
Singapore Grouper Iridovirus Disturbed Glycerophospholipids Homeostasis: Cytosolic Phospholipase A2 Was Essential for Virus Replication

Abstract
:1. Introduction
2. Results
2.1. Infection with SGIV Disturbed Glycerophospholipids (GPs) Homeostasis
2.2. Infection with SGIV Resulted in Alteration of Fatty Acyls (FAs)
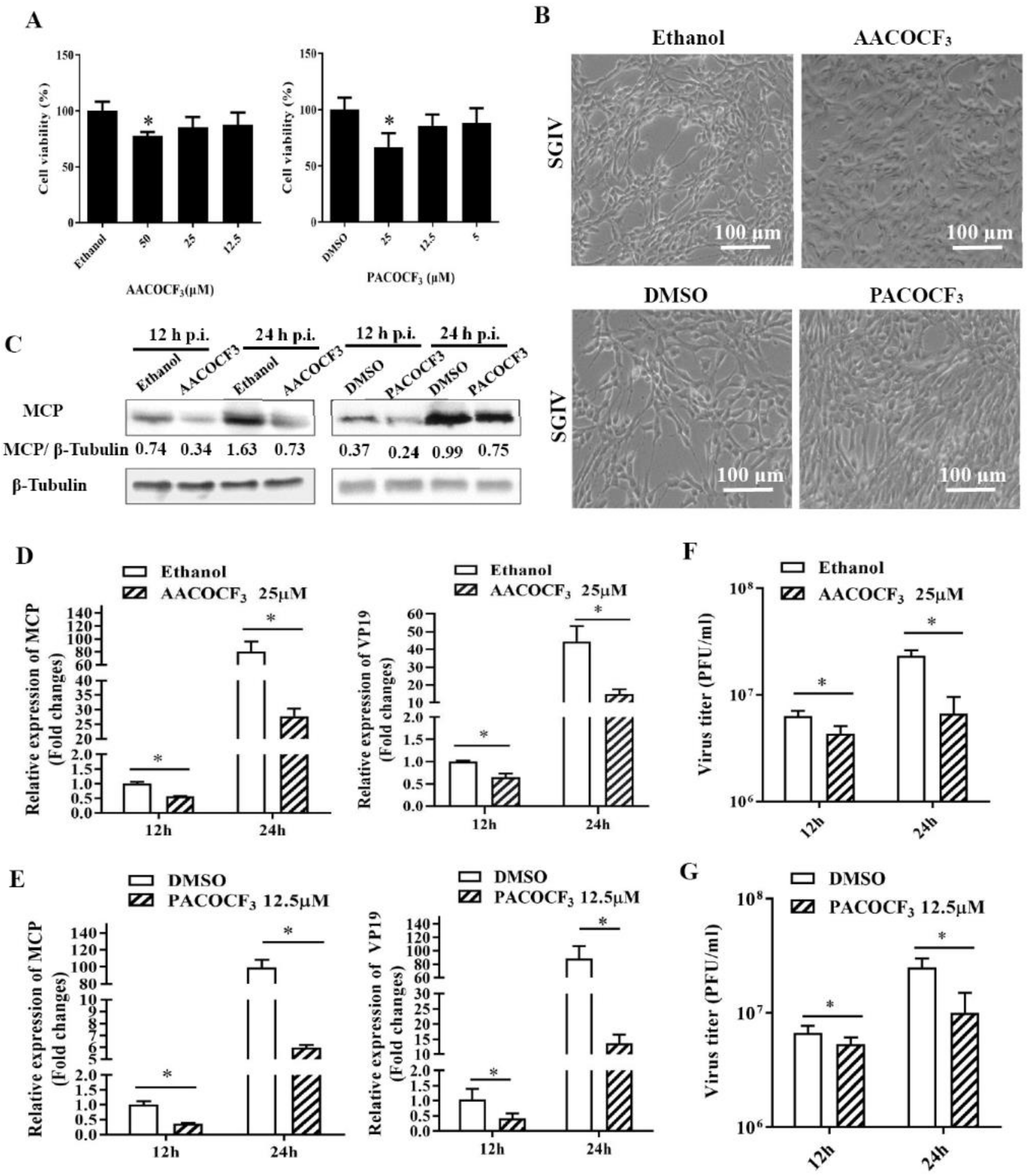
2.3. Pharmacological Inhibition of PLA2 Activity Affected SGIV Replication
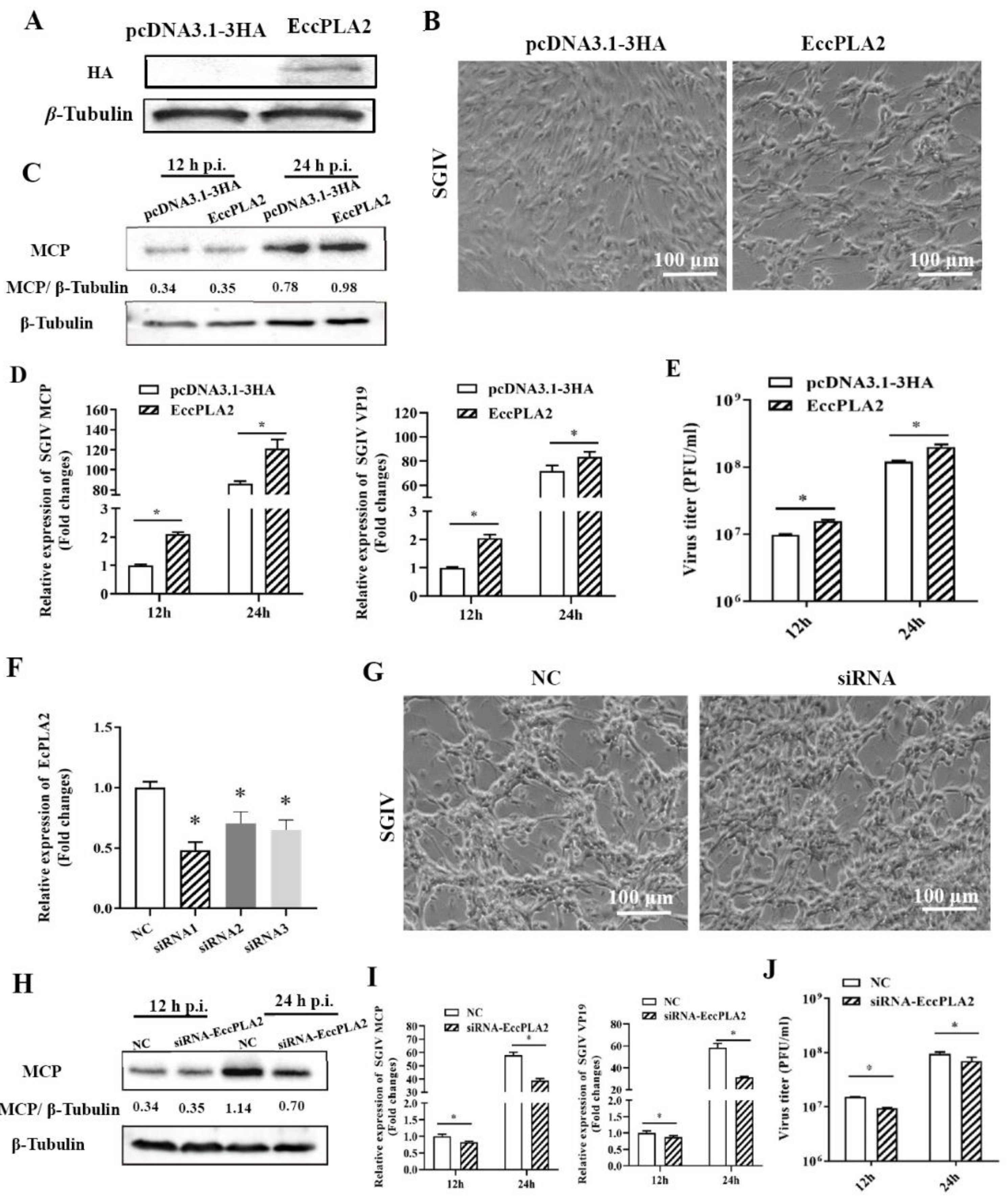
2.4. Grouper Cytosolic Phospholipase A2-Alpha (EccPLA2α) Was Essential for SGIV Infection
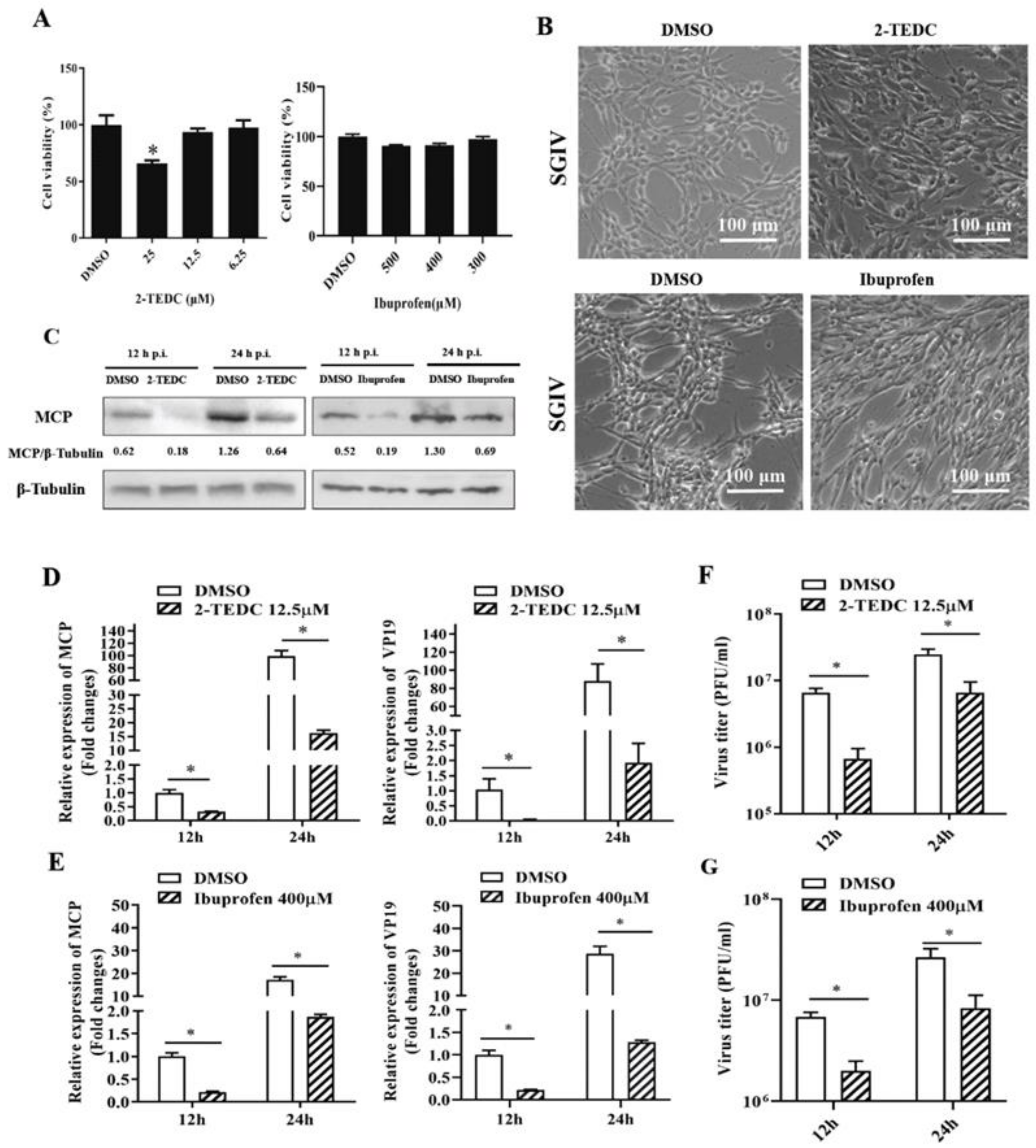
2.5. Inhibition of Arachidonic Acid Metabolism Impaired SGIV Replication
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Virus, and Reagents
4.2. Virus Infection
4.3. Lipidomic Analysis
4.4. Cell Transfection
4.5. Virus Titer Assay
4.6. Western Blotting
4.7. RNA Extraction, cDNA Synthesis, and Quantitative PCR (qPCR)
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chinchar, V.G.; Yu, K.H.; Jancovich, J.K. The molecular biology of frog virus 3 and other iridoviruses infecting cold-blooded vertebrates. Viruses 2011, 3, 1959–1985. [Google Scholar] [CrossRef] [Green Version]
- Rothermel, B.B.; Travis, E.R.; Miller, D.L.; Hill, R.L.; McGuire, J.L.; Yabsley, M.J. High occupancy of stream salamanders despite high ranavirus prevalence in a southern appalachians watershed. Ecohealth 2011, 10, 184–189. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Waltzek, T.B.; Subramaniam, K. Ranaviruses and other members of the family Iridoviridae: Their place in the virosphere. Virology 2017, 511, 259–271. [Google Scholar] [CrossRef]
- Braunwald, J.; Nonnenmacher, H.; Tripier-Darcy, F. Ultrastructural and biochemical of frog virus 3 uptake by BHK-21 cells. J. Gen. Virol. 1985, 66 Pt 2, 283–293. [Google Scholar] [CrossRef]
- Wang, S.W.; Huang, X.H.; Huang, Y.H.; Hao, X.; Xu, H.J.; Cai, M.J.; Wang, H.D.; Qin, Q.W. Entry of a novel marine DNA virus, Singapore grouper iridovirus, into host cells occurs via clathrin-mediated endocytosis and macropinocytosis in a pH-dependent manner. J. Virol. 2014, 88, 13047–13063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Tran, B.N.; Wang, F.; Ounjai, P.; Wu, J.; Hew, C.L. Visualization of Assembly Intermediates and Budding Vacuoles of Singapore Grouper Iridovirus in Grouper Embryonic Cells. Sci. Rep. 2016, 6, 18696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutsafi, Y.; Fridmann-Sirkis, Y.; Milrot, E.; Hevroni, L.; Minsky, A. Infection cycles of large DNA viruses: Emerging themes and underlying questions. Virology 2014, 466–467, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.H.; Huang, Y.H.; Yuan, X.P.; Zhang, Q.Y. Electron microscopic examination of the viromatrix of Rana grylio virus in a fish cell line. J. Virol. Methods 2006, 133, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Ketter, E.; Randall, G. Virus Impact on Lipids and Membranes. Annu. Rev. Virol. 2019, 6, 319–340. [Google Scholar] [CrossRef]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; Pasa-Tolic, L.; Metz, T.O.; Adamec, J.; Kuhn, R.J. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 2012, 8, e1002584. [Google Scholar] [CrossRef]
- Chotiwan, N.; Andre, B.G.; Sanchez-Vargas, I.; Islam, M.N.; Grabowski, J.M.; Hopf-Jannasch, A.; Gough, E.; Nakayasu, E.; Blair, C.D.; Belisle, J.T.; et al. Dynamic remodeling of lipids coincides with dengue virus replication in the midgut of Aedes aegypti mosquitoes. PLoS Pathog. 2018, 14, e1006853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.; Chu, H.; Yang, D.; Sze, K.H.; Lai, P.M.; Yuan, S.; Shuai, H.; Wang, Y.; Kao, R.Y.; Chan, J.F.; et al. Characterization of the Lipidomic Profile of Human Coronavirus-Infected Cells: Implications for Lipid Metabolism Remodeling upon Coronavirus Replication. Viruses 2019, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Gaunt, E.R.; Zhang, Q.; Cheung, W.; Wakelam, M.J.O.; Lever, A.M.L.; Desselberger, U. Lipidome analysis of rotavirus-infected cells confirms the close interaction of lipid droplets with viroplasms. J. Gen. Virol. 2013, 94 Pt 7, 1576–1586. [Google Scholar] [CrossRef]
- Queiroz, A.; Pinto, I.F.D.; Lima, M.; Giovanetti, M.; de Jesus, J.G.; Xavier, J.; Barreto, F.K.; Canuto, G.A.B.; do Amaral, H.R.; de Filippis, A.M.B.; et al. Lipidomic Analysis Reveals Serum Alteration of Plasmalogens in Patients Infected With ZIKA Virus. Front. Microbiol. 2019, 10, 753. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Zou, Z.; Chu, H.; Chan, G.; Tsang, J.O.; Lai, P.M.; Yuan, S.; Yip, C.C.; Yin, F.; Kao, R.Y.; et al. Lipidomic Profiling Reveals Significant Perturbations of Intracellular Lipid Homeostasis in Enterovirus-Infected Cells. Int. J. Mol. Sci. 2019, 20, 5952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, E.P. The biological synthesis of phospholipids. Can. J. Biochem. Physiol. 1956, 34, 334–348. [Google Scholar] [CrossRef]
- Liebscher, S.; Ambrose, R.L.; Aktepe, T.E.; Mikulasova, A.; Prier, J.E.; Gillespie, L.K.; Lopez-Denman, A.J.; Rupasinghe, T.W.T.; Tull, D.; McConville, M.J.; et al. Phospholipase A2 activity during the replication cycle of the flavivirus West Nile virus. PLoS Pathog. 2018, 14, e1007029. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Hardt, M.; Schwudke, D.; Neuman, B.W.; Pleschka, S.; Ziebuhr, J. Inhibition of Cytosolic Phospholipase A (2) alpha Impairs an Early Step of Coronavirus Replication in Cell Culture. J. Virol. 2018, 92, e01463-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Pei, R.; Guo, M.; Han, Q.; Lai, J.; Wang, Y.; Wu, C.; Zhou, Y.; Lu, M.; Chen, X. Cytosolic phospholipase A2 gamma is involved in hepatitis C virus replication and assembly. J. Virol. 2012, 86, 13025–13037. [Google Scholar] [CrossRef] [Green Version]
- Menzel, N.; Fischl, W.; Hueging, K.; Bankwitz, D.; Frentzen, A.; Haid, S.; Gentzsch, J.; Kaderali, L.; Bartenschlager, R.; Pietschmann, T. MAP-kinase regulated cytosolic phospholipase A2 activity is essential for production of infectious hepatitis C virus particles. PLoS Pathog. 2012, 8, e1002829. [Google Scholar] [CrossRef] [Green Version]
- Jeewandara, C.; Gomes, L.; Udari, S.; Paranavitane, S.A.; Shyamali, N.L.; Ogg, G.S.; Malavige, G.N. Secretory phospholipase A2 in the pathogenesis of acute dengue infection. Immun. Inflamm. Dis. 2016, 5, 7–15. [Google Scholar] [CrossRef]
- Gibson-Kueh, S.; Netto, P.; Ngoh-Lim, G.H.; Chang, S.F.; Ho, L.L.; Qin, Q.W.; Chua, F.H.; Ng, M.L.; Ferguson, H.W. The pathology of systemic iridoviral disease in fish. J. Comp. Pat. 2003, 129, 111–119. [Google Scholar] [CrossRef]
- Qin, Q.W.; Chang, S.F.; Ngoh-Lim, G.H.; Gibson-Kueh, S.; Shi, C.; Lam, T.J. Characterization of a novel ranavirus isolated from grouper Epinephelus tauvina. Dis. Aquat. Organ. 2003, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.W.; Lam, T.J.; Sin, Y.M.; Shen, H.; Chang, S.F.; Ngoh, G.H.; Chen, C.L. Electron microscopic observations of a marine fish iridovirus isolated from brown-spotted grouper, Epinephelus tauvina. J. Virol. Methods 2001, 98, 17–24. [Google Scholar] [CrossRef]
- Wu, J.; Chan, R.; Wenk, M.R.; Hew, C.L. Lipidomic study of intracellular Singapore grouper iridovirus. Virology 2010, 399, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Huang, Y.; Ouyang, Z.; Xu, L.; Yan, Y.; Cui, H.; Han, X.; Qin, Q. Singapore grouper iridovirus, a large DNA virus, induces nonapoptotic cell death by a cell type dependent fashion and evokes ERK signaling. Apoptosis 2011, 16, 831–845. [Google Scholar] [CrossRef]
- Pham, P.H.; Lai, Y.S.; Lee, F.F.; Bols, N.C.; Chiou, P.P. Differential viral propagation and induction of apoptosis by grouper iridovirus (GIV) in cell lines from three non-host species. Virus Res. 2012, 167, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, D.; Hashidate, T.; Shimizu, T.; Shindou, H. Diversity and function of membrane glycerophospholipids generated by the remodeling pathway in mammalian cells. J. Lipid Res. 2014, 55, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, J.S. Immunomodulation by eicosanoids and anti-inflammatory drugs. Curr. Opin. Immunol. 1989, 2, 264–268. [Google Scholar] [CrossRef]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, R.; Ghorpade, A. Cytosolic phospholipase A2 regulates alcohol-mediated astrocyte inflammatory responses in HIV-associated neurocognitive disorders. Cell Death Discov. 2015, 1, 15045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, C.; Xu, F.; Zhang, S.; You, J.; You, X.; Qiu, L.; Zheng, J.; Ye, L.; Zhang, X. Hepatitis B virus X protein promotes liver cell proliferation via a positive cascade loop involving arachidonic acid metabolism and p-ERK1/2. Cell Res. 2010, 20, 563–575. [Google Scholar] [CrossRef]
- Heaton, N.S.; Randall, G. Dengue virus-induced autophagy regulates lipid metabolism. Cell Host Microbe 2010, 8, 422–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Acebes, M.A.; Blazquez, A.B.; Jimenez de Oya, N.; Escribano-Romero, E.; Saiz, J.C. West Nile virus replication requires fatty acid synthesis but is independent on phosphatidylinositol-4-phosphate lipids. PLoS ONE 2011, 6, e24970. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhang, Y.; Zheng, J.; Wang, L.; Qin, Q.; Huang, X. Metabolic profiles of fish nodavirus infection in vitro: RGNNV induced and exploited cellular fatty acid synthesis for virus infection. Cell Microbiol. 2020, 22, e13216. [Google Scholar] [CrossRef]
- Leventis, P.A.; Grinstein, S. The distribution and function of phosphatidylserine in cellular membranes. Annu. Rev. Biophys. 2010, 39, 407–427. [Google Scholar] [CrossRef]
- Calder, P.C. Eicosanoids. Essays. Biochem. 2020, 64, 423–441. [Google Scholar]
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Hirabayashi, T.; Yamamoto, K. Recent progress in phospholipase A (2) research: From cells to animals to humans. Prog. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef]
- Chen, N.; Warner, J.L.; Reiss, C.S. NSAID treatment suppresses VSV propagation in mouse CNS. Virology 2000, 276, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, N.; Bisher, M.E.; Enquist, L.W. Cyclooxygenase-1 and -2 are required for production of infectious pseudorabies virus. J. Virol. 2004, 78, 12964–12974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, H.A.; Kim, M.S.; Jang, S.Y.; Lee, Y.M.; Ahn, J.H.; Lee, C.H. Cellular signals involved in cyclooxygenase-2 expression induced by human cytomegalovirus. Virus Res. 2009, 146, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, Y.; OuYang, Z.; Cai, J.; Yan, Y.; Qin, Q. Roles of stress-activated protein kinases in the replication of Singapore grouper iridovirus and regulation of the inflammatory responses in grouper cells. J. Gen. Virol. 2011, 92 Pt 6, 1292–1301. [Google Scholar] [CrossRef]
- Huang, X.H.; Huang, Y.H.; Sun, J.J.; Han, X.; Qin, Q.W. Characterization of two grouper Epinephelus akaara cell lines: Application to studies of Singapore grouper iridovirus (SGIV) propagation and virus-host interaction. Aquaculture 2009, 292, 172–179. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Sequence (5′-3′) |
|---|---|
| EcPLA2α-3HA-KpnI-F | CGGGGTACCATGGCTTCCAATATAATTGTGGA |
| EcPLA2α-3HA-BamHI-R | CGCGGATCCCTGGTTATCAGTTCCCAGGA |
| siRNA1-EcPLA2α | GCAGCAGUUCUCUCACAAATT |
| siRNA2-EcPLA2α | GCACAACAUCCUGGAGUUATT |
| siRNA3-EcPLA2α | GGUGGAGUUCAGCCCGUAUTT |
| EcPLA2α-RT-F | CAGTGATGGTGGTTCG |
| EcPLA2α-RT-R | ATGTTGTGCTGGTTGG |
| Actin-RT-F | TACGAGCTGCCTGACGGACA |
| Actin-RT-R | GGCTGTGATCTCCTTCTGCA |
| SGIV MCP-RT-F | GCA CGCTTCTCTCACCTTCA |
| SGIV MCP-RT-R | AACGGCAACGGGAGCACTA |
| SGIV VP19-RT-F | TCCAAGGGAGAAACTGTAAG |
| SGIV VP19-RT-R | GGGGTAAGCGTGAAGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, N.; Zheng, J.; Wang, W.; Zhi, L.; Qin, Q.; Huang, Y.; Huang, X. Singapore Grouper Iridovirus Disturbed Glycerophospholipids Homeostasis: Cytosolic Phospholipase A2 Was Essential for Virus Replication. Int. J. Mol. Sci. 2021, 22, 12597. https://doi.org/10.3390/ijms222212597
Ni N, Zheng J, Wang W, Zhi L, Qin Q, Huang Y, Huang X. Singapore Grouper Iridovirus Disturbed Glycerophospholipids Homeostasis: Cytosolic Phospholipase A2 Was Essential for Virus Replication. International Journal of Molecular Sciences. 2021; 22(22):12597. https://doi.org/10.3390/ijms222212597
Chicago/Turabian StyleNi, Na, Jiaying Zheng, Wenji Wang, Linyong Zhi, Qiwei Qin, Youhua Huang, and Xiaohong Huang. 2021. "Singapore Grouper Iridovirus Disturbed Glycerophospholipids Homeostasis: Cytosolic Phospholipase A2 Was Essential for Virus Replication" International Journal of Molecular Sciences 22, no. 22: 12597. https://doi.org/10.3390/ijms222212597