
AXIN2 Reduces the Survival of Porcine Induced Pluripotent Stem Cells (piPSCs)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
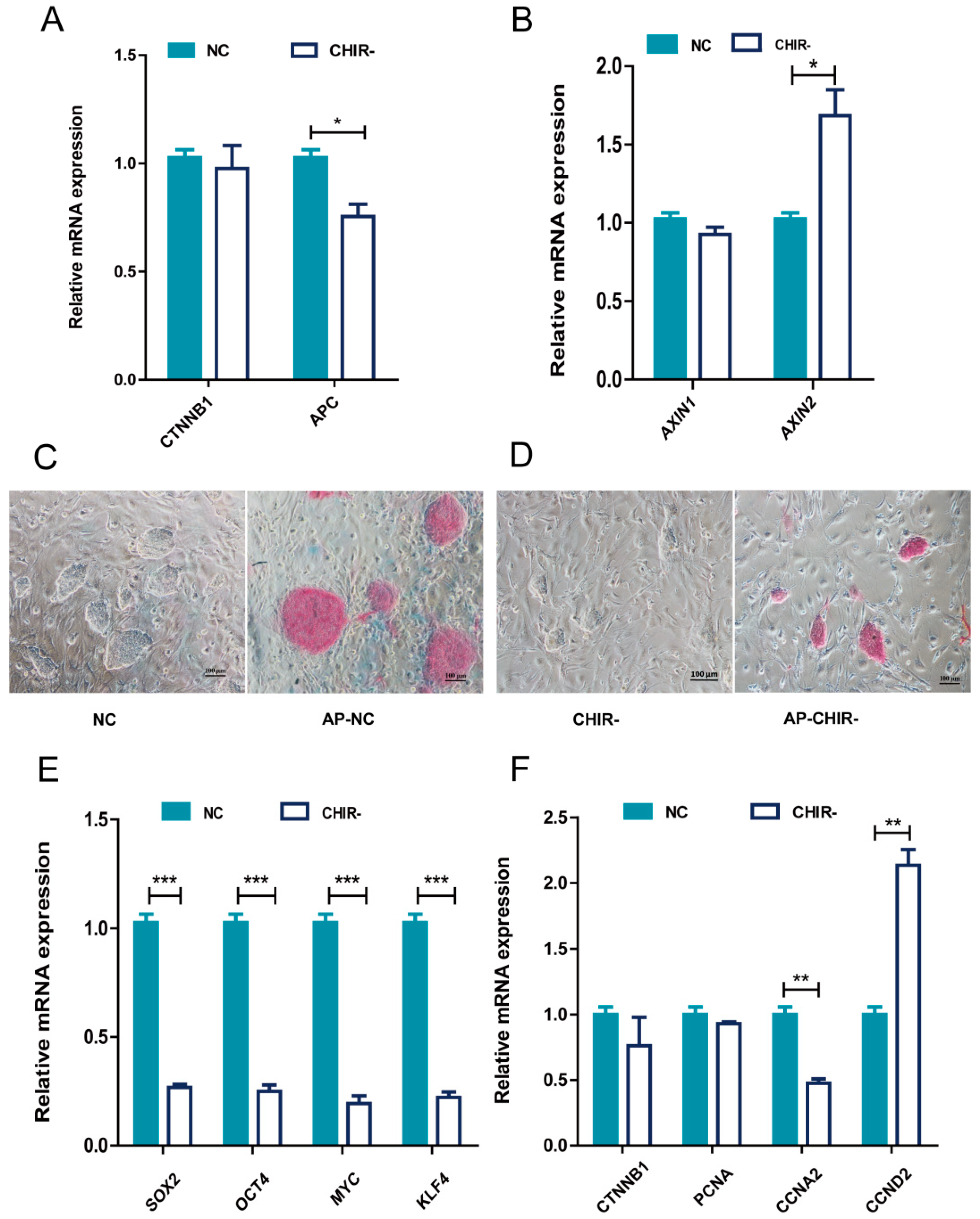
2.1. The Absence of CHIR99021 Affected the Pluripotency and Proliferation of piPSCs
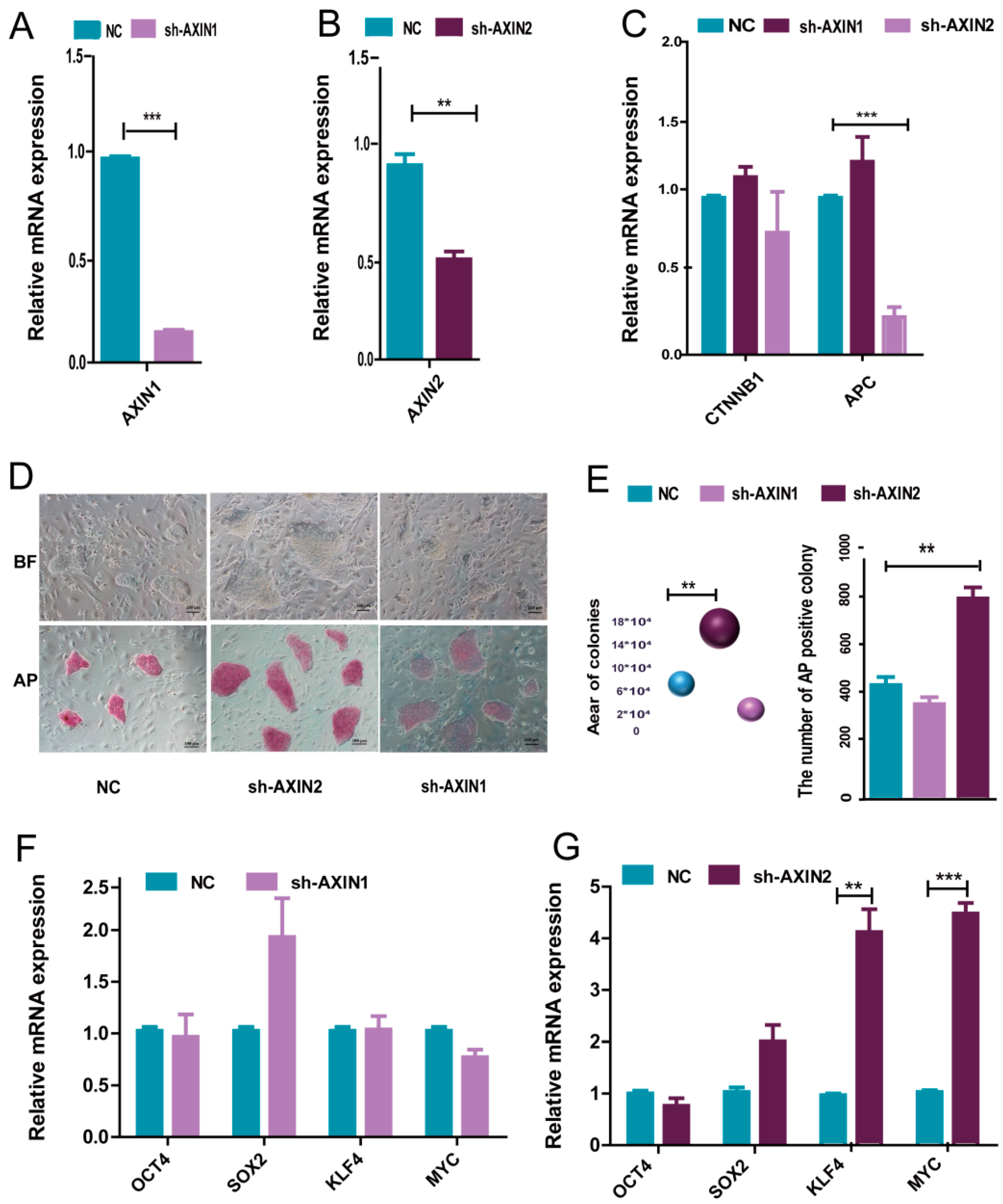
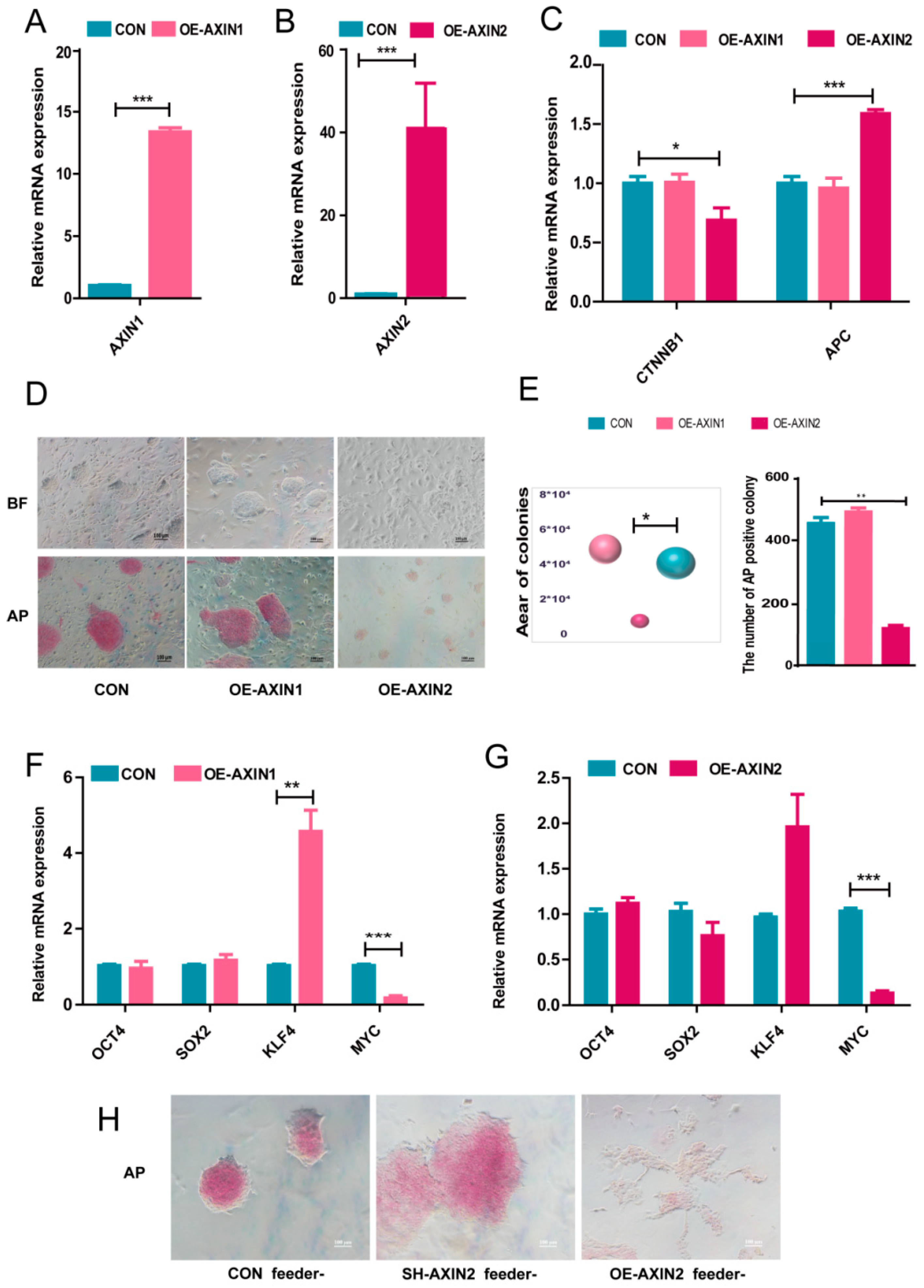
2.2. AXIN2 Had a Stronger Influence on the Pluripotency of piPSCs Than AXIN1
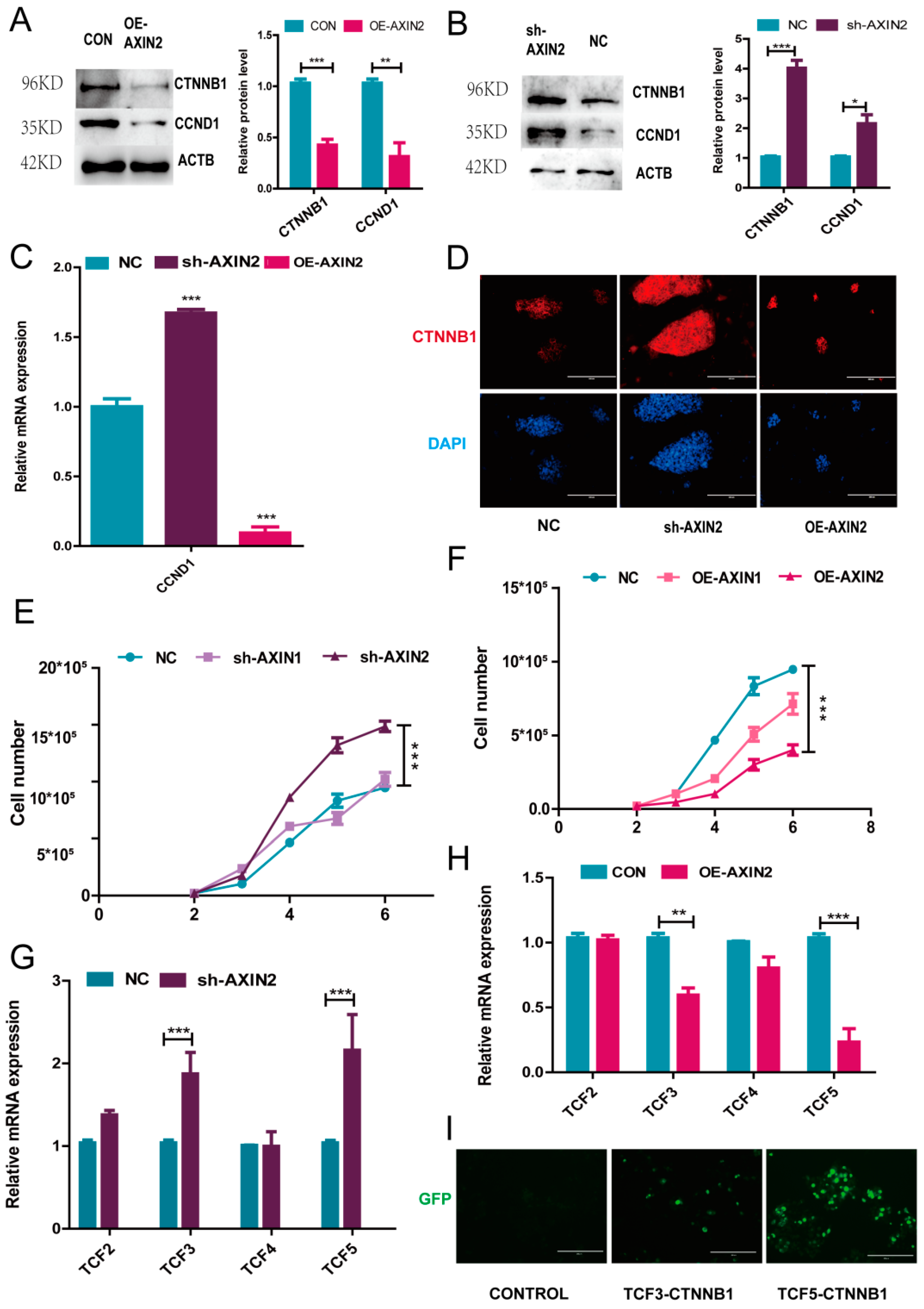
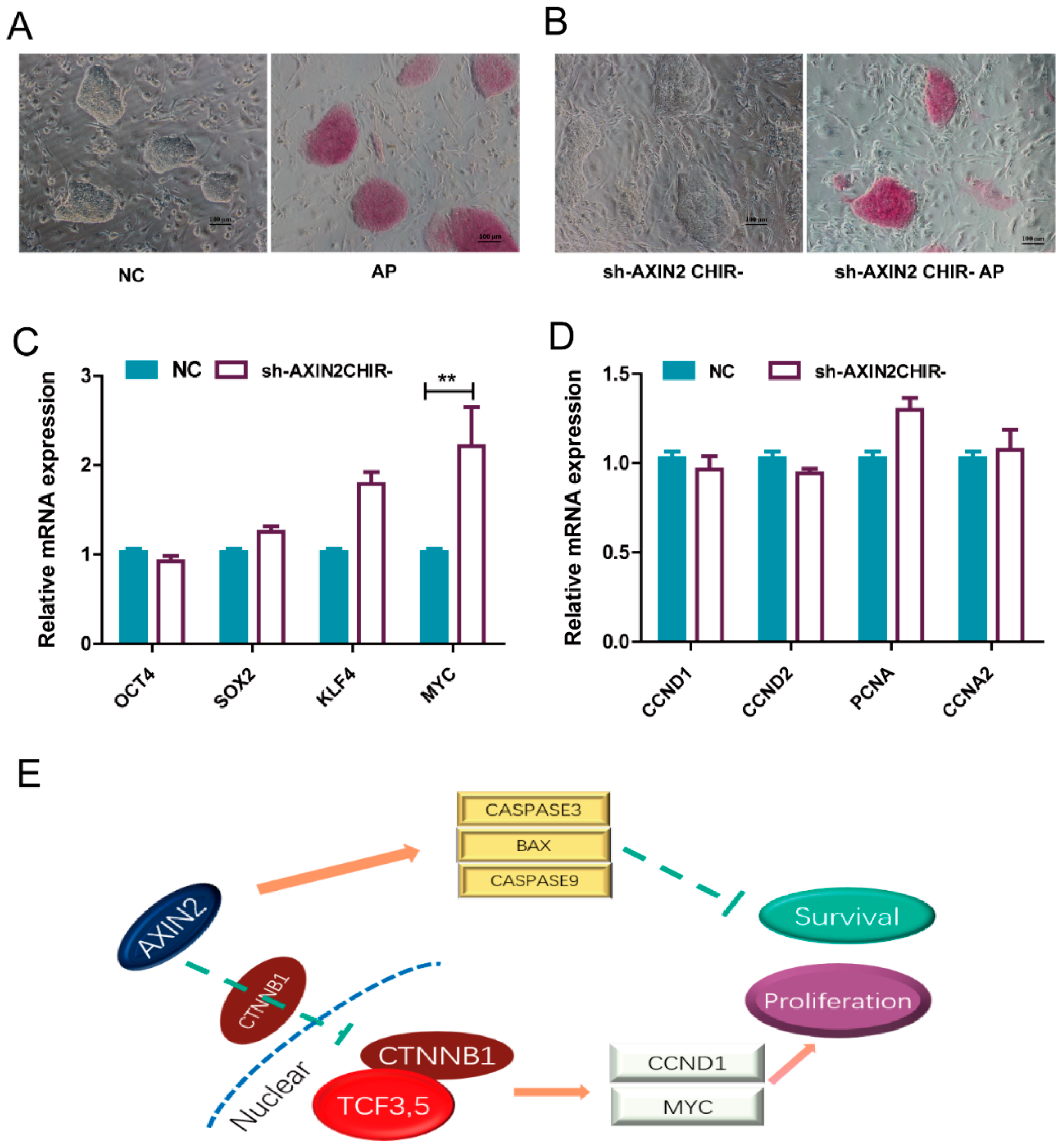
2.3. AXIN2 Regulates the Proliferation of piPSCs via CCND1
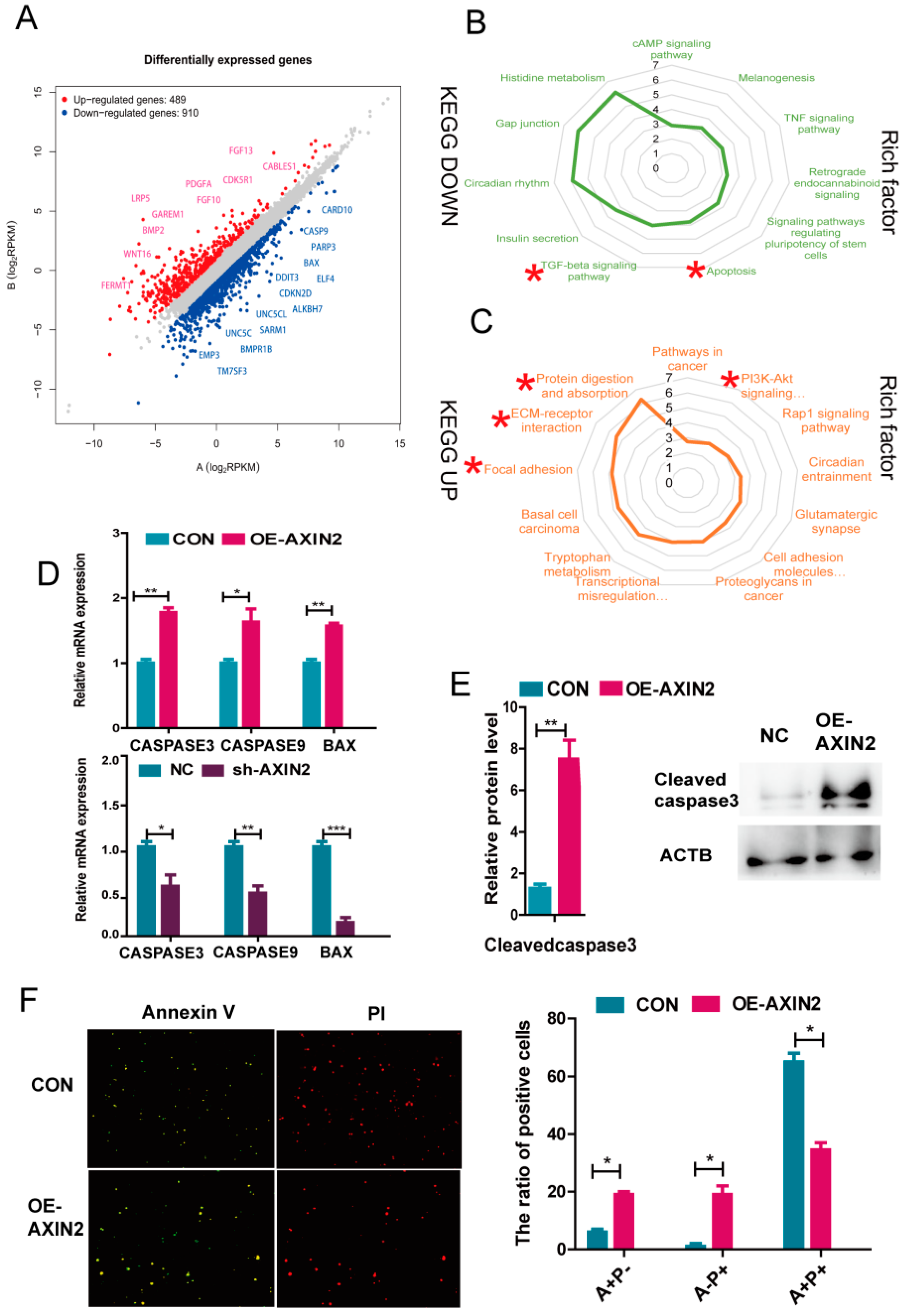
2.4. AXIN2 Knockdown Enhanced Anti-Apoptotic Ability of piPSCs through Inhibiting Caspase Family and BAX
2.5. piPSCs with Reduced Expression of AXIN2 Maintained Self-Renewal without CHIR99021
3. Discussion
4. Methods
4.1. Cell Culture
4.2. Plasmid and Cloning
4.3. Lentivirus Packaging and Transduction
4.4. AP Staining
4.5. RNA Extraction, Reverse Transcription and qRT-PCR Detection
4.6. Western Blot
4.7. Apoptosis Detection
4.8. RNA-Seq Analysis
4.9. Immunofluorescence Staining
4.10. Bimolecular Fluorescence Complementation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behrens, J.; Jerchow, B.A.; Wurtele, M.; Grimm, J.; Asbrand, C.; Wirtz, R.; Kuhl, M.; Wedlich, D.; Birchmeier, W. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science 1998, 280, 596–599. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, A.J.; Pierart, H.; Meek, S.; Bogerman, A.; Sutherland, L.; Murray, H.; Mountjoy, E.; Downing, A.; Talbot, R.; Sartori, C.; et al. Reprogramming pig fetal fibroblasts reveals a functional LIF signaling pathway. Cell. Reprogram. 2012, 14, 112–122. [Google Scholar] [CrossRef]
- Petkov, S.; Hyttel, P.; Niemann, H. The small molecule inhibitors PD0325091 and CHIR99021 reduce expression of pluripotency-related genes in putative porcine induced pluripotent stem cells. Cell. Reprogram. 2014, 16, 235–240. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Lin, S.C. Axin: A master scaffold for multiple signaling pathways. Neurosignals 2004, 13, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Lyashenko, N.; Winter, M.; Migliorini, D.; Biechele, T.; Moon, R.T.; Hartmann, C. Differential requirement for the dual functions of beta-catenin in embryonic stem cell self-renewal and germ layer formation. Nat. Cell Biol. 2011, 13, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Raggioli, A.; Junghans, D.; Rudloff, S.; Kemler, R. Beta-catenin is vital for the integrity of mouse embryonic stem cells. PLoS ONE 2014, 9, e86691. [Google Scholar]
- Wray, J.; Kalkan, T.; Gomez-Lopez, S.; Eckardt, D.; Cook, A.; Kemler, R.; Smith, A. Inhibition of glycogen synthase kinase-3 alleviates Tcf3 repression of the pluripotency network and increases embryonic stem cell resistance to differentiation. Nat. Cell Biol. 2011, 13, 838–845. [Google Scholar] [CrossRef] [Green Version]
- Sato, N.; Meijer, L.; Skaltsounis, L.; Greengard, P.; Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 2004, 10, 55–63. [Google Scholar] [CrossRef]
- Zhang, P.; Chang, W.H.; Fong, B.; Gao, F.; Liu, C.; Al Alam, D.; Bellusci, S.; Lu, W. Regulation of induced pluripotent stem (iPS) cell induction by Wnt/beta-catenin signaling. J. Biol. Chem. 2014, 289, 9221–9232. [Google Scholar] [CrossRef] [Green Version]
- Marson, A.; Foreman, R.; Chevalier, B.; Bilodeau, S.; Kahn, M.; Young, R.A.; Jaenisch, R. Wnt signaling promotes reprogramming of somatic cells to pluripotency. Cell Stem Cell 2008, 3, 132–135. [Google Scholar] [CrossRef] [Green Version]
- Li, V.S.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt signaling through inhibition of beta-catenin degradation in an intact Axin1 complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, A. Roles of Axin in the Wnt signalling pathway. Cell. Signal. 1999, 11, 777–788. [Google Scholar] [CrossRef]
- Leung, J.Y.; Kolligs, F.T.; Wu, R.; Zhai, Y.; Kuick, R.; Hanash, S.; Cho, K.R.; Fearon, E.R. Activation of AXIN2 expression by beta-catenin-T cell factor. A feedback repressor pathway regulating Wnt signaling. J. Biol. Chem. 2002, 277, 21657–21665. [Google Scholar] [CrossRef] [Green Version]
- Bernkopf, D.B.; Hadjihannas, M.V.; Behrens, J. Negative-feedback regulation of the Wnt pathway by conductin/axin2 involves insensitivity to upstream signalling. J. Cell Sci. 2015, 128, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Greely, H.T.; Jaenisch, R.; Nakauchi, H.; Rossant, J.; Belmonte, J.C.I. Stem cells and interspecies chimaeras. Nature 2016, 540, 51–59. [Google Scholar] [CrossRef]
- Ou, Q.; Jacobson, Z.; Abouleisa, R.R.E.; Tang, X.L.; Hindi, S.M.; Kumar, A.; Ivey, K.N.; Giridharan, G.; El-Baz, A.; Brittian, K.; et al. Physiological Biomimetic Culture System for Pig and Human Heart Slices. Circ. Res. 2019, 125, 628–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ma, Y.; Yang, J.Y.; Cheng, D.; Liu, X.; Ma, X.; West, F.D.; Wang, H. Comparative gene expression signature of pig, human and mouse induced pluripotent stem cell lines reveals insight into pig pluripotency gene networks. Stem Cell Rev. Rep. 2014, 10, 162–176. [Google Scholar] [CrossRef]
- Zhu, Z.; Wu, X.; Li, Q.; Zhang, J.; Yu, S.; Shen, Q.; Zhou, Z.; Pan, Q.; Yue, W.; Qin, D.; et al. Histone demethylase complexes KDM3A and KDM3B cooperate with OCT4/SOX2 to define a pluripotency gene regulatory network. FASEB J. 2021, 35, e21664. [Google Scholar] [CrossRef]
- Aulicino, F.; Pedone, E.; Sottile, F.; Lluis, F.; Marucci, L.; Cosma, M.P. Canonical Wnt Pathway Controls mESC Self-Renewal Through Inhibition of Spontaneous Differentiation via beta-Catenin/TCF/LEF Functions. Stem Cell Rep. 2020, 15, 646–661. [Google Scholar] [CrossRef]
- Chia, I.V.; Costantini, F. Mouse axin and axin2/conductin proteins are functionally equivalent in vivo. Mol. Cell Biol. 2005, 25, 4371–4376. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, R.; Hattori, Y.; Hosogi, S.; Toda, Y.; Takata, K.; Ashihara, E. A novel dipeptide type inhibitor of the Wnt/beta-catenin pathway suppresses proliferation of acute myelogenous leukemia cells. Biochem. Biophys. Res. Commun. 2021, 535, 73–79. [Google Scholar] [CrossRef]
- Fu, F.; Deng, Q.; Li, R.; Wang, D.; Yu, Q.X.; Yang, X.; Lei, T.Y.; Han, J.; Pan, M.; Zhen, L.; et al. AXIN2 gene silencing reduces apoptosis through regulating mitochondria-associated apoptosis signaling pathway and enhances proliferation of ESCs by modulating Wnt/beta-catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 418–427. [Google Scholar] [PubMed]
- Figeac, N.; Zammit, P.S. Coordinated action of Axin1 and Axin2 suppresses beta-catenin to regulate muscle stem cell function. Cell. Signal. 2015, 27, 1652–1665. [Google Scholar] [CrossRef]
- Faunes, F.; Hayward, P.; Descalzo, S.M.; Chatterjee, S.S.; Balayo, T.; Trott, J.; Christoforou, A.; Ferrer-Vaquer, A.; Hadjantonakis, A.K.; Dasgupta, R.; et al. A membrane-associated beta-catenin/Oct4 complex correlates with ground-state pluripotency in mouse embryonic stem cells. Development 2013, 140, 1171–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz Descalzo, S.; Rue, P.; Faunes, F.; Hayward, P.; Jakt, L.M.; Balayo, T.; Garcia-Ojalvo, J.; Martinez Arias, A. A competitive protein interaction network buffers Oct4-mediated differentiation to promote pluripotency in embryonic stem cells. Mol. Syst. Biol. 2013, 9, 694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardehali, R.; Inlay, M.A.; Ali, S.R.; Tang, C.; Drukker, M.; Weissman, I.L. Overexpression of BCL2 enhances survival of human embryonic stem cells during stress and obviates the requirement for serum factors. Proc. Natl. Acad. Sci. USA 2011, 108, 3282–3287. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Pan, Q.; Zhao, W.; Wu, X.; Yu, S.; Shen, Q.; Zhang, J.; Yue, W.; Peng, S.; Li, N.; et al. BCL2 enhances survival of porcine pluripotent stem cells through promoting FGFR2. Cell Prolif. 2021, 54, e12932. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, R.; Shen, Q.; Zhu, Z.; Zhang, J.; Wu, X.; Zhao, W.; Li, N.; Yang, F.; Wei, H.; et al. ESRRB Facilitates the Conversion of Trophoblast-Like Stem Cells From Induced Pluripotent Stem Cells by Directly Regulating CDX2. Front. Cell Dev. Biol. 2021, 9, 712224. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Wang, H.; Dai, B.; Wang, X.; Zhou, D.; Shen, J.; Guo, F.; Wang, J.; Zhou, J.; Wang, H.; et al. Human induced pluripotent stem cell-derived cardiomyocytes reveal abnormal TGFbeta signaling in type 2 diabetes mellitus. J. Mol. Cell Cardiol. 2020, 142, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Itoh, F.; Watabe, T.; Miyazono, K. Roles of TGF-beta family signals in the fate determination of pluripotent stem cells. Semin. Cell Dev. Biol. 2014, 32, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Rui, H.; Wang, J.; Lin, S.; He, Y.; Chen, M.; Li, Q.; Ye, Z.; Zhang, S.; Chan, S.C.; et al. Axin is a scaffold protein in TGF-beta signaling that promotes degradation of Smad7 by Arkadia. EMBO J. 2006, 25, 1646–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Yu, T.; Cai, Y.; Wang, H. Preserving self-renewal of porcine pluripotent stem cells in serum-free 3i culture condition and independent of LIF and b-FGF cytokines. Cell Death Discov. 2018, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.L.; Zhu, Z.S.; Xiao, X.; Zhou, Z.; Yu, S.; Shen, Q.Y.; Zhang, J.Q.; Yue, W.; Zhang, R.; He, X.; et al. LIN28A inhibits DUSP family phosphatases and activates MAPK signaling pathway to maintain pluripotency in porcine induced pluripotent stem cells. Zool. Res. 2021, 42, 377–388. [Google Scholar] [CrossRef]
- Yue, W.; Sun, J.; Zhang, J.; Chang, Y.; Shen, Q.; Zhu, Z.; Yu, S.; Wu, X.; Peng, S.; Li, N.; et al. Mir-34c affects the proliferation and pluripotency of porcine induced pluripotent stem cell (piPSC)-like cells by targeting c-Myc. Cells Dev. 2021, 166, 203665. [Google Scholar] [CrossRef]
- Shen, Q.Y.; Yu, S.; Zhang, Y.; Zhou, Z.; Zhu, Z.S.; Pan, Q.; Lv, S.; Niu, H.M.; Li, N.; Peng, S.; et al. Characterization of porcine extraembryonic endoderm cells. Cell Prolif. 2019, 52, e12591. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.D.; Du, X.M.; Yang, D.H.; Ma, F.L.; Yu, X.W.; Zhang, M.F.; Li, N.; Peng, S.; Liao, M.Z.; Li, G.P.; et al. Dmrt1 regulates the immune response by repressing the TLR4 signaling pathway in goat male germline stem cells. Zool. Res. 2021, 42, 14–27. [Google Scholar] [CrossRef]
- Miller, K.E.; Kim, Y.; Huh, W.K.; Park, H.O. Bimolecular Fluorescence Complementation (BiFC) Analysis: Advances and Recent Applications for Genome-Wide Interaction Studies. J. Mol. Biol. 2015, 427, 2039–2055. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Yu, S.; Shen, Q.; Zhao, W.; Zhang, J.; Wu, X.; Zhu, Z.; Wu, X.; Li, N.; Peng, S.; et al. AXIN2 Reduces the Survival of Porcine Induced Pluripotent Stem Cells (piPSCs). Int. J. Mol. Sci. 2021, 22, 12954. https://doi.org/10.3390/ijms222312954
Zhang R, Yu S, Shen Q, Zhao W, Zhang J, Wu X, Zhu Z, Wu X, Li N, Peng S, et al. AXIN2 Reduces the Survival of Porcine Induced Pluripotent Stem Cells (piPSCs). International Journal of Molecular Sciences. 2021; 22(23):12954. https://doi.org/10.3390/ijms222312954
Chicago/Turabian StyleZhang, Rui, Shuai Yu, Qiaoyan Shen, Wenxu Zhao, Juqing Zhang, Xiaolong Wu, Zhenshuo Zhu, Xiaojie Wu, Na Li, Sha Peng, and et al. 2021. "AXIN2 Reduces the Survival of Porcine Induced Pluripotent Stem Cells (piPSCs)" International Journal of Molecular Sciences 22, no. 23: 12954. https://doi.org/10.3390/ijms222312954