
Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize?

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
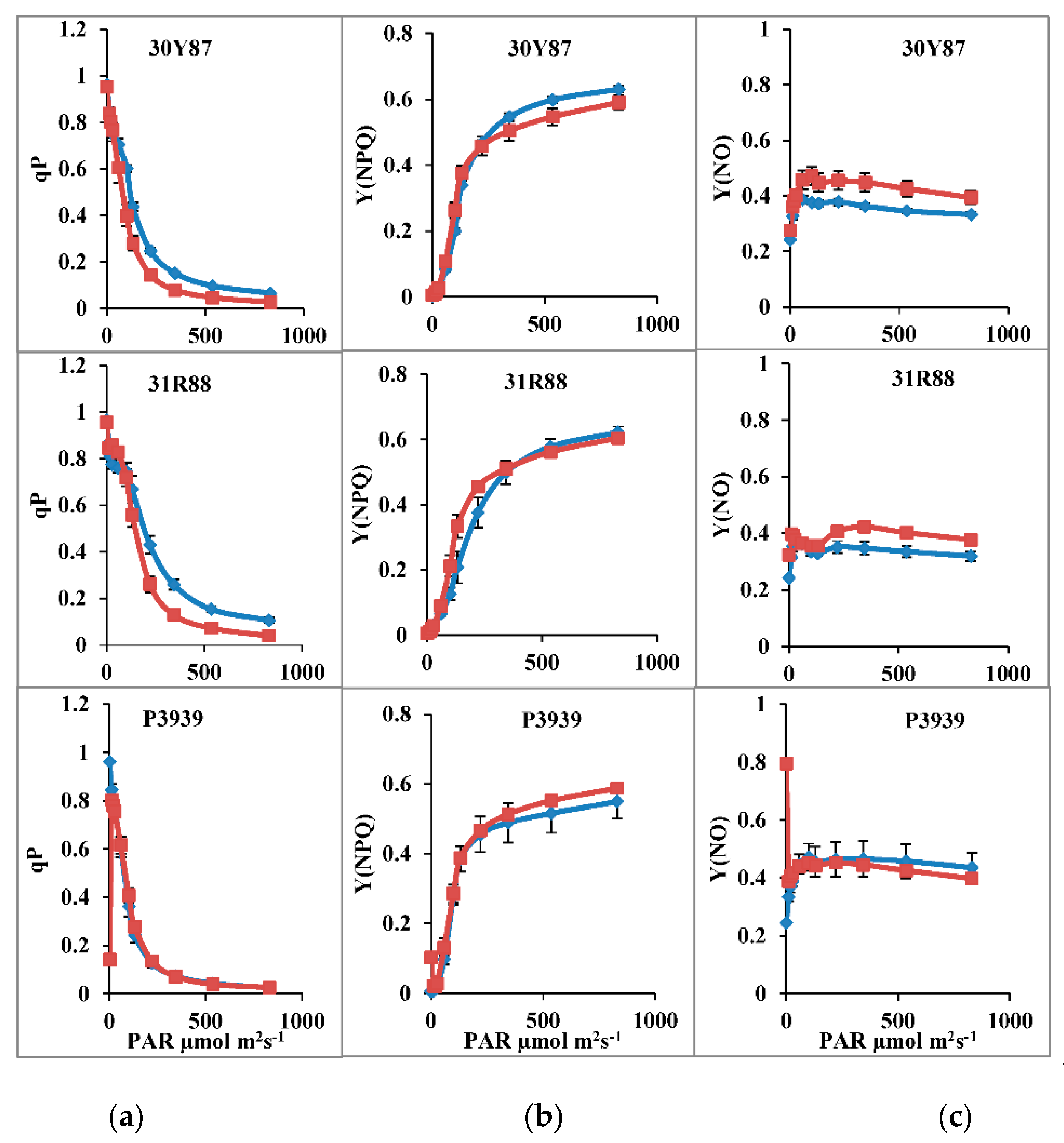
2.1. Light-Response Curve
2.1.1. Measurement of Activity of PSII, ETR(II), qP
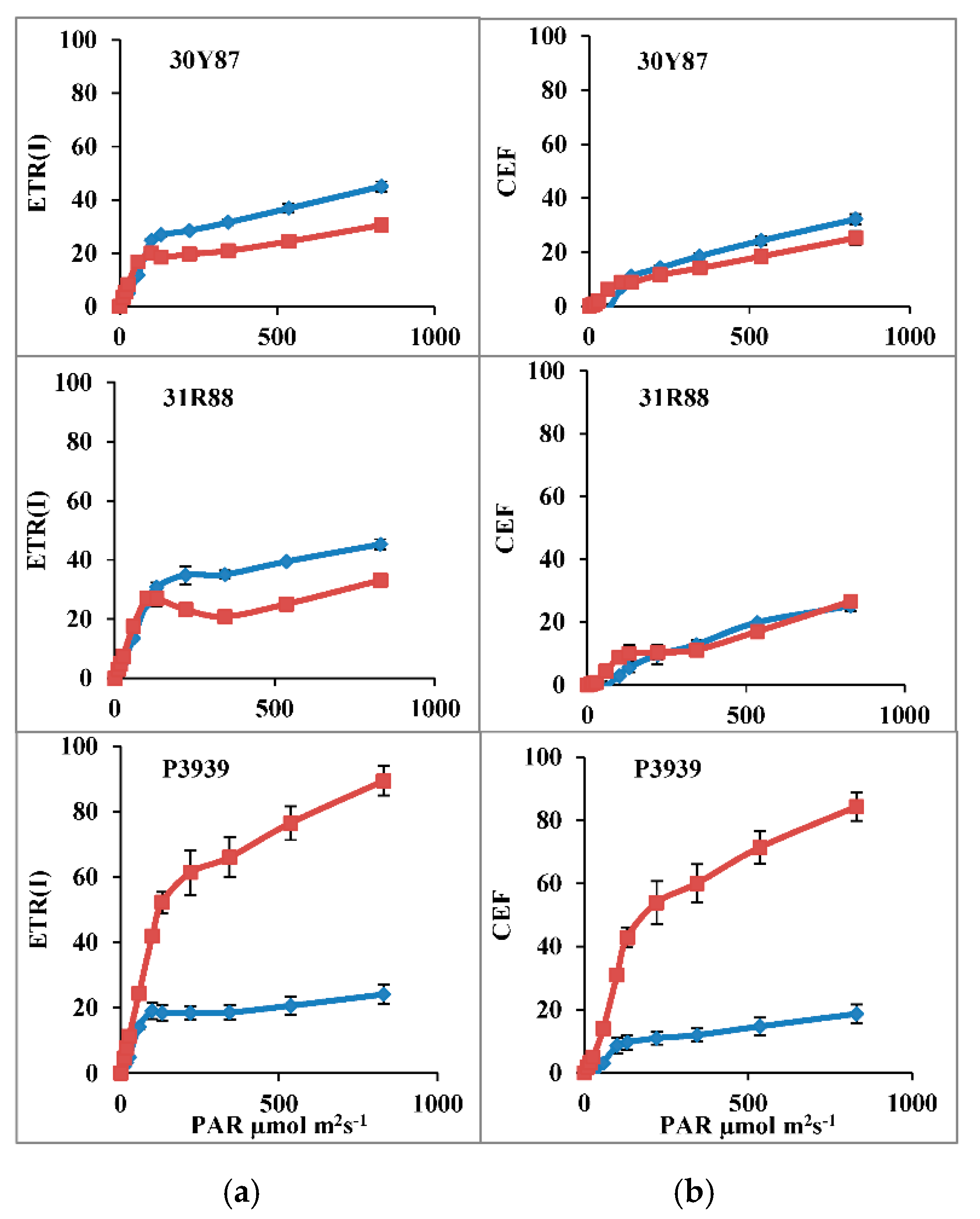
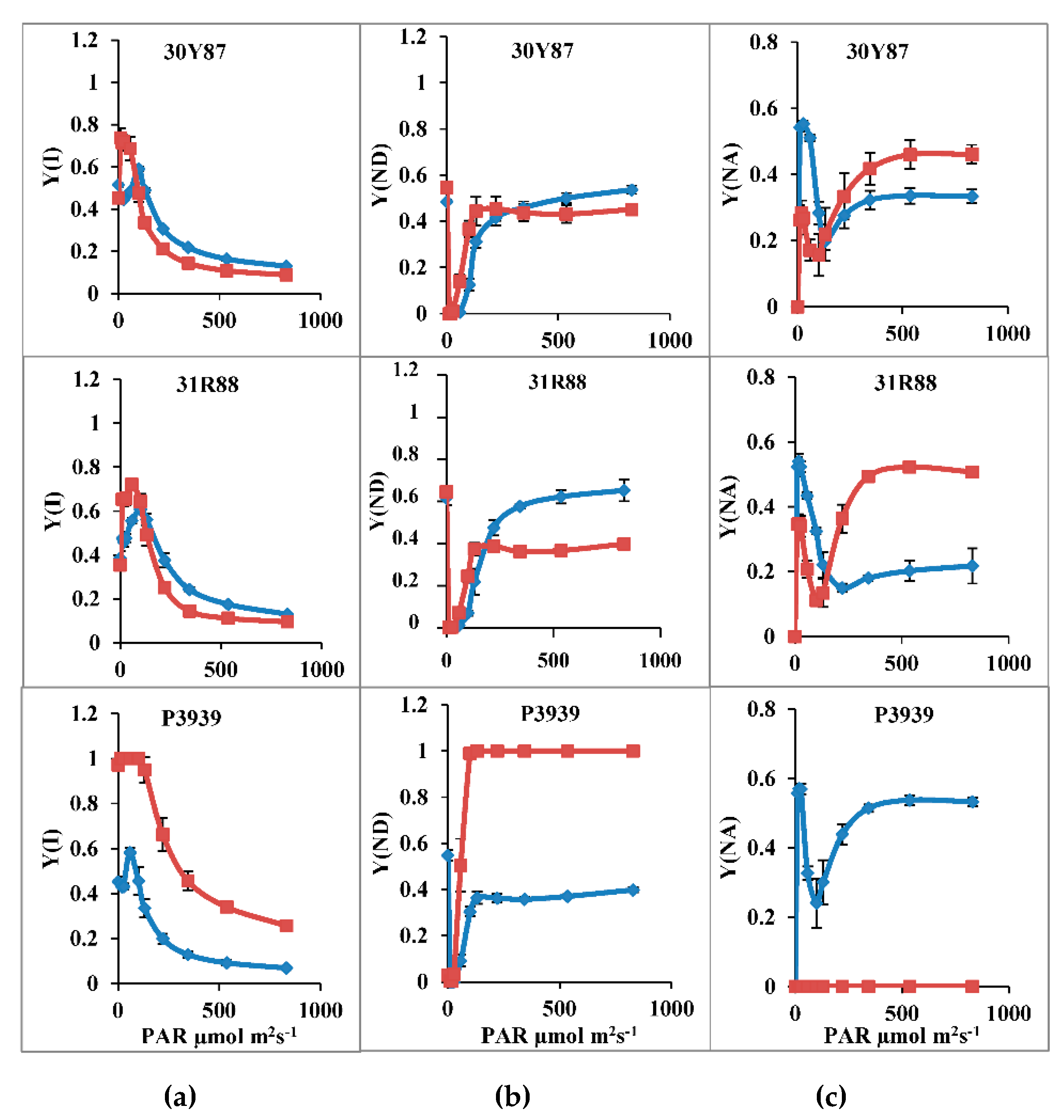
2.1.2. Measurement of Activity of PSI, ETR(I), Y(ND), Y(NA) and CEF
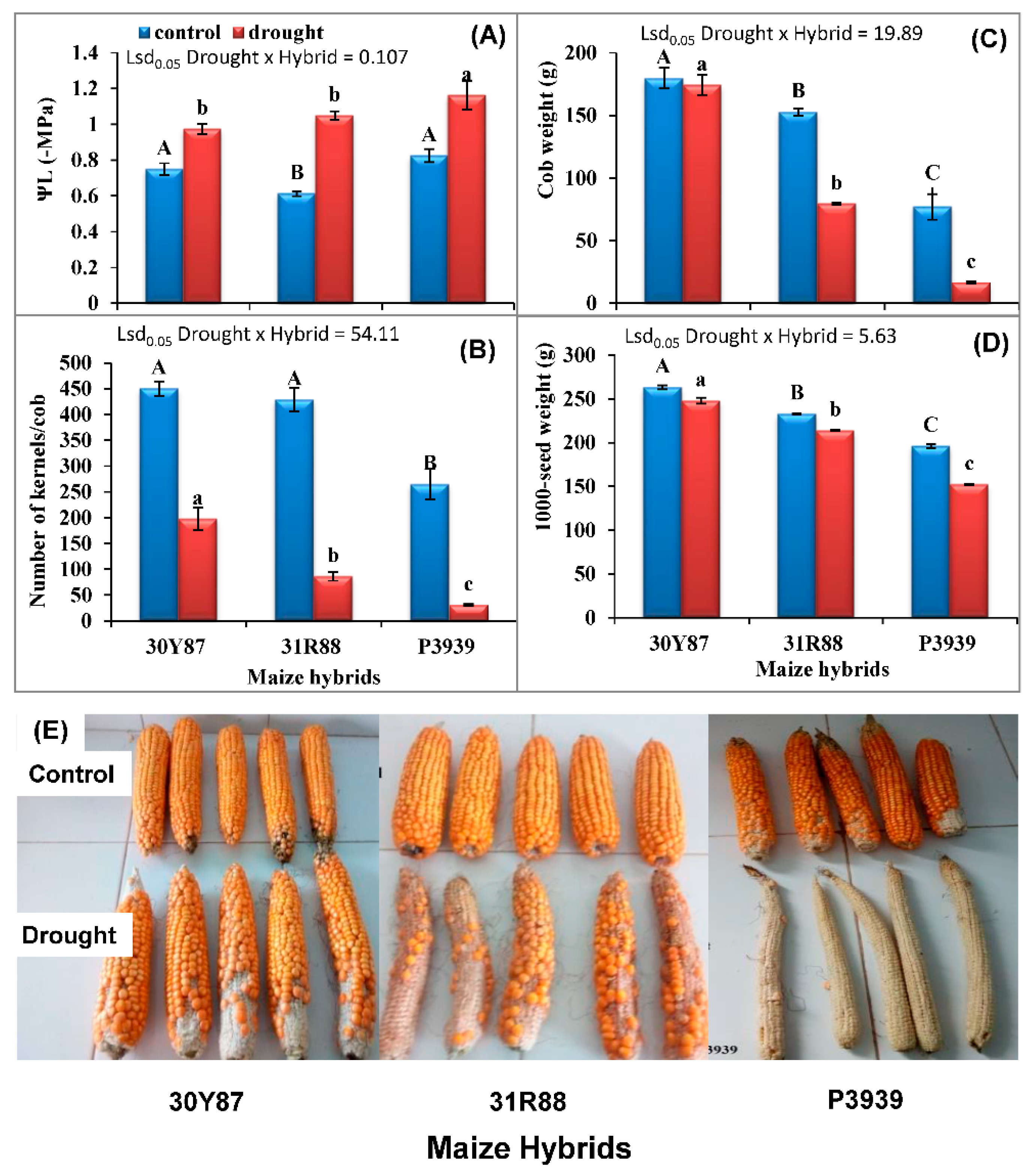
2.2. Measurement of Leaf Water Potential and Yield of Maize
3. Discussion
4. Materials and Methods
4.1. Pot Experiment
4.1.1. Measurement of PSII and PSI Activities as Rapid Light Curve Response
4.1.2. Measurement of Leaf Water Potential
4.2. Field Experiment
Yield Attributes
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blum, A. Heterosis, stress, and the environment: A possible road map towards the general improvement of crop yield. J. Exp. Bot. 2013, 64, 4829–4837. [Google Scholar] [CrossRef] [Green Version]
- Blum, A.; Tuberosa, R. Dehydration survival of crop plants and its measurement. J. Exp. Bot. 2018, 65, 975–981. [Google Scholar] [CrossRef]
- Ashraf, M.; Athar, H.-U.-R. Preface. Pak. J. Bot. 2006, 38, 1357. [Google Scholar]
- Athar, H.R.; Ashraf, M. Strategies for Crop Improvement Against Salinity and Drought Stress: An Overview. In Salinity and Water Stress: Improving Crop Efficiency; Ashraf, M., Ozturk, M., Athar, H.R., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 44, pp. 1–16. [Google Scholar]
- Ogbaga, C.C.; Stepien, P.; Athar, H.-U.-R.; Ashraf, M. Engineering Rubisco activase from thermophilic cyanobacteria into high-temperature sensitive plants. Crit. Rev. Biotechnol. 2018, 38, 559–572. [Google Scholar] [CrossRef]
- Athar, H.R.; Ashraf, M. Photosynthesis under drought stress. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; CRC Press: New York, NY, USA, 2005; pp. 795–810. [Google Scholar]
- Sanchez-Bragado, R.; Vicente, R.; Molero, G.; Serret, M.D.; Maydup, M.L.; Araus, J.L. New avenues for increasing yield and stability in C3 cereals: Exploring ear photosynthesis. Curr. Opin. Plant Biol. 2020, 56, 223–234. [Google Scholar] [CrossRef]
- Simkin, A.J.; Faralli, M.; Ramamoorthy, S.; Lawson, T. Photosynthesis in non-foliar tissues: Implications for yield. Plant J. 2020, 101, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Burgess, S.J.; de Becker, E.M.; Long, S.P. Photosynthesis in the fleeting shadows: An overlooked opportunity for increasing crop productivity? Plant J. 2020, 101, 874–884. [Google Scholar] [CrossRef]
- Araus, J.L.; Sanchez-Bragado, R.; Vicente, R. Improving crop yield and resilience through photosynthesis optimisation: Panacea or pipe dream? J. Exp. Bot. 2021, 72, 3936–3955. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Ruban, A.V.; Nixon, P.J. Photosynthesis solutions to enhance productivity. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160374. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Escalona, J.M.; Evain, S.; Gulias, J.; Moya, I.; Osmond, C.B.; Medrano, H. Steady-state chlorophyll fluorescence (Fs) measurements as a tool to follow variations of net CO2 assimilation and stomatal conductance during water-stress in C3 plants. Physiol. Plant. 2002, 114, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Sonoike, K. Photoinhibition and Protection of Photosystem I. In Photosystem I: The Light-Driven Plastocyanin: Ferredoxin Oxidoreductase; Golbeck, J.H., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 657–668. [Google Scholar]
- Asada, K. The water–water cycle in chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Bano, H.; Athar, H.-U.-R.; Zafar, Z.U.; Ogbaga, C.C.; Ashraf, M. Peroxidase activity and operation of photo-protective component of NPQ play key roles in drought tolerance of mung bean [Vigna radiata (L.) Wilcziek]. Physiol. Plant. 2021, 172, 603–614. [Google Scholar] [CrossRef]
- Ogbaga, C.C.; Athar, H.-U.-R. Inclusion of photoprotective parameters in photosynthesis-measuring systems to improve the interpretation of photosynthesis and productivity. Photosynthetica 2019, 57, 712–713. [Google Scholar] [CrossRef] [Green Version]
- Lima Neto, M.C.; Cerqueira, J.V.A.; Cunha, J.R.; Ribeiro, R.V.; Silveira, J.A.G. Cyclic electron flow, NPQ and photorespiration are crucial for the establishment of young plants of Ricinus communis and Jatropha curcas exposed to drought. Plant Biol. 2017, 19, 650–659. [Google Scholar] [CrossRef]
- Murchie, E.H.; Ruban, A.V. Dynamic non-photochemical quenching in plants: From molecular mechanism to productivity. Plant J. 2020, 101, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Signorelli, S.; Casaretto, E.; Sainz, M.; Díaz, P.; Monza, J.; Borsani, O. Antioxidant and photosystem II responses contribute to explain the drought-heat contrasting tolerance of two forage legumes. Plant Physiol. Biochem. 2013, 70, 195–203. [Google Scholar] [CrossRef]
- Ruban, A.V.; Wilson, S. The Mechanism of Non-Photochemical Quenching in Plants: Localization and Driving Forces. Plant Cell Physiol. 2021, 62, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Suorsa, M.; Rossi, F.; Tadini, L.; Labs, M.; Colombo, M.; Jahns, P.; Kater, M.M.; Leister, D.; Finazzi, G.; Aro, E.-M.; et al. PGR5-PGRL1-Dependent Cyclic Electron Transport Modulates Linear Electron Transport Rate in Arabidopsis thaliana. Mol. Plant 2016, 9, 271–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamim, F.; Athar, H.-U.-R.; Waheed, A. Role of osmolytes in degree of water stress tolerance in tomato. Pak. J. Phytopathol. 2013, 25, 6. [Google Scholar]
- Shamim, F.; Khan, K.; Athar, H.-U.-R.; Waheed, A. Protection of photosynthetic machinery by up-regulation of antioxidant enzymes in contrasting tomato genotypes under drought. Pak. J. Bot. 2015, 47, 1231–1239. [Google Scholar]
- Shamim, F.; Saqlan, S.M.; Athar, H.-U.-R.; Waheed, A. Screening and selection of tomato genotypes/cultivars for drought tolerance using multivariate analysis. Pak. J. Bot. 2014, 46, 1165–1178. [Google Scholar]
- Messmer, R.; Fracheboud, Y.; Banziger, M.; Vargas, M.; Stamp, P.; Ribaut, J.M. Drought stress and tropical maize: QTL-by-environment interactions and stability of QTLs across environments for yield components and secondary traits. Theor. Appl. Genet. 2009, 119, 913–930. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.N.; Ferreira-Silva, S.L.; Viégas, R.A.; Silveira, J.A.G. The role of organic and inorganic solutes in the osmotic adjustment of drought-stressed Jatropha curcas plants. Environ. Exp. Bot. 2010, 69, 279–285. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikkanen, M.; Mekala, N.R.; Aro, E.-M. Photosystem II photoinhibition-repair cycle protects Photosystem I from irreversible damage. Biochim. Biophys. Acta 2014, 1837, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauser, R.; Athar, H.U.R.; Ashraf, M. Chlorophyll fluorescence: A potential indicator for rapid assessment of water stress tolerance in canola (Brassica napus L.). Pak. J. Bot. 2006, 38, 1501–1509. [Google Scholar]
- Ogbaga, C.C.; Athar, H.-U.-R. The need to incorporate fast and slow relaxation kinetic parameters into photosynthesis-measuring systems. Sci. Afr. 2019, 4, e00106. [Google Scholar] [CrossRef]
- Athar, H.-U.-R.; Ambreen, S.; Javed, M.; Hina, M.; Rasul, S.; Zafar, Z.U.; Manzoor, H.; Ogbaga, C.C.; Afzal, M.; Al-Qurainy, F.; et al. Influence of sub-lethal crude oil concentration on growth, water relations and photosynthetic capacity of maize (Zea mays L.) plants. Environ. Sci. Pollut. Res. 2016, 23, 18320–18331. [Google Scholar] [CrossRef]
- Yi, X.-P.; Zhang, Y.-L.; Yao, H.-S.; Han, J.-M.; Chow, W.S.; Fan, D.-Y.; Zhang, W.-F. Changes in activities of both photosystems and the regulatory effect of cyclic electron flow in field-grown cotton (Gossypium hirsutum L.) under water deficit. J. Plant Physiol. 2018, 220, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Klughammer, C.; Schreiber, U. An improved method, using saturating light pulses, for the determination of photosystem I quantum yield via P700x+-absorbance changes at 830 nm. Planta 1994, 192, 261–268. [Google Scholar] [CrossRef]
- Kramer, D.M.; Evans, J.R. The Importance of Energy Balance in Improving Photosynthetic Productivity. Plant Physiol. 2011, 155, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Ruuska, S.A.; Badger, M.R.; Andrews, T.J.; von Caemmerer, S. Photosynthetic electron sinks in transgenic tobacco with reduced amounts of Rubisco: Little evidence for significant Mehler reaction. J. Exp. Bot. 2000, 51 (Suppl. 1), 357–368. [Google Scholar] [CrossRef] [Green Version]
- Golding, A.J.; Johnson, G.N. Down-regulation of linear and activation of cyclic electron transport during drought. Planta 2003, 218, 107–114. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | df | Y(II) | ETR(II) | qP |
|---|---|---|---|---|
| Treatment | 1 | 0.304 *** | 718.7 *** | 0.229 *** |
| PAR | 10 | 1.500 *** | 566.6 *** | 2.481 *** |
| Maize hybrids. | 2 | 0.234 *** | 725.4 *** | 0.631 *** |
| Treatment * PAR | 10 | 0.0261 *** | 52.50 *** | 0.040 *** |
| Treatment * hybrids. | 2 | 0.001 ns | 178.7 *** | 0.006 * |
| PAR * hybrids. | 20 | 0.028 *** | 52.96 *** | 0.054 *** |
| Treatment * PAR * hybrids. | 20 | 0.028 *** | 18.77 *** | 0.053 *** |
| Error | 198 | 9.635 | 2.930 | 0.001 *** |
| Total | 263 |
| Source of Variation | df | Y(NPQ) | NPQ | Y(NO) |
|---|---|---|---|---|
| Treatment | 1 | 1.244 *** | 1.055 *** | 0.005 ns |
| PAR | 10 | 2.042 *** | 5.173 *** | 0.090 *** |
| Maize hybrids. | 2 | 1.112 *** | 1.174 *** | 0.030 *** |
| Treatment * PAR | 10 | 0.065 *** | 0.820 *** | 0.033 *** |
| Treatment * hybrids. | 2 | 1.025 *** | 0.189 *** | 0.246 *** |
| PAR * hybrids. | 20 | 0.062 *** | 0.777 *** | 0.046 *** |
| Treatment * PAR * hybrids. | 20 | 0.073 *** | 0.332 *** | 0.020 *** |
| Error | 198 | 0.001 | 0.023 | 0.001 |
| Total | 263 |
| Source of Variation | df | Y(I) | ETR(I) | Y(ND) | Y(NA) |
|---|---|---|---|---|---|
| Treatment | 1 | 2.027 *** | 2357.4 *** | 0.882 *** | 1.769 *** |
| PAR | 10 | 1.031 *** | 5728.9 *** | 1.383 *** | 0.324 *** |
| Maize hybrids. | 2 | 0.616 *** | 1931.1 *** | 0.639 *** | 0.303 *** |
| Treatment * PAR | 10 | 0.092 *** | 83.53 *** | 0.131 *** | 0.122 *** |
| Treatment * hybrids. | 2 | 1.304 *** | 6970.2 *** | 0.878 *** | 1.099 *** |
| PAR * hybrids. | 20 | 0.017 *** | 141.9 *** | 0.114 *** | 0.008 *** |
| Treatment * PAR * hybrids. | 20 | 0.020 *** | 617.7 *** | 0.127 *** | 0.059 *** |
| Error | 198 | 0.002 | 11.18 | 0.002 | 0.002 |
| Total | 263 |
| Source of Variation | df | ΨL | 1000-Seed Weight | df | Cob Weight | Kernel No./Cob |
|---|---|---|---|---|---|---|
| Treatment | 1 | 0.666 *** | 4024.08 *** | 1 | 32155 *** | 1121760 *** |
| Maize hybrids | 2 | 0.059 *** | 13426.5 *** | 2 | 84941 *** | 150059 *** |
| Treatment * Maize hybrids | 2 | 0.022 * | 478.2 *** | 2 | 6508 *** | 19006.6 ** |
| Error | 18 | 0.005 | 14.38 | 54 | 492.5 | 3641.6 |
| Total | 23 | 59 |
| Parameters | Physiological Interplay | Calculated Formula |
|---|---|---|
| Y(II) | Effective quantum yield of PSII | ΦPSII = (FM′ − FS′/FM′) = ΔF/FM |
| ETR(II) | PSII electron transport rate | ETR = Y(II) × PAR × 0.42 |
| NPQ | Nonphotochemical quenching of Fm | (Fm − Fm′)/Fm′ |
| qP | Photochemical quenching based on ‘’puddle’’ model | (Fm′ − Fs′)/(Fm′ − Fo′) |
| Y(NPQ) | Quantum yield of pH-dependent energy dissipation in PSII | ΦNPQ = 1 − ΦPSII − 1/[NPQ + 1 + qL(FM/FO − 1)] Kramer et al., 2004) |
| Y(NO) | Quantum yield of nonregulated dissipation of energy in PSI | ΦNO = 1/[NPQ + 1 + qL × (FM/FO − 1)] Kramer et al., 2004) |
| Y(I) | Effective quantum yield of PSI photochemistry | (Pm′ − P/Pm) |
| ETR(I) | PSI electron transport rate | ETR = Y(I) × PAR × 0.42 |
| Y(ND) | Fraction of total P700 that is oxidized due to donor-side limitation | P/Pm |
| Y(NA) | Fraction of total P700 that cannot be oxidized due to lack of acceptors | Pm − Pm′/Pm |
| Y(CEF) | Cyclic electron flow estimated by Y(I) − Y(II) | Y(I) − Y(II) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, N.; Athar, H.-u.-R.; Kalaji, H.M.; Wróbel, J.; Mahmood, S.; Zafar, Z.U.; Ashraf, M. Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize? Int. J. Mol. Sci. 2021, 22, 13490. https://doi.org/10.3390/ijms222413490
Bashir N, Athar H-u-R, Kalaji HM, Wróbel J, Mahmood S, Zafar ZU, Ashraf M. Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize? International Journal of Molecular Sciences. 2021; 22(24):13490. https://doi.org/10.3390/ijms222413490
Chicago/Turabian StyleBashir, Nahidah, Habib-ur-Rehman Athar, Hazem M. Kalaji, Jacek Wróbel, Seema Mahmood, Zafar Ullah Zafar, and Muhammad Ashraf. 2021. "Is Photoprotection of PSII One of the Key Mechanisms for Drought Tolerance in Maize?" International Journal of Molecular Sciences 22, no. 24: 13490. https://doi.org/10.3390/ijms222413490