Genome-Wide Identification and Functional Characterization of the Cation Proton Antiporter (CPA) Family Related to Salt Stress Response in Radish (Raphanus sativus L.)


 and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Classification of RsCPA Members in Radish
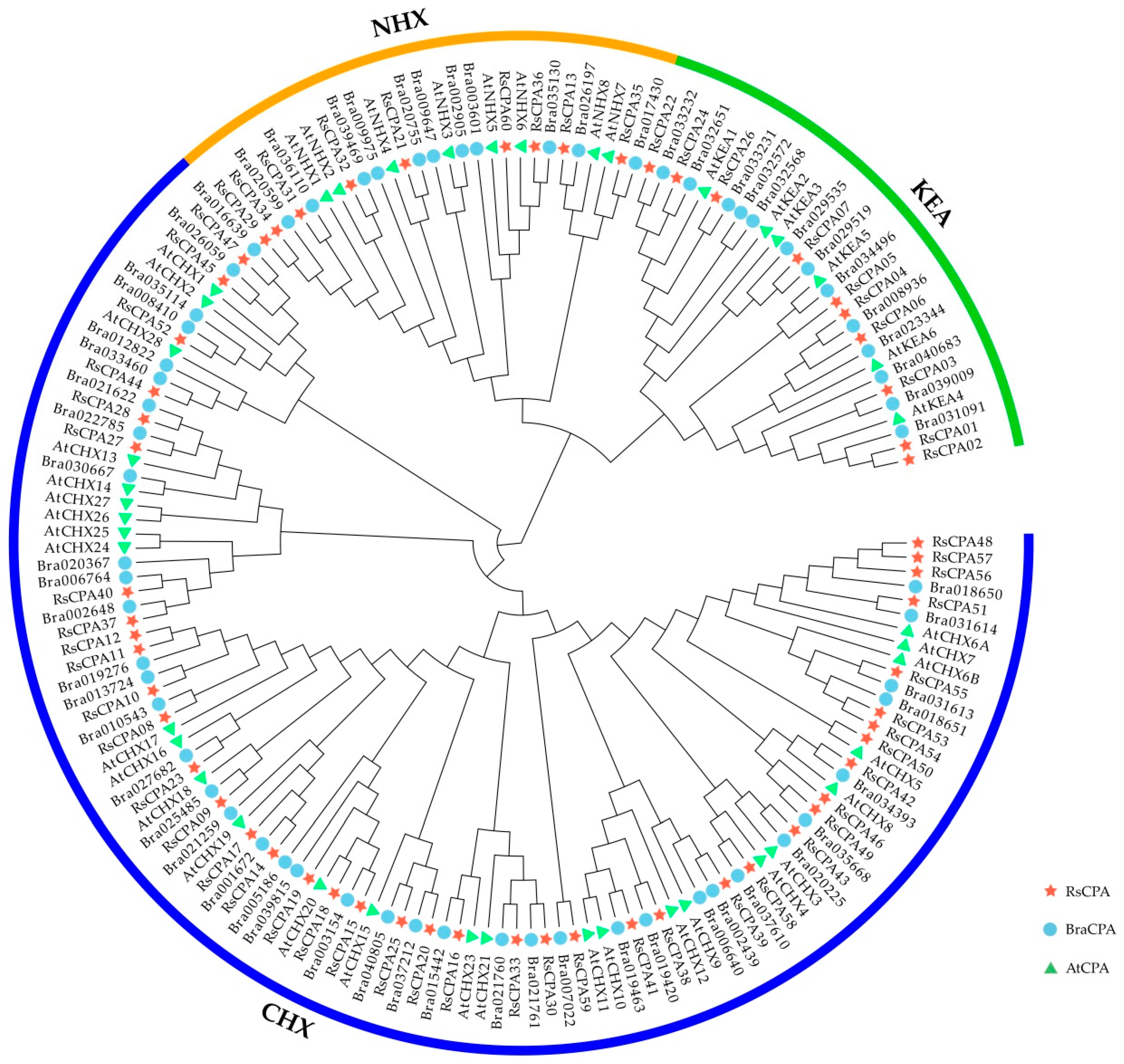
2.2. Phylogenetic Analysis of RsCPA Members
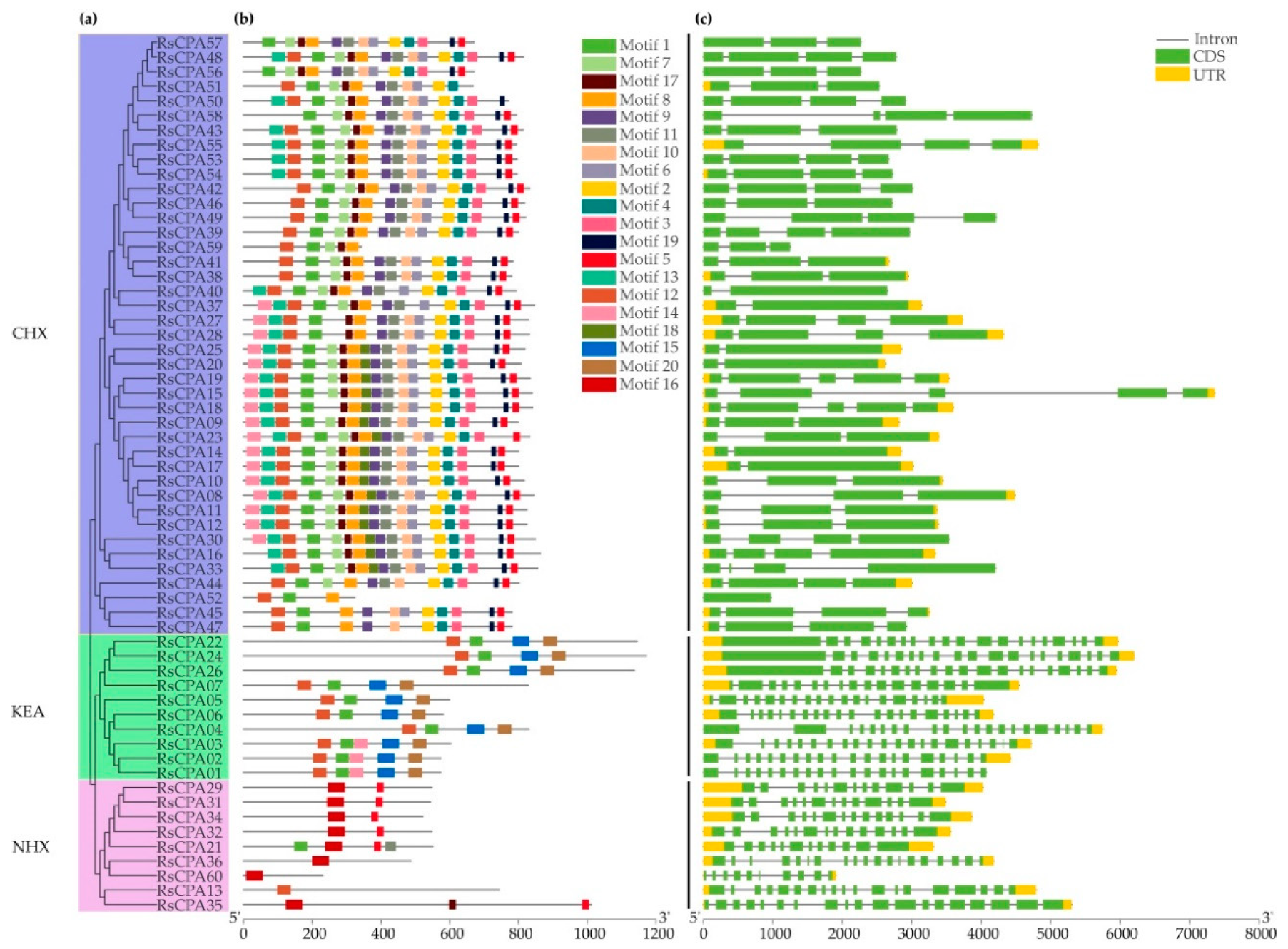
2.3. Gene Structure and Motif Composition Analysis
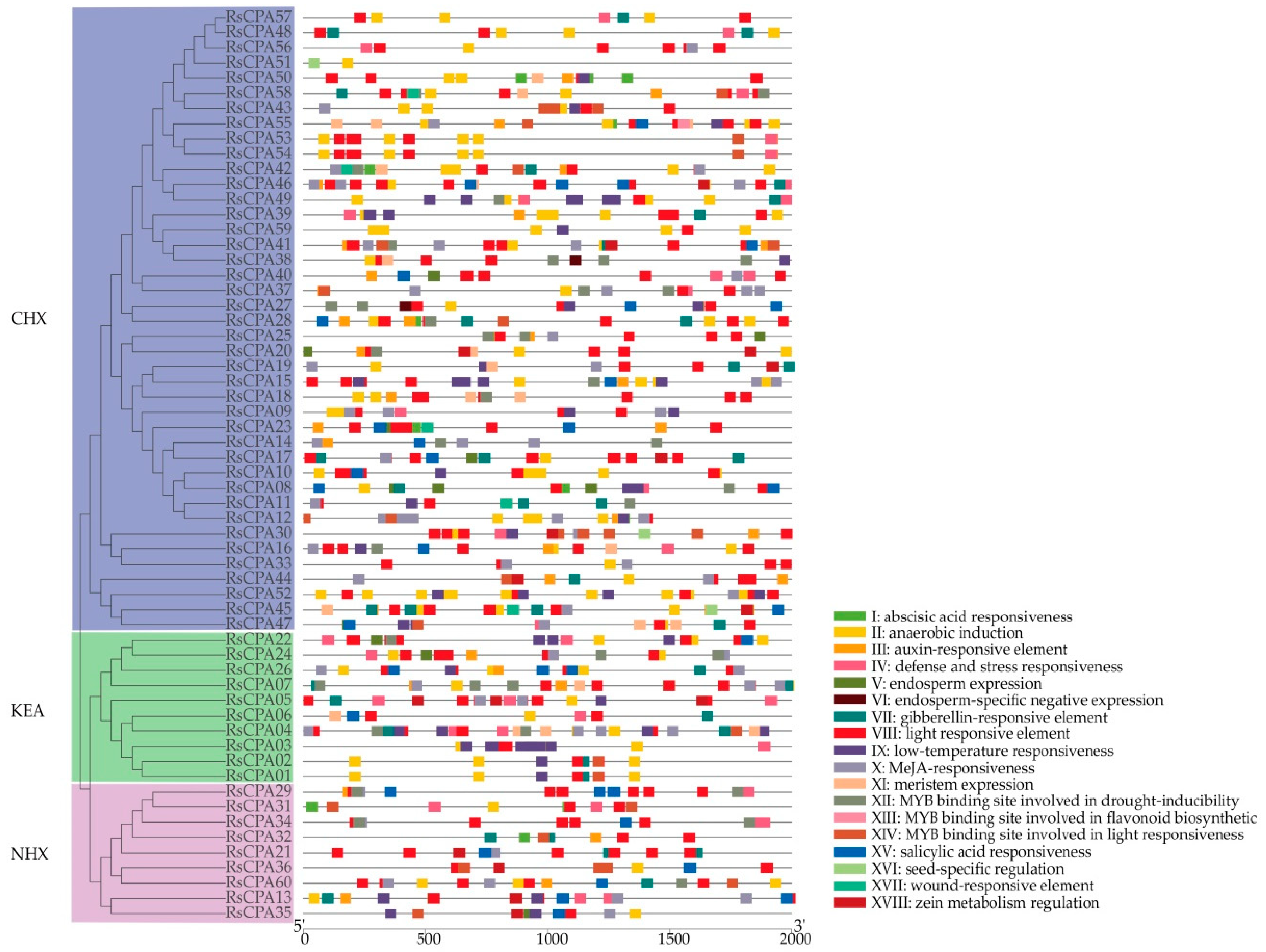
2.4. Promoter Elements and Transmembrane Region Analysis
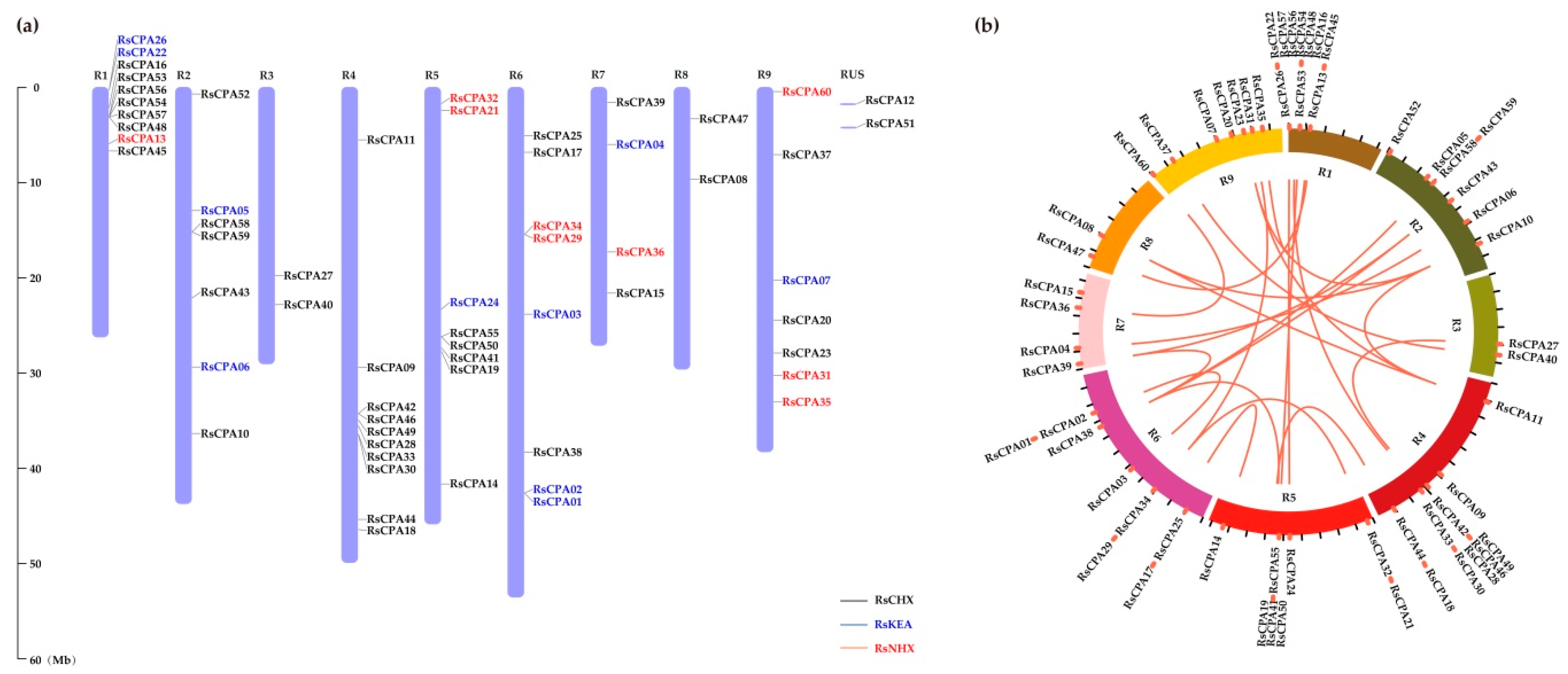
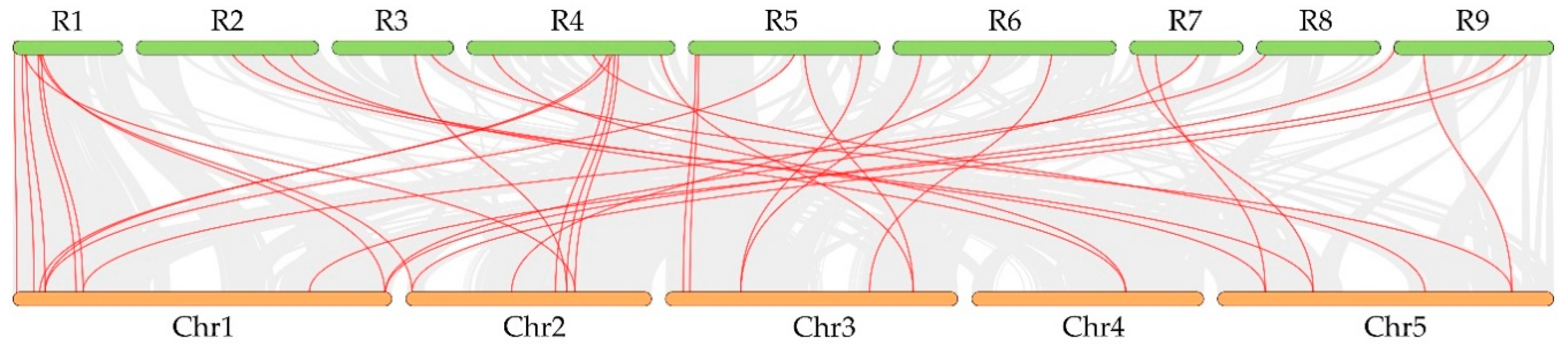
2.5. Chromosomal Localization and Gene Distribution Analysis
2.6. Evolution Analysis of the RsCPA Genes
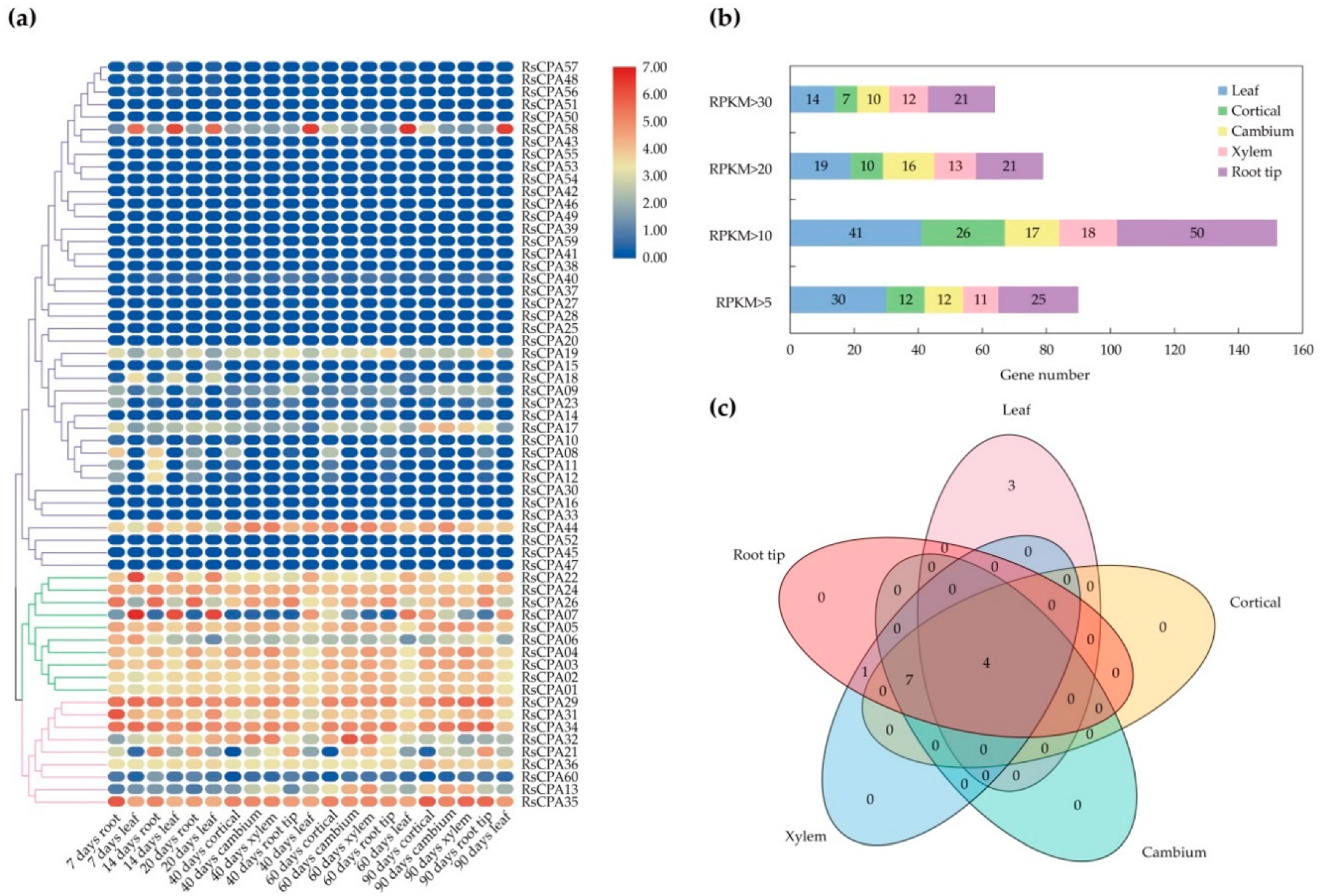
2.7. Spatial and Temporal Expression Profiles of RsCPA Genes
2.8. The RsCPA Genes Expression Levels under Abotic Stresses
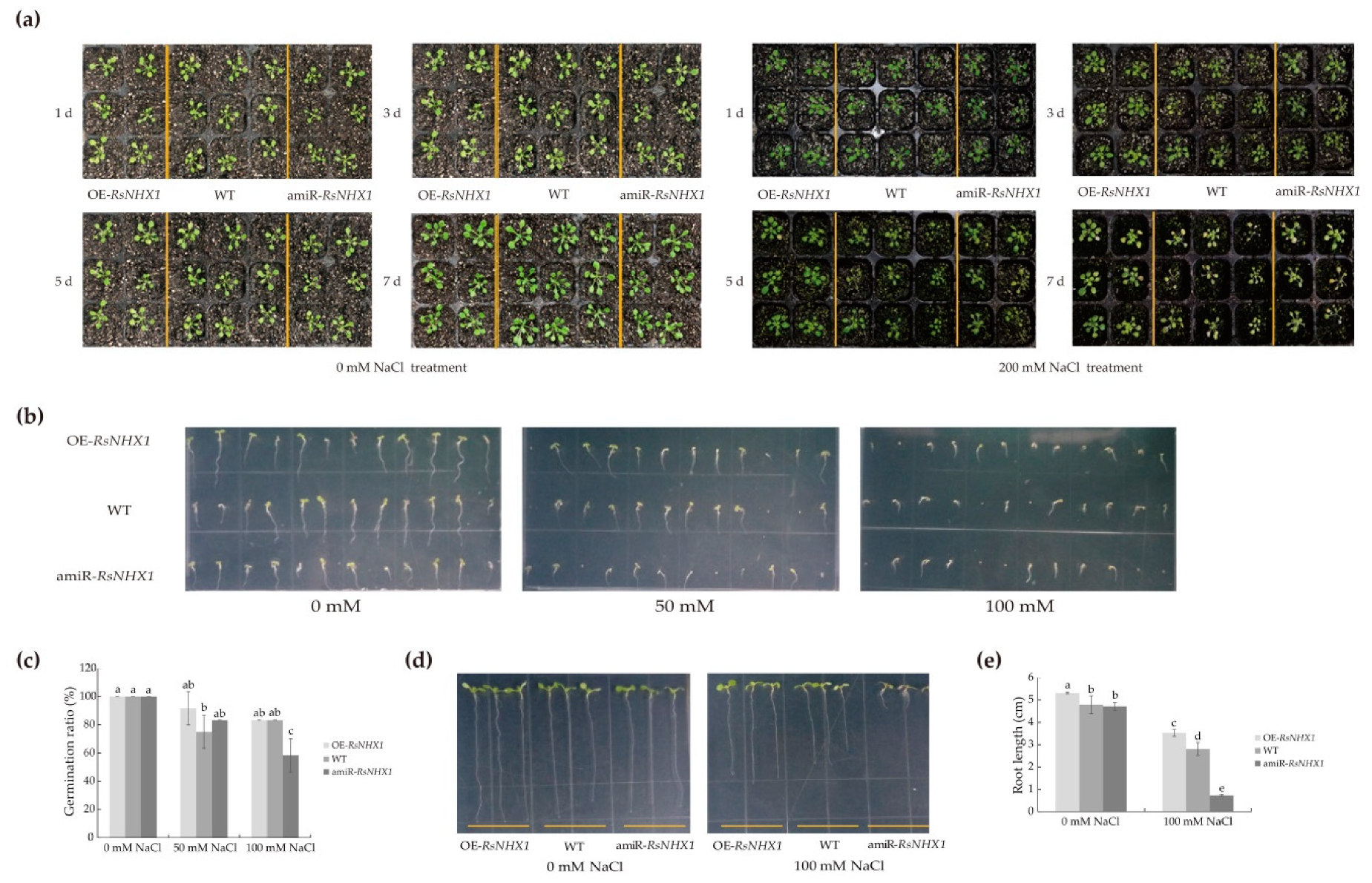
2.9. Ectopic Expression of the RsNHX1 Gene in Arabidopsis Can Influence Salt Tolerance
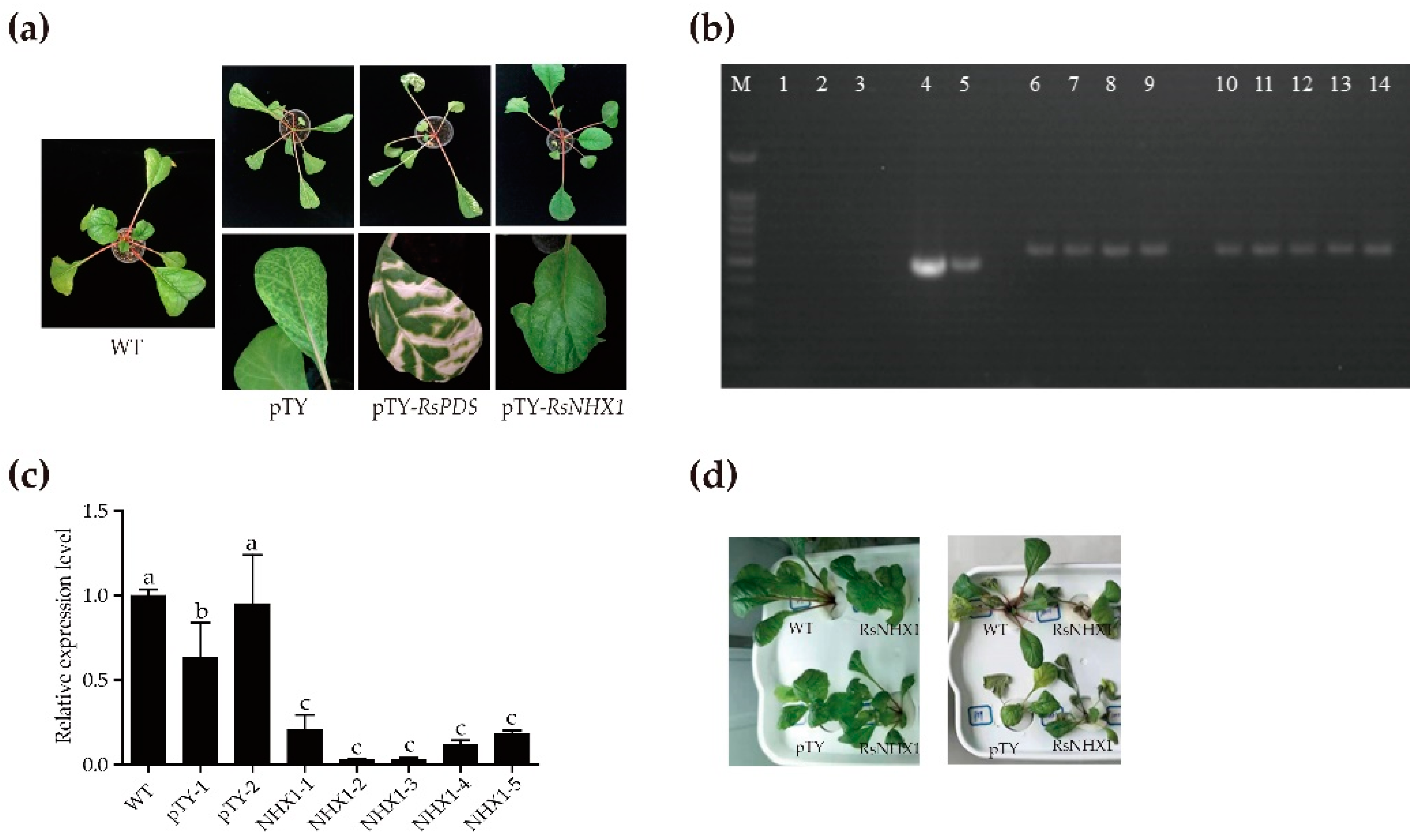
2.10. Functional Analysis of RsNHX1 in Radish Confirms that It Can Positively Regulates Salt Tolerance
3. Discussion
3.1. Genome-Wide Identification and Phylogenetic Analysis of CPA Genes in Radish
3.2. Evolutionary Characterization of the RsCPA Family
3.3. Roles of RsCPA Genes in Response to Different Abiotic Stresses
3.4. Potential Functions of RsNHXs Genes in Salt Stress
4. Materials and Methods
4.1. Sequence Collection, CPA Identification, and Phylogenetic Analysis
4.2. Gene Structure, Motif Composition, and Promoter Element Analysis
4.3. Synteny Analysis and Chromosomal Localization
4.4. Expression Analysis of RsCPAs Based on the RNA-Seq. Data
4.5. Plant Material, Salt Stress Treatment, and RT-qPCR Expression Analysis
4.6. Genetic Transformation and Generation of RsNHX1 Transgenic Lines in Arabidopsis
4.7. VIGS-Mediated Silencing of RsNHX1 in Radish
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | Amino Acids |
| ABA | Abscisic Acid |
| AtCPA | Arabidopsis thaliana Cation proton antiporter gene |
| BraCPA | Brassica rapa Cation proton antiporter gene |
| BRAD | Brassica database |
| Cd | Cadmium |
| Chr(s) | Chromosome(s) |
| CHX | Cation/H+ exchanger |
| CPA | Cation proton antiporter |
| Cr | Chromium |
| DD | Dispersed duplication |
| GA3 | Gibberellin A3 |
| GRAVY | Grand average of hydropathicity |
| HM | Heavy metal |
| HMM | Hidden Markov Model |
| KEA | K+ efflux antiporter |
| MCScanX | Multiple Collinearity Scan toolkit |
| MeJA | Methyl Jasmonate |
| MEME | Multiple Em for Motif Elicitation |
| MW | Molecular weight |
| NHX | Na+/H+ exchanger family |
| NJ | Neigbor-joining |
| ORF | Open reading frame |
| Pb | Lead |
| PD | Proximal duplicatio |
| pI | Theoretical isoelectric point |
| PlantDGD | Plant Duplicate Gene Database |
| RGD | Radish Genome Database |
| RPKM | Reads per kilobase per million reads |
| RsCPA | Raphanus sativus Cation proton antiporter gene |
| RT–qPCR | Real-Time Quantitative Polymerase Chain Reaction |
| TD | Tandem duplication |
| TMHMM | TransMembrane prediction was analyzed by Hidden Markov Models |
| TRD | Transposed duplication |
| TYMV | Turnip yellow mosaic virus |
| UTR | Untranslated region |
| VIGS | Virus-induced gene silencing |
| WGD | Whole-genome duplication |
| WT | Wide type |
References
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 16, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanroj, S.; Wang, G.Y.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and diversified gene families of monovalent cation/H+ antiporters from algae to flowering plants. Front. Plant Sci. 2012, 9, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.Y.; Yang, X.; Xia, X.; Yin, W.L. Comparative analysis of cation/proton antiporter superfamily in plants. Gene 2013, 521, 245–251. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Chen, Q.J.; Chai, M.F.; Lu, P.L.; Su, Z.; Qin, Z.X.; Chen, J.; Wang, X.C. AtNHX8, a member of the monovalent cation: Proton antiporter-1 family in Arabidopsis thaliana, encodes a putative Li+/H+ antiporter. Plant J. 2007, 49, 718–728. [Google Scholar] [CrossRef]
- Véry, A.A.; Sentenac, H. Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 2003, 54, 575–603. [Google Scholar] [CrossRef]
- Inês, S.P.; Sónia, N.; Melissa, M.P.; Isabel, A.A.; Margarida, M.O.; Michael, D.P. Different evolutionary histories of two cation/proton exchanger gene families in plants. BMC Plant Biol. 2013, 13, 97. [Google Scholar]
- Jia, Q.; Zheng, C.; Sun, S.; Amjad, H.; Liang, K.; Lin, W. The role of plant cation/proton antiporter gene family in salt tolerance. Biol. Plant. 2018, 62, 617–629. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary origins of eukaryotic sodium/proton exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Li, D.L.; Qiu, N.W.; Song, Y.G. The functions of plant cation/proton antiporters. Biol. Plant. 2018, 62, 421–427. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.M.; Sanders, D.; et al. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Li, Q.; Wang, H.Y.; Zhang, J.L.; Du, J.; Feng, H.M.; Blumwald, E.; Yu, L.; Xu, G.H. Two NHX-type transporters from Helianthus tuberosus improve the tolerance of rice to salinity and nutrient deficiency stress. Plant Biotechnol. J. 2017, 16, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, J.; Zhong, Y.; Cramer, G.R.; Cheng, Z.M.M. Genome-wide analysis of the cation/proton antiporter (CPA) super family genes in grapevine (Vitis vinifera L.). Plant Omics 2015, 8, 300–311. [Google Scholar]
- Zhou, H.S.; Qi, K.J.; Liu, X.; Yin, H.; Wang, P.; Chen, J.Q.; Wu, J.Y.; Zhang, S.L. Genome-wide identification and comparative analysis of the cation proton antiporters family in pear and four other Rosaceae species. Mol. Genet. Genom. 2016, 291, 1727–1742. [Google Scholar] [CrossRef]
- Banjara, M.; Zhu, L.F.; Shen, G.X.; Payton, P.; Zhang, H. Expression of an Arabidopsis sodium/proton antiporter gene (AtNHX1) in peanut to improve salt tolerance. Plant Biotechnol. Rep. 2012, 6, 59–67. [Google Scholar] [CrossRef]
- Bassil, E.; Tajima, H.; Liang, Y.C.; Ohto, M.A.; Ushijima, K.; Nakano, R.; Esumi, T.; Coku, A.; Belmonte, M.; Blumwald, E. The Arabidopsis Na+/H+ antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell 2011, 23, 3482–3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassil, E.; Ohto, M.; Esumi, T.; Tajima, H.; Zhu, Z.; Cagnac, O.; Belmonte, M.; Peleg, Z.; Yamaguchi, T.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 2011, 23, 224–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sze, H.; Padmanaban, S.; Cellier, F.; Honys, D.; Cheng, N.H.; Bock, K.W.; Conéjéro, G.; Li, X.Y.; Twell, D.; Ward, J.M.; et al. Expression patterns of a novel AtCHX gene family highlight potential roles in osmotic adjustment and K+ homeostasis in pollen development. Plant Physiol. 2004, 136, 2532–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [Green Version]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Yokoi, S.; Quintero, F.J.; Cubero, B.; Ruiz, M.T.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 2002, 30, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.T.; Liu, H.; Gao, X.S.; Zhang, H.X. Knock-out of Arabidopsis AtNHX4 gene enhances tolerance to salt stress. Biochem. Biophys. Res. Commun. 2009, 382, 637–641. [Google Scholar] [CrossRef]
- Mishra, S.; Alavilli, H.; Lee, B.; Panda, S.K.; Sahoo, L. Cloning and characterization of a novel vacuolar Na+/H+ antiporter gene (VuNHX1) from drought hardy legume, cowpea for salt tolerance. Plant Cell Tissue Organ 2015, 120, 19–33. [Google Scholar] [CrossRef]
- Sun, M.H.; Ma, Q.J.; Liu, X.; Zhu, X.P.; Hu, D.G.; Hao, Y.J. Molecular cloning and functional characterization of MdNHX1 reveals its involvement in salt tolerance in apple calli and Arabidopsis. Sci. Hortic. 2017, 215, 126–133. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, C.; Hu, Y.; Zhu, X.P.; Hu, D.G.; Hao, Y.J. The novel Na+/H+ antiporter gene SpNHX1 from Sesuvium portulacastrum confers enhanced salt tolerance to transgenic yeast. Acta Physiol. Plant. 2018, 40, 61. [Google Scholar] [CrossRef]
- Zhang, W.W.; Meng, J.J.; Xing, J.Y.; Yang, S.; Guo, F.; Li, X.G.; Wan, S.B. The K+/H+ antiporter AhNHX1 improved tobacco tolerance to NaCl stress by enhancing K+ retention. J. Plant Biol. 2017, 60, 259–267. [Google Scholar] [CrossRef]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellier, F.; Conéjéro, G.; Ricaud, L.; Luu, D.T.; Lepetit, M.; Gosti, F.; Casse, F. Characterization of AtCHX17, a member of the cation/H+ exchangers, CHX family, from Arabidopsis thaliana suggests a role in K+ homeostasis. Plant J. 2004, 39, 834–846. [Google Scholar] [CrossRef]
- Kunz, H.H.; Gierth, M.; Herdean, A.; Satoh-Cruzd, M.; Kramerd, D.M.; Speteac, C.; Schroeder, J.I. Plastidial transporters KEA-1, -2, and-3 are essential for chloroplast osmoregulation, integrity, and pH regulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 7480–7485. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jun, L.; Shi, W.M.; Yan, H.; Han, L. Identification and localized expression of putative K+/H+ antiporter genes in Arabidopsis. Acta Physiol. Plant. 2015, 37, 101. [Google Scholar]
- Sun, X.C.; Xu, L.; Wang, Y.; Yu, R.G.; Zhu, X.W.; Luo, X.B.; Gong, Y.Q.; Wang, R.H.; Limera, C.; Zhang, K.Y.; et al. Identification of novel and salt-responsive miRNAs to explore miRNA-mediated regulatory network of salt stress response in radish (Raphanus sativus L.). BMC Genom. 2015, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.C.; Wang, Y.; Xu, L.; Li, C.; Zhang, W.; Luo, X.B.; Jiang, H.Y.; Liu, L.W. Unraveling the root proteome changes and its relationship to molecular mechanism underlying salt stress response in radish (Raphanus sativus L.). Front. Plant Sci. 2017, 8, 1192. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.M.; Kim, N.; Ahn, B.O.; Oh, M.; Chung, W.H.; Chung, H.; Jeong, S.; Lim, K.B.; Hwang, Y.J.; Kim, G.B.; et al. Elucidating the triplicated ancestral genome structure of radish based on chromosome-level comparison with the Brassica genomes. Theor. Appl. Genet. 2016, 129, 1357–1372. [Google Scholar] [CrossRef]
- De, G.A.; Lanave, C.; Saccone, C. Genome duplication and gene-family evolution: The case of three OXPHOS gene families. Gene 2008, 421, 1–6. [Google Scholar]
- Airoldi, C.A.; Davies, B. Gene duplication and the evolution of plant MADS-box transcription factors. J. Genet. Genom. 2012, 39, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.J.; Xu, L.; Wang, Y.; Cheng, W.W.; Luo, X.B.; Xie, Y.; Fan, L.X.; Liu, L.W. Genome-wide characterization and evolutionary analysis of heat shock transcription factors (HSFs) to reveal their potential role under abiotic stresses in radish (Raphanus sativus L.). BMC Genom. 2019, 20, 772. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Lu, Z.; Wei, H.; Zhang, J.; Yang, X.; Wu, A.; Ma, L.; Kang, M.; Lu, J.; Wang, H.; et al. Genome-Wide identification and expression analysis of the NHX (Sodium/Hydrogen Antiporter) gene family in cotton. Front. Genet. 2020, 11, 964. [Google Scholar] [CrossRef]
- Fan, L.X.; Xu, L.; Wang, Y.; Tang, M.J.; Liu, L.W. Genome- and Transcriptome-wide characterization of bZIP gene family identifies potential members involved in abiotic stress response and anthocyanin biosynthesis in radish (Raphanus sativus L.). Int. J. Mol. Sci. 2019, 20, 6334. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.T.; Chen, X.; Wu, B.Y.; Yuan, X.X.; Zhang, H.M.; Cui, X.Y.; Liu, X.Q. Whole-Genome identification and expression analysis of KEA and NHX antiporter family under abiotic stress in soybean. J. Integr. Agric. 2015, 14, 1171–1183. [Google Scholar] [CrossRef]
- Jha, A.; Joshi, M.; Yadav, N.S.; Agarwal, P.; Jha, B. Cloning and characterization of the Salicornia brachiata Na+/H+ antiporter gene SbNHX1 and its expression by abiotic stress. Mol. Biol. Rep. 2011, 38, 1965–1973. [Google Scholar] [CrossRef]
- Hamed, A.; Hamid, R.K.; Hadi, H.; Iraj, T. Assessment of the vacuolar Na+/H+ antiporter (NHX1) transcriptional changes in Leptochloa fusca L. in response to salt and cadmium stresses. Mol. Biol. Res. Commun. 2015, 4, 133–142. [Google Scholar]
- Sharma, H.; Taneja, M.; Upadhyay, S.K. Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics 2020, 112, 356–370. [Google Scholar] [CrossRef]
- Yang, L.; Han, Y.J.; Wu, D.; Yong, W.; Liu, M.M.; Wang, S.T.; Liu, W.X.; Lu, M.Y.; Wei, Y.; Sun, J.S. Salt and cadmium stress tolerance caused by overexpression of the Glycine Max Na+/H+ antiporter (GmNHX1) gene in duckweed (Lemna turionifera 5511). Aquat. Toxicol. 2017, 192, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wei, Y.; Li, N.; Zeng, J.Y.; Han, Y.J.; Zuo, Z.J.; Wang, S.T.; Zhu, Y.R.; Zhang, Y.; Sun, J.S.; et al. Declined cadmium accumulation in Na+/H+ antiporter (NHX1) transgenic duckweed under cadmium stress. Ecotox. Environ. Saf. 2019, 182, 109397. [Google Scholar] [CrossRef] [PubMed]
- Akram, U.; Song, Y.H.; Liang, C.Z.; Abid, M.A.; Askari, M.; Myat, A.A.; Abbas, M.; Malik, W.; Ali, Z.; Guo, S.D.; et al. Genome-wide characterization and expression analysis of NHX gene family under salinity stress in Gossypium barbadense and its comparison with Gossypium Hirsutum. Genes 2020, 11, 803. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Aquib, A.; Anand, R. Analysis of physiological traits and expression of NHX and SOS3 genes in bread wheat (Triticum aestivum L.) under salinity stress. J. Pharmacogn. Phytochem. 2020, 9, 362–366. [Google Scholar]
- Ma, W.; Ren, Z.; Zhou, Y.; Zhao, J.; Zhang, F.; Feng, J.; Liu, W.; Ma, X. Genome-wide identification of the Gossypium hirsutum NHX genes reveals that the endosomal-type GhNHX4A is critical for the salt tolerance of cotton. Int. J. Mol. Sci. 2020, 21, 7712. [Google Scholar] [CrossRef]
- Sun, T.J.; Fan, L.; Yang, J.; Cao, R.Z.; Yang, C.Y.; Zhang, J.; Wang, D. A glycine max sodium/hydrogen exchanger enhances salt tolerance through maintaining higher Na+ efflux rate and K+/Na+ ratio in Arabidopsis. BMC Plant Biol. 2019, 19, 469. [Google Scholar] [CrossRef] [Green Version]
- Mushke, R.; Rajesh, Y.; Kirti, P.B. Improved salinity tolerance and growth performance in transgenic sunflower plants via ectopic expression of a wheat antiporter gene (TaNHX2). Mol. Biol. Rep. 2019, 46, 5941–5953. [Google Scholar] [CrossRef] [PubMed]
- Lamesch, P.; Berardini, T.Z.; Li, D.H.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M.; et al. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.H.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Liu, S.Y.; Wu, J.; Fang, L.; Sun, S.L.; Liu, B.; Li, P.X.; Hua, W.; Wang, X.W. BRAD, the genetics and genomics database for Brassica plants. BMC Plant. Biol. 2011, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, S.J.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L.L. The Pfam protein families database. Nucleic Acids Res. 2008, 32, D138. [Google Scholar]
- Sun, Y.N.; Buhler, J. Designing patterns for profile HMM search. Bioinformatics 2007, 23, e36. [Google Scholar] [CrossRef]
- Eddy, S.R.; Pearson, W.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Anderson, J.B.; Cherukuri, P.F.; DeWweese-Scott, C.; Geer, L.Y.; Gwadz, M.; He, S.Q.; Hurwitz, D.I.; Jackson, J.D.; Ke, Z.X. CDD: A Conserved Domain Database for protein classification. Nucleic Acids Res. 2005, 33, D192–D196. [Google Scholar] [CrossRef]
- Zdobnov, E.M.; Apweiler, R. InterProScan—An integration platform for the signature-recognition methods in InterPro. Bioinformatics 2001, 17, 847–848. [Google Scholar] [CrossRef] [Green Version]
- Elisabeth, G.; Alexandre, G.; Christine, H.; Ivan, I.; Ron, D.A.; Amos, B. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Olivier, G.; Mike, S. Neighbor-Joining Revealed. Mol. Biol. Evol. 2006, 23, 1997–2000. [Google Scholar]
- Zhang, H.K.; Gao, S.H.; Lercher, M.J.; Hu, S.N.; Chen, W.H. EvolView, an online tool for visualizing, annotating and managing phylogenetic trees. Nucleic Acids Res. 2012, 40, 569–572. [Google Scholar] [CrossRef]
- Chen, C.J.; Xia, R.; Chen, H.; He, Y.H. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Bailey, T.L. MEME-ChIP: Motif analysis of large DNA datasets. Bioinformatics 2011, 27, 1696–1697. [Google Scholar]
- Stephane, R.; Patrice, D.; Marc, V.M.; Pierre, R. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar]
- Chen, Y.J.; Yu, P.; Luo, J.C.; Jiang, Y. Secreted protein prediction system combining CJ-SPHMM, TMHMM, and PSORT. Mamm. Genome 2003, 14, 859–865. [Google Scholar] [CrossRef]
- Damian, S.; Andrea, F.; Michael, K.; Milan, S.; Alexander, R.; Pablo, M.; Tobias, D.; Manuel, S.; Jean, M.; Peer, B. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, 561–568. [Google Scholar]
- Qiao, X.; Li, Q.H.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.L.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization-diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Tang, H.B.; Debarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, Y.; Shimomura, M.; Komatsu, K.; Namiki, N.; Shibata-Hatta, M.; Imai, M.; Katayose, Y.; Mukai, Y.; Kanamori, H.; Kurita, K.; et al. The radish genome and comprehensive gene expression profile of tuberous root formation and development. Sci. Rep. 2015, 5, 10835. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, Y.; Liu, W.; Wang, J.; Zhu, X.W.; Zhang, K.Y.; Yu, R.G.; Wang, R.H.; Xie, Y.; Zhang, W.; et al. De novo sequencing of root transcriptome reveals complex cadmium-responsive regulatory networks in radish (Raphanus sativus L.). Plant Sci. 2015, 236, 313–323. [Google Scholar] [CrossRef]
- Xie, Y.; Ye, S.; Wang, Y.; Xu, L.; Zhu, X.W.; Yang, J.; Feng, H.Y.; Yu, R.G.; Karanja, B.; Gong, Y.Q.; et al. Transcriptome-based gene profiling provides novel insights into the characteristics of radish root response to Cr stress with next-generation sequencing. Front. Plant Sci. 2015, 6, 202. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Chen, Y.L.; Shen, H.; Gong, Y.Q.; Limera, C.; Liu, L.W. Transcriptome profiling of radish (Raphanus sativus L.) root and identification of genes involved in response to Lead (Pb) stress with next generation sequencing. PLoS ONE 2013, 8, e66539. [Google Scholar] [CrossRef]
- Wang, R.H.; Mei, Y.; Xu, L.; Zhu, X.W.; Wang, Y.; Guo, J.; Liu, L.W. Genome-wide characterization of differentially expressed genes provides insights into regulatory network of heat stress response in radish (Raphanus sativus L.). Funct. Integr. Genom. 2018, 18, 225–239. [Google Scholar] [CrossRef]
- Sun, X.C.; Xu, L.; Wang, Y.; Luo, X.B.; Zhu, X.W.; Kinuthia, K.B.; Nie, S.S.; Feng, H.Y.; Li, C.; Liu, L.W. Transcriptome-based gene expression profiling identifies differentially expressed genes critical for salt stress response in radish (Raphanus sativus L.). Plant Cell Rep. 2016, 35, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, Y.; Zhai, L.L.; Xu, Y.Y.; Wang, L.J.; Zhu, X.W.; Gong, Y.Q.; Yu, R.G.; Limera, C.; Liu, L.W. Genome-wide identification and characterization of cadmium-responsive microRNAs and their target genes in radish (Raphanus sativus L.) roots. J. Exp. Bot. 2013, 14, 4271–4287. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yu, J.; Yang, X.D.; Wang, Q.; Gao, L.W.; Yang, Y.; Xiao, D.; Liu, T.K.; Li, Y.; Hou, X.L.; Zhang, C.W. Efficient virus-induced gene silencing in Brassica rapa using a turnip yellow mosaic virus vector. Biol. Plant. 2018, 62, 826–834. [Google Scholar] [CrossRef]
- Topping, J.F.; Wei, W.B.; Clarke, M.C.; Muskett, P.; Lindsey, K. Agrobacterium-mediated transformation of Arabidopsis thaliana: Application in T-DNA tagging. Methods Mol. Biol. 1995, 49, 63–76. [Google Scholar]
- Schwab, R.; Ossowski, S.; Riester, M.; Warthmann, N.; Weigel, D. Highly specific gene silencing by artificial microRNAs in Arabidopsis. Plant Cell 2006, 18, 1121–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 2010, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.R.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 2, 641–646. [Google Scholar] [CrossRef]
- Li, M.R.; Lin, X.J.; Li, H.Q.; Pan, X.P.; Wu, G.J. Overexpression of AtNHX5 improves tolerance to both salt and water stress in rice (Oryza sativa L.). Plant Cell Tissue Org. 2011, 107, 283–293. [Google Scholar] [CrossRef]
- Simmonds, D.H.; Donaldson, P.A. Genotype screening for proliferative embryogenesis and biolistic transformation of short-season soybean genotypes. Plant Cell Rep. 2000, 19, 485–490. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | CHX Number | NHX Number | KEA Number | Total Gene Number |
|---|---|---|---|---|
| Arabidopsis thaliana | 28 | 8 | 6 | 42 |
| Raphanus sativus | 41 | 9 | 10 | 60 |
| Brassica rapa | 40 | 11 | 13 | 64 |
| Pyrus bretschneideri | 27 | 14 | 12 | 53 |
| Malus domestica | 42 | 12 | 7 | 61 |
| Prunus persica | 26 | 6 | 5 | 37 |
| Fragaria vesca | 24 | 6 | 5 | 35 |
| Prunus mume | 23 | 7 | 4 | 34 |
| Vitis vinifera | 17 | 8 | 4 | 29 |
| Oryza sativa | 18 | 8 | 4 | 30 |
| Zea mays | 16 | 11 | 6 | 33 |
| Sorghum bicolor | 17 | 7 | 4 | 28 |
| Selaginella | 3 | 7 | 4 | 14 |
| Ostreococcus | 0 | 6 | 4 | 10 |
| Chlorophyta reinhardtii | 0 | 9 | 3 | 12 |
| Physcomitrella patens | 5 | 10 | 7 | 22 |
| Populus trichocarpa | 29 | 8 | 7 | 44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Ying, J.; Zhang, Y.; Xu, L.; Zhang, W.; Ni, M.; Zhu, Y.; Liu, L. Genome-Wide Identification and Functional Characterization of the Cation Proton Antiporter (CPA) Family Related to Salt Stress Response in Radish (Raphanus sativus L.). Int. J. Mol. Sci. 2020, 21, 8262. https://doi.org/10.3390/ijms21218262
Wang Y, Ying J, Zhang Y, Xu L, Zhang W, Ni M, Zhu Y, Liu L. Genome-Wide Identification and Functional Characterization of the Cation Proton Antiporter (CPA) Family Related to Salt Stress Response in Radish (Raphanus sativus L.). International Journal of Molecular Sciences. 2020; 21(21):8262. https://doi.org/10.3390/ijms21218262
Chicago/Turabian StyleWang, Yan, Jiali Ying, Yang Zhang, Liang Xu, Wanting Zhang, Meng Ni, Yuelin Zhu, and Liwang Liu. 2020. "Genome-Wide Identification and Functional Characterization of the Cation Proton Antiporter (CPA) Family Related to Salt Stress Response in Radish (Raphanus sativus L.)" International Journal of Molecular Sciences 21, no. 21: 8262. https://doi.org/10.3390/ijms21218262