G151000.2 | 3 | Chr4:299599404–299602584 (−) | 813 | 270 | 29.90 | 4.76 | TaGF14c# |
| TaGRF4-B | TraesCS4B02G159900.1 | 3 | Chr4:310973441–310976945 (+) | 822 | 273 | 30.22 | 4.75 | TaGF14g# |
| TaGRF4-D | TraesCS4D02G155900.1 | 2 | Chr4:209095178–209098724 (+) | 801 | 266 | 29.49 | 4.76 | TaGF14n# |
| TaGRF5-B | TraesCS4B02G148900.1 | 2 | Chr4:217846208–217849422 (+) | 738 | 245 | 27.56 | 5.90 | TaGF14i# |
| TaGRF6-A | TraesCS7A02G295500.1 | 1 | Chr7:386346364–386357777 (−) | 780 | 259 | 28.70 | 4.80 | TaGF14o# |
| TaGRF6-B | TraesCS7B02G183800.1 | 1 | Chr7:292828452–292841187 (+) | 786 | 261 | 28.80 | 4.80 | Identified in this study |
| TaGRF6-D | TraesCS7D02G292400.1 | 1 | Chr7:356896682–356911158 (+) | 786 | 261 | 28.80 | 4.74 | TaGF14q#, TaGF14t# |
| TaGRF-like1 | TraesCS4A02G167100.1 | 1 | Chr4:412260859–412267803(+) | 735 | 244 | 27.25 | 6.32 | Identified in this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, W.; Chen, W.; Zhu, X.; Zhou, X.; Jin, Y.; Zhan, C.; Liu, G.; Liu, X.; Ma, D.; Qiao, Y. Genome-Wide Identification and Characterization of Wheat 14-3-3 Genes Unravels the Role of TaGRF6-A in Salt Stress Tolerance by Binding MYB Transcription Factor. Int. J. Mol. Sci. 2021, 22, 1904. https://doi.org/10.3390/ijms22041904
Shao W, Chen W, Zhu X, Zhou X, Jin Y, Zhan C, Liu G, Liu X, Ma D, Qiao Y. Genome-Wide Identification and Characterization of Wheat 14-3-3 Genes Unravels the Role of TaGRF6-A in Salt Stress Tolerance by Binding MYB Transcription Factor. International Journal of Molecular Sciences. 2021; 22(4):1904. https://doi.org/10.3390/ijms22041904
Chicago/Turabian StyleShao, Wenna, Wang Chen, Xiaoguo Zhu, Xiaoyi Zhou, Yingying Jin, Chuang Zhan, Gensen Liu, Xi Liu, Dongfang Ma, and Yongli Qiao. 2021. "Genome-Wide Identification and Characterization of Wheat 14-3-3 Genes Unravels the Role of TaGRF6-A in Salt Stress Tolerance by Binding MYB Transcription Factor" International Journal of Molecular Sciences 22, no. 4: 1904. https://doi.org/10.3390/ijms22041904