
Immunobiological Properties and Clinical Applications of Interleukin-38 for Immune-Mediated Disorders: A Systematic Review Study

 ,
,
Abstract
:1. Introduction
2. Methodology
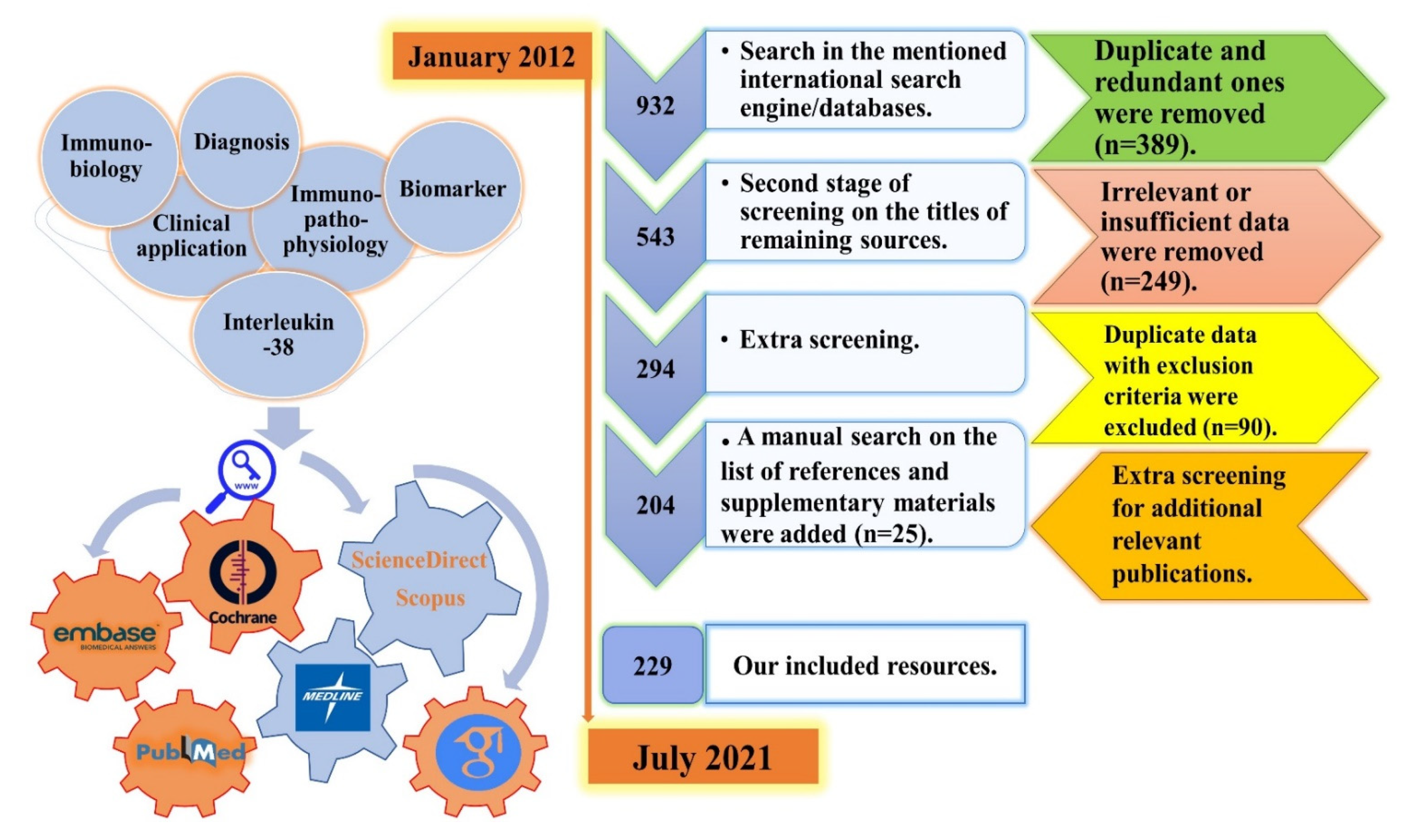
2.1. Literature Search Strategy, and Screening Process
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction
2.4. Quality Assessment and Bio-Statistical Analysis
2.5. Ethical Statement
3. Results
3.1. Immunobiology of IL-38
3.1.1. Activation of the IL-1 Superfamily Members
3.1.2. IL-38 Receptors
IL-1R1
IL-36R
IL-1 Receptor Accessory Protein-like 1 (IL-1RAPL1)
3.1.3. Inhibitory Properties of IL-38
3.2. Role of IL-38 in Immune-Mediated Diseases
3.2.1. Role of IL-38 in Autoimmune Diseases
Rheumatoid Arthritis (RA)
Systemic Lupus Erythematosus (SLE)
Psoriasis
Atopic Dermatitis (AD)
Kawasaki Disease (KD)
Gouty Arthritis
Sjögren Syndrome (SS)
Crohn’s Disease (CD)
HLA-B27-Associated Anterior Uveitis and Idiopathic Anterior Uveitis (IAU)
3.2.2. Role of IL-38 in Inflammatory Disorders
Inflammation-Induced Corneal Neovascularization
Hidradenitis Suppurativa (HS)
Chronic Primary Angle Closure Glaucoma (CPACG)
Retinal Ischemia and Proliferative Vascular Diseases
3.2.3. Role of IL-38 in Metabolic Disorders
Type2 Diabetes Mellitus (T2DM) and Obesity
Pregnancy and Gestational Diabetes Mellitus (GDM)
Hyperlipidemia
3.2.4. Role of IL-38 in CVDs
Myocardial Infarction (MI)
3.2.5. IL-38 and Cancer Co-Relation
3.2.6. Role of IL-38 in Lung and Respiratory Diseases
Lung Fibrosis
Lung Adenocarcinoma
Chronic Obstructive Pulmonary Disease (COPD) and Acute Lung Injury (ALI)
Drug-Induced ILD
3.2.7. Role of IL-38 in Infectious Diseases
Hepatitis
Lethal Sepsis
Aspergillosis
4. Future Directions and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Adeno-associated virus | AAV |
| Activator protein-1 | AP-1 |
| Acute lung injury | ALI |
| Acute primary angle closure glaucoma | APACG |
| Acute respiratory distress syndrome | ARDS |
| Age related cataract | ARC |
| Antigen-induced arthritis | AIA |
| A proliferation-inducing ligand | APRIL |
| Bronchoalveolar lavage fluid | BALF |
| Behcet’s disease | BD |
| Body mass index | BMI |
| Cardiac troponin I | cTnINT |
| Cardiovascular diseases | CVDs |
| C–C motif chemokine ligand | CCL |
| Cecal ligation puncture | CLP |
| Centers for Disease Control and Prevention | CDC |
| Central nervous system | CNS |
| Chronic obstructive pulmonary disease | COPD |
| Chronic primary angle closure glaucoma | CPACG |
| Collagen-induced arthritis | CIA |
| Colorectal cancer | CRC |
| Compensatory anti-inflammatory responses syndrome | CARS |
| Coronary artery disease | CADs |
| C-reactive protein | CRP |
| Crohn’s disease | CD |
| C-X-C motif chemokine ligand | CXCL |
| Dendritic cells | DCs |
| Enzyme linked immunosorbent assay | ELISA |
| Fibroblast-like synoviocytes | FLSs |
| Granulocyte-colony stimulating factor | G-CSF |
| Gestational diabetes mellitus | GDM |
| Hepatitis B virus | HBV |
| Hidradenitis suppurativa | HS |
| Idiopathic anterior uveitis | IAU |
| Idiopathic pulmonary fibrosis | IPF |
| Imiquimod | IMQ |
| Inflammatory bowel disease | IBD |
| Interstitial lung disease | ILD |
| Interferon-gamma | IFN-γ |
| Interleukin | IL |
| Interleukin-1 receptor accessory protein | IL-1RAcP |
| Interleukin-1 receptor accessory protein-like 1 | IL-1RAcPL1 |
| Kawasaki disease | KD |
| Left ventricular ejection fraction | LVEF |
| Lipopolysaccharide | LPS |
| Macrophage-colony stimulating factor | M-CSF |
| Mean deviation of visual field | MDVF |
| Monocyte chemoattractant protein-1 | MCP-1 |
| Myocardial infarction | MI |
| Non-small cell lung cancer | NSCLC |
| Normal human keratinocytes | NHKs |
| Nuclear localization signal | NLS |
| Nuclear factor-kappa B | NF-κB |
| Osteoarthritis | OA |
| Ovalbumin | OVA |
| Oxygen-induced retinopathy | OIR |
| Percutaneous coronary intervention | PCI |
| Peripheral blood mononuclear cells | PBMCs |
| Plasmacytoid dendritic cells | pDCs |
| Primary Sjögren syndrome | pSS |
| ProB-type natriuretic peptide | NTproBNP |
| Reactive oxygen species | ROS |
| Receptor antagonist | RA |
| Reverse transcriptase-polymerase chain reaction | RT-PCR |
| Rheumatoid arthritis | RA |
| Serum transfer-induced arthritis | STIA |
| Single nucleotide polymorphism | SNP |
| Sjögren syndrome | SS |
| ST-elevated myocardial infarction | STEMI |
| Systematic lupus erythematosus | SLE |
| T helper | THP |
| Toll like receptor-4 | TLR-4 |
| Tumor necrosis factor | TNF |
| Type1 diabetes mellitus | T1DM |
| Type2 diabetes mellitus | T2DM |
| Ulcerative colitis | UC |
| Vascular endothelial cells growth factor | VEGF |
| Virologic response | VR |
| World Health Organization | WHO |
References
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I.; Glass, C.K. Anti-Inflammatory Therapy in Chronic Disease: Challenges and Opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosh, E.; Bahmaie, N.; Esmaeilzadeh, A. Evolution in Immune Gene Therapy of Glioblastoma; Interleukin-37 as a Novel Candidate. Clin Oncol. 2019, 4, 1618. [Google Scholar]
- Moghadam, S.; Erfanmanesh, M.; Esmaeilzadeh, A. Interleukin 35 and Hepatocyte Growth Factor; as a novel combined immune gene therapy for Multiple Sclerosis disease. Med. Hypotheses 2017, 109, 102–105. [Google Scholar] [CrossRef]
- Esmaeilzadeh, A.; Mohammadzadeh, A.; Bahmaie, N. New Generation of Promising Immunotherapeutics Approaches for Psoriasis Dilemma; IL-35 Gene as a Potentiated Candidate. Inflamm. Cell Signal. 2018, 5, 91171950. [Google Scholar]
- Bahmaie, N.; Faghihzadeh, S.; Esmaeilzadeh, A.; Amini, B. Immunomodulatory Effects of Helicobacter pylori on Pro and Anti-inflammatory Cytokines Production in Peripheral Whole Blood Cells Culture. Biosci. Biotechnol. Res. Asia 2016, 13, 2221–2230. [Google Scholar] [CrossRef]
- Bahmaie, N.; Esmaeilzadeh, A. Sepsis biomarkers; A new approach in the diagnosis and follow up treatment of infectious diseases. Lab. Diagn. 2016, 7, 53–64. [Google Scholar]
- Elias, M.; Zhao, S.; Le, H.T.; Wang, J.; Neurath, M.F.; Neufert, C.; Fiocchi, C.; Rieder, F. IL-36 in chronic inflammation and fibrosis—bridging the gap? J. Clin. Investig. 2021, 131, e144336. [Google Scholar] [CrossRef]
- Martin, P.; Goldstein, J.D.; Mermoud, L.; Diaz-Barreiro, A.; Palmer, G. IL-1 Family Antagonists in Mouse and Human Skin Inflammation. Front. Immunol. 2021, 12, 652846. [Google Scholar] [CrossRef]
- Iznardo, H.; Puig, L. The interleukin-1 family cytokines in psoriasis: Pathogenetic role and therapeutic perspectives. Expert Rev. Clin. Immunol. 2021, 17, 187–199. [Google Scholar] [CrossRef]
- Fields, J.; Günther, S.; Sundberg, E.J. Structural Basis of IL-1 Family Cytokine Signaling. Front. Immunol. 2019, 10, 1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Guo, Y.; Xiao, L.; Pan, W.; Ma, J. Dendritic cells should not be overlooked when studying the effect of IL-38 administration in arthritis. Ann. Rheum. Dis. 2017, 77, e12. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Stoeckman, A.K.; Wu, G.; Boeckermann, A.N.; Azam, T.; Netea, M.G.; Joosten, L.A.B.; van der Meer, J.W.M.; Hao, R.; Kalabokis, V.; et al. IL-38 binds to the IL-36 receptor and has biological effects on immune cells similar to IL-36 receptor antagonist. Proc. Natl. Acad. Sci. USA 2012, 109, 3001–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, F.; Esmaeilzadeh, A. IL-1R2: A novel approach for gene therapy in atherosclerosis. Hypothesis J. 2016, 14, e1. [Google Scholar]
- Dinarello, C.A. Introduction to the interleukin-1 family of cytokines and receptors: Drivers of innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Fietta, P.; Costa, E.; Delsante, G. Interleukins (ILs), a fascinating family of cytokines. Part II: ILs from IL-20 to IL-38. Theor. Biol. Forum 2015, 108, 19–40. [Google Scholar]
- Tsutsui, H.; Cai, X.; Hayashi, S. Interleukin-1 Family Cytokines in Liver Diseases. Mediat. Inflamm. 2015, 2015, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Khosh, E.; Bahmaie, N.; Elahi, R.; Esmaeilzadeh, A. Clinical Applications of Interleukin-37; As a Key Player in the Immunopathogenesis of Immune Disorders. Iran. J. Allergy Asthma Immunol. 2020, 19, 209–228. [Google Scholar] [CrossRef]
- Esmaeilzadeh, A.; Pouyan, S.; Erfanmanesh, M. Is Interleukin-38 a key player cytokine in atherosclerosis immune gene therapy? Med. Hypotheses 2019, 125, 139–143. [Google Scholar] [CrossRef]
- Manesh, M.E.; Esmaeilzadeh, A.; Mirzaei, M.H. IL-24: A novel gene therapy candidate for immune system upregulation in Hodgkin’s lymphoma. J. Medical Hypotheses Ideas 2015, 9, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, M.H.; Esmaeilzadeh, A. Overexpression of MDA-7/IL-24 as an anticancer cytokine in gene therapy of thyroid carcinoma. J. Med. Hypotheses Ideas 2014, 8, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Piri, Z.; Esmaeilzadeh, A.; Hajikhanmirzaei, M. Interleukin-25 as a candidate gene in immunogene therapy of pancreatic cancer. J. Med. Hypotheses Ideas 2012, 6, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Gallenga, C.E.; Pandolfi, F.; Caraffa, A.; Kritas, S.K.; Ronconi, G.; Toniato, E.; Martinotti, S.; Conti, P. Interleukin-1 family cytokines and mast cells: Activation and inhibition. J. Boil. Regul. Homeost. Agents 2019, 33, 1–6. [Google Scholar]
- Wang, M.; Wang, B.; Ma, Z.; Sun, X.; Tang, Y.; Li, X.; Wu, X. Detection of the novel IL-1 family cytokines by QAH-IL1F-1 assay in rheumatoid arthritis. Cell. Mol. Boil. 2016, 62, 31–34. [Google Scholar]
- Vaillant, A.A.J.; Qurie, A. Interleukin. In StatPearls Internet; StatPearls Publishing: Bethesda, MD, USA, 2018. [Google Scholar]
- Conti, P. Impact of cytokines in mast cells allergic inflammation. Inter. Trends Immun. 2013, 1, 5–16. [Google Scholar]
- Yuan, X.L.; Li, Y.; Pan, X.H.; Zhou, M.; Gao, Q.Y.; Li, M.C. Production of recombinant human interleukin-38 and its inhibitory effect on the expression of proinflammatory cytokines in THP-1 cells. Mol. Biol. 2016, 50, 405–411. [Google Scholar] [CrossRef]
- Talabot-Ayer, D.; Mermoud, L.; Borowczyk, J.; Drukala, J.; Wolnicki, M.; Modarressi, A.; Boehncke, W.-H.; Brembilla, N.; Palmer, G. Interleukin-38 interacts with destrin/actin-depolymerizing factor in human keratinocytes. PLoS ONE 2019, 14, e0225782. [Google Scholar] [CrossRef] [Green Version]
- Moorlag, S.J.; Röring, R.J.; Joosten, L.A.; Netea, M.G. The role of the interleukin-1 family in trained immunity. Immunol. Rev. 2017, 281, 28–39. [Google Scholar] [CrossRef]
- Mora, J.; Han, Y.; Wittig, I.; Putyrski, M.; Ernst, A.; Brüne, B.; Weigert, A. Apoptosis-induced Tolerance: Role of the Novel Cytokine IL-38: P116. Cytokine 2016, 87, 109–110. [Google Scholar]
- Mattii, M.; Ayala, F.; Balato, N.; Filotico, R.; Lembo, S.; Schiattarella, M.; Patruno, C.; Marone, G.; Balato, A. The balance between pro- and anti-inflammatory cytokines is crucial in human allergic contact dermatitis pathogenesis: The role of IL-1 family members. Exp. Dermatol. 2013, 22, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The Interleukin-1 Family: Back to the Future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef] [Green Version]
- Palomo, J.; Dietrich, D.; Martin, P.; Palmer, G.; Gabay, C. The interleukin (IL)-1 cytokine family—Balance between agonists and antagonists in inflammatory diseases. Cytokine 2015, 76, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Günther, S.; Sundberg, E.J. Molecular determinants of agonist and antagonist signaling through the IL-36 receptor. J. Immunol. 2014, 193, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Schett, G.; Dayer, J.-M.; Manger, B. Interleukin-1 function and role in rheumatic disease. Nat. Rev. Rheumatol. 2016, 12, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Quan, Y.; Yue, Y.; Heng, X.; Che, F. Interleukin-37: A crucial cytokine with multiple roles in disease and potentially clinical therapy (Review). Oncol. Lett. 2018, 15, 4711–4719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Huang, Z.; Li, H.; Liu, X.; Zheng, S.G.; Su, W. IL-38: A New Player in Inflammatory Autoimmune Disorders. Biomolecules 2019, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.; Arend, W.; Sims, J.; Smith, D.; Blumberg, H.; O’Neill, L.; Goldbach-Mansky, R.; Pizarro, T.; Hoffman, H.; Bufler, P.; et al. IL-1 family nomenclature. Nat. Immunol. 2010, 11, 973. [Google Scholar] [CrossRef] [Green Version]
- Lukens, J.R.; Gross, J.M.; Kanneganti, T.-D. IL-1 family cytokines trigger sterile inflammatory disease. Front. Immunol. 2012, 3, 315. [Google Scholar] [CrossRef] [Green Version]
- Karumbaiah, L.; Norman, S.E.; Rajan, N.B.; Anand, S.; Saxena, T.; Betancur, M.; Patkar, R.; Bellamkonda, R.V. The upregulation of specific interleukin (IL) receptor antagonists and paradoxical enhancement of neuronal apoptosis due to electrode induced strain and brain micromotion. Biomaterials 2012, 33, 5983–5996. [Google Scholar] [CrossRef]
- Italiani, P.; Carlesi, C.; Giungato, P.; Puxeddu, I.; Borroni, B.; Bossù, P.; Migliorini, P.; Siciliano, G.; Boraschi, D. Evaluating the levels of interleukin-1 family cytokines in sporadic amyotrophic lateral sclerosis. J. Neuroinflammation 2014, 11, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, S.; Smith, D.W.; Gardiner, B.; Grodzinsky, A.J. Systems Based Study of the Therapeutic Potential of Small Charged Molecules for the Inhibition of IL-1 Mediated Cartilage Degradation. PLoS ONE 2016, 11, e0168047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Moschen, A.R.; Szabo, G. Interleukin-1 and inflammasomes in alcoholic liver disease/acute alcoholic hepatitis and nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Hepatology 2016, 64, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Harris, J. Autophagy and IL-1 family cytokines. Front. Immunol. 2013, 4, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayer, J.-M.; Oliviero, F.; Punzi, L. A Brief History of IL-1 and IL-1 Ra in Rheumatology. Front. Pharmacol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Lamacchia, C.; Palmer, G.; Rodriguez, E.; Martin, P.; Vigne, S.; Seemayer, C.A.; Talabot-Ayer, D.; Towne, J.E.; Gabay, C. The severity of experimental arthritis is independent of IL-36 receptor signaling. Arthritis Res. Ther. 2013, 15, R38. [Google Scholar] [CrossRef] [Green Version]
- Lopetuso, L.R.; Chowdhry, S.; Pizarro, T.T. Opposing Functions of Classic and Novel IL-1 Family Members in Gut Health and Disease. Front. Immunol. 2013, 4, 181. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J. Cell death and inflammation: The case for IL-1 family cytokines as the canonical DAMP s of the im-mune system. FEBS J. 2016, 283, 2599–2615. [Google Scholar] [CrossRef]
- Voronov, E.; Apte, R.N. IL-1 in Colon Inflammation, Colon Carcinogenesis and Invasiveness of Colon Cancer. Cancer Microenviron. 2015, 8, 187–200. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Interleukin-1 in cardiac injury, repair, and remodeling: Pathophysiologic and translational concepts. Discoveries 2015, 3, e41. [Google Scholar] [CrossRef] [Green Version]
- Buhl, A.-L.; Wenzel, J. Interleukin-36 in Infectious and Inflammatory Skin Diseases. Front. Immunol. 2019, 10, 1162. [Google Scholar] [CrossRef]
- Striz, I. Cytokines of the IL-1 family: Recognized targets in chronic inflammation underrated in organ trans-plantations. Clin. Sci. 2017, 131, 2241–2256. [Google Scholar] [CrossRef]
- Garraud, T.; Harel, M.; Boutet, M.-A.; Le Goff, B.; Blanchard, F. The enigmatic role of IL-38 in inflammatory diseases. Cytokine Growth Factor Rev. 2018, 39, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Catalan-Dibene, J.; McIntyre, L.L.; Zlotnik, A. Interleukin 30 to Interleukin 40. J. Interferon Cytokine Res. 2018, 38, 423–439. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.S.; Kaiser, P.; Fife, M. The chicken IL-1 family: Evolution in the context of the studied vertebrate lineage. Immunogenetics 2014, 66, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A. Overview of the interleukin-1 family of ligands and receptors. Semin. Immunol. 2013, 25, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, L.A. The IL-1 cytokine family and its role in inflammation and fibrosis in the lung. Semin. Immunopathol. 2016, 38, 517–534. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, L.; Rui, W.; Li, X.; Xuan, D.; Zheng, S.; Yu, Y.; Zhang, J.; Kong, N.; Zhu, X.; et al. New Interleukins in Psoriasis and Psoriatic Arthritis Patients: The Possible Roles of Interleukin-33 to Interleukin-38 in Disease Activities and Bone Erosions. Dermatology 2017, 233, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Huang, M.; Yao, Y.-M. Recent advances in the biology of IL-1 family cytokines and their potential roles in development of sepsis. Cytokine Growth Factor Rev. 2019, 45, 24–34. [Google Scholar] [CrossRef]
- Kwak, A.; Lee, Y.; Kim, H.; Kim, S. Intracellular interleukin (IL)-1 family cytokine processing enzyme. Arch. Pharmacal. Res. 2016, 39, 1556–1564. [Google Scholar] [CrossRef]
- Gresnigt, M.S.; van de Veerdonk, F.L. Biology of IL-36 cytokines and their role in disease. Semin. Immunol. 2013, 25, 458–465. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, D.; van de Veerdonk, F.; Eisenmesser, E.; Joosten, L.A.B.; Dinarello, C. OP0170 Human il-38 reduces joint inflammation in a mouse model of gouty arthritis. Ann. Rheum. Dis. 2018, 77, 135. [Google Scholar]
- Palomo, J.; Troccaz, S.; Talabot-Ayer, D.; Rodriguez, E.; Palmer, G. The severity of imiquimod-induced mouse skin inflammation is independent of endogenous IL-38 expression. PLoS ONE 2018, 13, e0194667. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.-S.; Wang, C.-J.; Lin, S.-H.; Xu, F. Research progress of inflammatory immunity and regulation mechanism of interleukin-38. Chin. J. Immunol. 2019, 12, 230–235. [Google Scholar]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor β, and TNF-α: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresnigt, M.S.; van de Veerdonk, F.L. The role of interleukin-1 family members in the host defence against Aspergillus fumigatus. Mycopathologia 2014, 178, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Lin, S.; Yan, X.; Wang, C.; Tu, H.; Yin, Y.; Cao, J. Interleukin 38 Protects Against Lethal Sepsis. J. Infect. Dis. 2018, 218, 1175–1184. [Google Scholar] [CrossRef]
- Alunno, A.; Carubbi, F.; Giacomelli, R.; Gerli, R. Cytokines in the pathogenesis of rheumatoid arthritis: New players and therapeutic targets. BMC Rheumatol. 2017, 1, 1–13. [Google Scholar] [CrossRef]
- Mora, J.; Schlemmer, A.; Wittig, I.; Richter, F.; Putyrski, M.; Frank, A.-C.; Han, Y.; Jung, M.; Ernst, A.; Weigert, A.; et al. Interleukin-38 is released from apoptotic cells to limit inflammatory macrophage responses. J. Mol. Cell Biol. 2016, 8, 426–438. [Google Scholar] [CrossRef]
- Shaik, Y.; Sabatino, G.; Maccauro, G.; Varvara, G.; Murmura, G.; Saggini, A.; Rosati, M.; Conti, F.; Cianchetti, E.; Caraffa, A.; et al. IL-36 Receptor Antagonist with Special Emphasis on IL-38; SAGE Publications Sage: London, UK, 2013. [Google Scholar]
- Tsai, Y.-C.; Tsai, T.-F. Anti-interleukin and interleukin therapies for psoriasis: Current evidence and clinical use-fulness. Therapeutic Adv. Musculoskelet. Dis. 2017, 9, 277–294. [Google Scholar] [CrossRef]
- Scheiermann, P.; Bachmann, M.; Härdle, L.; Pleli, T.; Piiper, A.; Zwissler, B.; Pfeilschifter, J.; Mühl, H. Application of IL-36 receptor n-tagonist weakens CCL20 expression and impairs recovery in the late phase of murine acetaminophen-induced liver injury. Sci. Rep. 2015, 5, 8521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensen, J.T.; Dawson, P.A.; Mychaleckyj, J.C.; Bowden, D.W. Identification of a novel human cytokine gene in the interleukin gene cluster on chromosome 2q12-14. J. Interferon Cytokine Res. 2001, 21, 899–904. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, D.M.; Teufel, L.U.; van de Veerdonk, F.L.; Joosten, L.A.; Netea, M.G.; Dinarello, C.A.; Arts, R.J.W. IL-38 prevents induc-tion of trained immunity by inhibition of mTOR signaling. J. Leukocyte Biol. 2021, 3, 23. [Google Scholar]
- Pfeiler, S.; Winkels, H.; Kelm, M.; Gerdes, N. IL-1 family cytokines in cardiovascular disease. Cytokine 2019, 122, 154215. [Google Scholar] [CrossRef]
- Balato, A.; Schiattarella, M.; Lembo, S.; Mattii, M.; Prevete, N.; Balato, N.; Ayala, F. Interleukin-1 family members are enhanced in psoriasis and suppressed by vitamin D and retinoic acid. Musculoskelet. Dermatol. Res. 2013, 305, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Chen, Z.; Huang, N.; Teng, X.; Zhang, J.; Wang, Z.; Wei, X.; Qin, K.; Liu, X.; Wu, X.; et al. Expression, purification of IL-38 in Escherichia coli and production of polyclonal antibodies. Protein Expr. Purif. 2015, 107, 76–82. [Google Scholar] [CrossRef]
- Noorani, M.; Lakhani, N. Kawasaki disease: Two case reports from the Aga Khan Hospital, Dar es Salaam-Tanzania. BMC Pediatr. 2018, 18, 334. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Xu, Y.; Gao, X.; Li, Y.; Li, M.; Wang, Y.; Gao, Q. Cloning and Sequence Analysis of Human Inter-leukin-38 Gene. CNKI 2021, 7, 20099. (In Chinese) [Google Scholar]
- Boutet, M.-A.; Nerviani, A.; Pitzalis, C. IL-36, IL-37, and IL-38 Cytokines in Skin and Joint Inflammation: A Comprehensive Review of Their Therapeutic Potential. Inter. J. Mol. Sci. 2019, 20, 1257. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-SWJS201302003.htm (accessed on 15 August 2021). [CrossRef] [Green Version]
- Clavel, G.; Thiolat, A.; Boissier, M.-C. Interleukin newcomers creating new numbers in rheumatology: IL-34 to IL-38. Jt. Bone Spine 2013, 80, 449–453. [Google Scholar] [CrossRef]
- Chu, M.; Chu, I.M.; Yung, E.C.; Lam, C.W.; Leung, T.F.; Wong, G.W.; Wong, C.K. Aberrant Expression of Novel Cytokine IL-38 and Regulatory T Lymphocytes in Childhood Asthma. Molecules 2016, 21, 933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, R.; Chen, J.; Jin, J.; Yu, Y.; Tian, Y.; Li, W.; Wang, W.; Zhou, H.; Su, S.B. The Effect of Interleukin 38 on Angiogenesis in a Model of Oxygen-induced Retinopathy. Sci. Rep. 2017, 7, 2756. [Google Scholar] [CrossRef] [Green Version]
- Rudloff, I.; Godsell, J.; Nold-Petry, C.A.; Harris, J.; Hoi, A.; Morand, E.F.; Nold, M.F. Brief Report: Interleukin-38 Exerts Anti-inflammatory Functions and Is Associated with Disease Activity in Systemic Lupus Erythematosus. Arthritis Rheumatol. 2015, 67, 3219–3225. [Google Scholar] [CrossRef] [PubMed]
- Zdravkovic, N.; Rosic, M.; Lutovac, M.; Zdravkovic, V. Physiology and Pathology of Cytokine: Commercial Production and Medical Use. Physiol. Pathol. Immunol. 2017, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Weigert, A.; Javier, M.; Brüne, B.; Dillmann, C.; Parnham, M.J.; Geisslinger, G. N-Terminally Truncated Interleukin-38. Eur. Patents EP2977384A1, 27 January 2016. [Google Scholar]
- Yuan, X.; Peng, X.; Li, Y.; Li, M. Role of IL-38 and Its Related Cytokines in Inflammation. Mediat. Inflamm. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mora, J.; Weigert, A. IL-1 family cytokines in cancer immunity–a matter of life and death. Biol. Chem. 2016, 397, 1125–1134. [Google Scholar] [CrossRef]
- Xia, H.; Liu, Y.; Fu, Y.; Li, M.; Wu, Y. Biology of interleukin-38 and its role in chronic inflammatory diseases. Int. Immunopharmacol. 2021, 95, 107528. [Google Scholar] [CrossRef]
- Xie, C.; Yan, W.; Quan, R.; Chen, C.; Tu, L.; Hou, X.; Fu, Y. Interleukin-38 is elevated in inflammatory bowel diseases and suppresses intestinal inflammation. Cytokine 2020, 127, 154963. [Google Scholar] [CrossRef]
- El-Esawy, F.; Elhabak, D.; Mohamed, S.; Mostafa, S. Evaluating the Role of Interleukin-36 and Interleukin-38 in Acne Vulgaris Patients. Benha J. Appl. Sci. 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Zarrabi, M.; Gholijani, N.; Shenavandeh, S.; Aflaki, E.; Amirghofran, Z. IL-38 serum levels in patients with Behcet’s disease and the relationship with clinical features. Eur. Cytokine Netk. 2019, 30, 82–87. [Google Scholar]
- Hatemi, G.; Seyahi, E.; Fresko, I.; Talarico, R.; Hamuryudan, V. One year in review 2020: Behçet’s syndrome. Clin. Exp. Rheumatol. 2020, 38, S3–S10. [Google Scholar]
- Yu, H.; Liu, Y.; Xie, W.; Xie, Q.; Liu, Q.; Cheng, L. IL-38 alleviates the inflammatory response and the degeneration of nucleus pulposus cells via inhibition of the NF-κB signaling pathway in vitro. Int. Immunopharmacol. 2020, 85, 106592. [Google Scholar] [CrossRef] [PubMed]
- Yahya, A.M.; Jasim, R.Z. Interleukin-38 (IL-38) is A Novel Biochemical Marker in Sera of Iraqi Patients with Alzheimers Disease. Biochem. Cell. Arch. 2021, 21, 1579–1584. [Google Scholar]
- Rafie, M.Z.; Esmaeilzadeh, A.; Ghoreishi, A.; Tahmasebi, S.; Faghihzadeh, E.; Elahi, R. IL-38 as an early predictor of the ischemic stroke prognosis. Cytokine 2021, 146, 155626. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Pantazopoulos, H.; Conti, P.; Leeman, S.E.; Theoharides, T.C. IL-38 inhibits microglial inflammatory me-diators and is decreased in amygdala of children with autism spectrum disorder. Proc. Natl. Acad. Sci. USA 2020, 117, 16475–16480. [Google Scholar] [CrossRef]
- Shi, L.; Ye, H.; Huang, J.; Li, Y.; Wang, X.; Xu, Z.; Chen, J.; Xiao, W.; Chen, R.; Yang, H. IL-38 Exerts Anti-Inflammatory and Antifibrotic Effects in Thyroid-Associated Ophthalmopathy. J. Clin. Endocrinol. Metab. 2021, 106, e3125–e3142. [Google Scholar] [CrossRef]
- Zarrabi, M.; Nazarinia, M.; Jaberi, A.R.; Gholijani, N.; Amirghofran, Z. Elevated IL-38 Serum Levels in Newly Diagnosed Multiple Sclerosis and Systemic Sclerosis Patients. Med. Princ. Pract. 2021, 29, 514–521. [Google Scholar]
- Huard, A.; Do, H.N.; Frank, A.-C.; Sirait-Fischer, E.; Fuhrmann, D.; Hofmann, M.C.J.; Raue, R.; Palmer, G.; Brüne, B.; de Bruin, N.; et al. IL-38 Ablation Reduces Local Inflammation and Disease Severity in Experimental Autoimmune Encephalomyelitis. J. Immunol. 2021, 206, 1058–1066. [Google Scholar] [CrossRef]
- Veerdonk, F.V.D.; Netea, M. New insights in the immunobiology of IL-1 family members. Front. Immunol. 2013, 4, 167. [Google Scholar] [CrossRef] [Green Version]
- Ummarino, D. Experimental arthritis: IL-38 promotes anti-inflammatory effects. Nat. Rev. Rheumatol. 2017, 13, 260. [Google Scholar] [CrossRef]
- Zhang, L.; Yan, J.-W.; Wang, Y.-J.; Wan, Y.-N.; Wang, B.-X.; Tao, J.-H.; Chen, B.; Li, B.-Z.; Yang, G.-J.; Wang, J. Association of interleukin 1 family with systemic sclerosis. Inflammation 2014, 37, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Khattab, F.M.; Khater, E.M.; Ibraheem, H.A. Serum levels of interleukin-38 in sufferers with atopic eczema. Egypt. J. Dermatol. Venerol. 2019, 39, 66. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, S.; Yang, Z. IL-38 inhibits osteoporosis via regulating PI3K/Akt/GSK3β/NFATc1 signaling pathway. Chin. J. Immunol. 2018, 34, 251–255. [Google Scholar]
- Sun, X.; Hou, T.; Cheung, E.; Iu, T.N.-T.; Tam, V.W.-H.; Chu, I.M.-T.; Tsang, M.S.-M.; Chan, P.K.-S.; Lam, C.W.-K.; Wong, C.-K. Anti-inflammatory mechanisms of the novel cytokine interleukin-38 in allergic asthma. Cell. Mol. Immunol. 2020, 17, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Z.P.M.; Ghafouri-Fard, S.; Komaki, A.; Mazdeh, M.; Taheri, M.; Eftekharian, M.M. Assessment of IL-38 Levels in Patients with Acquired Immune-Mediated Polyneuropathies. J. Mol. Neurosci. 2020, 70, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Tsang, M.S.-M.; Sun, X.; Wong, C.K. The Role of New IL-1 Family Members (IL-36 and IL-38) in Atopic Dermatitis, Allergic Asthma, and Allergic Rhinitis. Curr. Allergy Asthma Rep. 2020, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hiz, P.; Kanbur, E.; Demir, N.; Akalin, H.; Cagan, E.; Pashazadeh, M.; Bal, S.H.; Tezcan, G.; Oral, H.B.; Budak, F. Roles of novel IL-1 family (IL-36, IL-37, and IL-38) members in chronic brucellosis. Cytokine 2020, 135, 155211. [Google Scholar] [CrossRef]
- Hahn, M.; Frey, S.; Hueber, A. The novel interleukin-1 cytokine family members in inflammatory diseases. Curr. Opin. Rheumatol. 2017, 29, 208–213. [Google Scholar] [CrossRef]
- Angelotti, F.; Parma, A.; Cafaro, G.; Capecchi, R.; Alunno, A.; Puxeddu, I. One year in review 2017: Pathogenesis of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 368–378. [Google Scholar]
- Boutet, M.-A.; Najm, A.; Bart, G.; Brion, R.; Touchais, S.; Trichet, V.; Layrolle, P.; Gabay, C.; Palmer, G.; Blanchard, F.; et al. IL-38 overexpression induces an-ti-inflammatory effects in mice arthritis models and in human macrophages in vitro. Ann. Rheum. Dis. 2017, 76, 1304–1312. [Google Scholar] [CrossRef]
- Boutet, M.-A.; Blanchard, F.; Le Goff, B. Response to:‘Does IL-38 act on macrophages and/or dendritic cells in ar-thritis?’by Jiang et al. Ann. Rheum. Dis. 2018, 77, e13. [Google Scholar] [CrossRef] [PubMed]
- Rudd, P.A.; Herrero, L.J. Viruses: Friends and Foes. Cartil. Repair Regen. 2018, 8, 824. [Google Scholar] [CrossRef] [Green Version]
- Harel, M.; Garraud, T.; Le Goff, B.; Blanchard, F. P060 Il-38 in arthritis maturation and degradation of this new IL-1 family anti-inflammatory cytokine. Poster Present. 2018, 77, A38–A39. [Google Scholar] [CrossRef] [Green Version]
- Pei, B.; Chen, K.; Zhou, S.; Min, D.; Xiao, W. IL-38 restrains inflammatory response of collagen-induced arthritis in rats via SIRT1/HIF-1α signaling pathway. Biosci. Rep. 2020, 40, 2431. [Google Scholar] [CrossRef]
- Joosten, L.; Dinarello, C. Biology of IL-38 and its role in disease. Immunol. Rev. 2018, 281, 191–196. [Google Scholar]
- Fattori, V.; Zarpelon, A.C.; Staurengo-Ferrari, L.; Borghi, S.M.; Zaninelli, T.H.; Da Costa, F.B.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; Casagrande, R.; et al. Budlein A, a Sesquiterpene Lactone from Viguiera robusta, Alleviates Pain and Inflammation in a Model of Acute Gout Arthritis in Mice. Front. Pharmacol. 2018, 9, 1076. [Google Scholar] [CrossRef] [Green Version]
- Hao, Z.; Juan, N.; Wang, H. Mechanism of IL-38 inhibits LPS/TLR4 induced inflammation in patients with rheumatoid arthritis. Chin. J. Immunol. 2017, 33, 1647–1651. [Google Scholar]
- Zhang, X.; He, T.; Ling, J.; Hu, X.; Wen, Y.; Wei, J.; Lu, H.L. Serum levels of interleukin-38 and interleukin-1β in the acute phase of Kawasaki disease in children. Zhongguo Dang Dai Er Ke Za Zhi 2018, 20, 543–548. (In Chinese) [Google Scholar]
- Takenaka, S.; Kaieda, S.; Kawayama, T.; Matsuoka, M.; Kaku, Y.; Kinoshita, T.; Sakazaki, T.; Okamoto, M.; Tominaga, M.; Kanesaki, K.; et al. IL-38: A new factor in rheuma-toid arthritis. Biochem. Biophys. Rep. 2015, 4, 386–391. [Google Scholar]
- Xu, W.-D.; Su, L.-C.; He, C.-S.; Huang, A.-F. Plasma interleukin-38 in patients with rheumatoid arthritis. Int. Immunopharmacol. 2018, 65, 1–7. [Google Scholar] [CrossRef]
- Jiang, L.; Zhou, X.; Huang, C.; Bao, J.; Li, J.; Xu, K.; Dong, D.; Wu, L. The elevated expression of IL-38 serves as an an-ti-inflammatory factor in osteoarthritis and its protective effect in osteoarthritic chondrocytes. Int. Immunopharmacol. 2021, 94, 107489. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, D.M.; Maas, R.J.; Smeekens, S.P.; Eisenmesser, E.; Redzic, J.S.; Helsen, M.M.; Powers, N.E.; Li, S.; Kalabokis, V.; Gresnigt, M.S.; et al. Human recombinant inter-leukin-38 suppresses inflammation in mouse models of local and systemic disease. Cytokine 2021, 137, 155334. [Google Scholar] [CrossRef] [PubMed]
- Abassifard, M.; Khorramdelazad, H.; Rezaee, S.; Jafarzadeh, A. Higher Circulating Concentration of Interleukin-38 in Patients with Knee Osteoarthritis: Its Association with Disease Severity. Iran. J. Allergy Asthma Immunol. 2021, 20, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, Z.; Akbarian, M.; Mirkazemi, S.; Shahlaee, A.; Alizadeh, Z.; Amirzargar, A.A.; Jamshidi, A.R.; Ghoroghi, S.; Poursani, S.; Nourijelyani, K.; et al. Interleukin-1 gene cluster and IL-1 receptor polymorphisms in Iranian patients with systemic lupus erythematosus. Rheumatol. Int. 2013, 33, 2591–2596. [Google Scholar] [CrossRef]
- Godsell, J.; Rudloff, I.; Harris, J.; Hoi, A.; Nold, M.; Morand, E. First Report of Il-38 In Systemic Lupus Erythematosus (sle): Ara8. Int. Med. J. 2014, 44, 3. [Google Scholar]
- Takeuchi, Y.; Seki, T.; Kobayashi, N.; Sano, K.; Shigemura, T.; Shimojo, H.; Matsumoto, K.; Agematsu, K. Analysis of serum IL-38 in juve-nile-onset systemic lupus erythematosus. Modern Rheum. 2018, 28, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Rudloff, I.; Godsell, J.; Nold-Petry, C.A.; Harris, J.; Hoi, A.; Morand, E.F.; Nold, M.F. Serum interleukin 38 is associated with disease severity and organ involvement in systemic lupus erythematosus. Eur. J. Immunol. 2016, 46, 41. [Google Scholar]
- Chu, M.; Tam, L.-S.; Zhu, J.; Jiao, D.; Liu, D.H.; Cai, Z.; Dong, J.; Lam, C.W.K.; Wong, C.K. In vivo anti-inflammatory activities of novel cytokine IL-38 in Murphy Roths Large (MRL)/lpr mice. Immunobiology 2017, 222, 483–493. [Google Scholar] [CrossRef]
- Han, Y.; Mora, J.; Huard, A.; da Silva, P.; Wiechmann, S.; Putyrski, M.; Schuster, C.; Elwakeel, E.; Lang, G.; Scholz, A.; et al. IL-38 Ameliorates Skin Inflammation and Limits IL-17 Production from γδ T Cells. Cell Rep. 2019, 27, 835–846.e5. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Liu, J.; Zhang, R.; Huang, X.; Sun, T.; Wu, Y.; Hambly, B.; Bao, S. IL-37 and 38 signalling in gestational diabetes. J. Reprod. Immunol. 2017, 124, 8–14. [Google Scholar] [CrossRef]
- Mermoud, L.; Talabot-Ayer, D.; Brembilla, N.; Borowczyk-Michalowska, J.J.; Palmer, G. P069 Constitutive overex-pression of interleukin 38 has a negative impact on human NHK keratinocyte fitness. Ann. Rheum. Dis. 2019, 78, A29. Available online: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=Serum+interleukin+38+is+associated+with+disease+severity+and+organ+involvement+in+systemic+lupus+erythematosus.+%E2%80%8E&btnG (accessed on 15 August 2021).
- Conti, P.; Pregliasco, F.; Bellomo, R.G.; Gallenga, C.E.; Caraffa, A.; Kritas, S.K.; Lauritano, D.; Ronconi, G. Mast Cells Mediate Skin Inflamma-tion in Psoriasis: A Novel Therapeutic Approach with the Anti-inflammatory Cytokines IL-37, IL-38 and IL-1Ra. Preprints 2021, 10, 520. [Google Scholar]
- Wu, Y.; Li, H.; Jiang, Z.; Lai, Y. The interleukin-1 family: A key regulator in the pathogenesis of psoriasis. Austin J. Clin. Immunol. 2014, 5, 1023. [Google Scholar]
- Mercurio, L.; Morelli, M.; Scarponi, C.; Eisenmesser, E.Z.; Doti, N.; Pagnanelli, G.; Gubinelli, E.; Mazzanti, C.; Cavani, A.; Ruvo, M.; et al. IL-38 has an anti-inflammatory action in psoriasis and its expression correlates with disease severity and therapeutic response to anti-IL-17A treatment. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Keermann, M.; Koks, S.; Reimann, E.; Abram, K.; Erm, T.; Silm, H.; Kingo, K. Expression of IL-36 family cytokines and IL-37 but not IL-38 is altered in psoriatic skin. J. Dermatol. Sci. 2015, 80, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Boutet, M.-A.; Bart, G.; Penhoat, M.; Amiaud, J.; Brulin, B.; Charrier, C.; Morel, F.; Lecron, J.; Rolliderkinderen, M.; Bourreille, A.; et al. Distinct expression of interleukin (IL)-36α, βandγ, their antagonist IL-36Ra and IL-38 in psoriasis, rheumatoid arthritis and Crohn’s disease. Clin. Exp. Immunol. 2016, 184, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Palomo, J.; Troccaz, S.; Rodriguez, E.; Talabot-Ayer, D.; Gabay, C.; Palmer, G. 02.11 Il-38 is not involved in the modulation of imq-induced skin inflammation. Ann. Rheum. Dis. 2017, 76, A12. [Google Scholar]
- Lauritano, D.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Di Emidio, P.; Martinotti, S.; Tetè, G.; Ross, R.; Conti, P. New aspect of allergic contact dermatitis, an inflammatory skin disorder mediated by mast cells: Can IL-38 help? Med. Hypotheses 2020, 139, 109687. [Google Scholar] [CrossRef]
- Zhou, Q.; Lin, F.F.; Liu, S.M.; Sui, X.F. Influence of the total saponin fraction from Dioscorea nipponica Makino on TLR2/4-IL1R receptor singnal pathway in rats of gouty arthritis. J. Ethnopharmacol. 2017, 206, 274–282. [Google Scholar] [CrossRef]
- Wei, W.; Syed Sayeed, A.; Shuang, C.; Yu, X.; Mohammad Amjad, K.; Jiang, L. From Molecular Mechanism to the Etiology of Sjogren Syndrome. Curr. Pharm. Des. 2018, 24, 1–9. [Google Scholar] [CrossRef]
- Ciccia, F.; Accardo-Palumbo, A.; Alessandro, R.; Alessandri, C.; Priori, R.; Guggino, G.; Raimondo, S.; Carubbi, F.; Valesini, G.; Giacomelli, R.; et al. Interleukin-36α axis is modulated in patients with primary Sjögren’s syndrome. Clin. Exp. Immunol. 2015, 181, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampeli, E.; Kalogirou, E.-M.; Piperi, E.; Mavragani, C.P.; Moutsopoulos, H.M. Tongue Atrophy in Sjögren Syndrome Patients with Mucosa-associated Lymphoid Tissue Lymphoma: Autoimmune Epithelitis beyond the Epitheli-al Cells of Salivary Glands? J. Rheumatol. 2018, 45, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Chen, Y.; Zhou, N.; Li, T.; Wang, H. Blockade of Th17 response by IL-38 in primary Sjögren’s syndrome. Mol. Immunol. 2020, 127, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Dige, A.; Magnusson, M.K.; Uhrenholt, C.; Rasmussen, T.K.; Kragstrup, T.; Ohman, L.; Dahlerup, J.; Agnholt, J. Effects of Anti-TNFalpha Treatment on Mucosal Expression of IL-17A, IL-21, and IL-22 and Cytokine-Producing T Cell Subsets in Crohn’s Disease. Mediators Inflamm. 2018, 26, 3279607. [Google Scholar]
- Fonseca-Camarillo, G.; Iturriaga-Goyon, E.; Furuzawa-Carballeda, J.; Yamamoto-Furusho, J.K. Role of IL-38 and its Antagonist in Patients with Inflammatory Bowel Disease. Gastroenterology 2017, 152, S762. [Google Scholar] [CrossRef]
- Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Iturriaga-Goyon, E.; Yamamoto-Furusho, J.K. P042 Expression of IL-38 and their antagonists in patients with inflammatory bowel disease. J. Crohn’s Colitis 2017, 11, S99. [Google Scholar] [CrossRef] [Green Version]
- Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Iturriaga-Goyon, E.; Yamamoto-Furusho, J.K. Differential Expres-sion of IL-36 Family Members and IL-38 by Immune and Nonimmune Cells in Patients with Active Inflam-matory Bowel Disease. BioMed Res. Int. 2018, 2018, 5140691. [Google Scholar] [CrossRef]
- Zhao, Y.; Mei, H.; Peng, H.; Gao, Y.; Chen, J. Molecular mechanisms of interleukin-38 inhibiting inflammatory bowel disease in children by regulating nuclear factor-κB and signal transduction and activator of transcrip-tion 3 pathway. Chin. J. Digesion. 2019, 15, 237–243. [Google Scholar]
- Zhao, B.; Chen, W.; Jiang, R.; Zhang, R.; Wang, Y.; Wang, L.; Gordon, L.; Chen, L. Expression profile of IL-1 family cytokines in aqueous humor and sera of patients with HLA-B27 associated anterior uveitis and idiopathic anterior uveitis. Exp. Eye Res. 2015, 138, 80–86. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, J.; Wang, Y.; Chen, J.; Li, X.; Liu, X.; Kong, E.; Su, S.B.; Zhang, Z. The Effect of Interleukin 38 on Inflammation-induced Corneal Neovascularization. Curr. Mol. Med. 2019, 19, 589–596. [Google Scholar] [CrossRef]
- Hessam, S.; Sand, M.; Gambichler, T.; Skrygan, M.; Rüddel, I.; Bechara, F. Interleukin-36 in hidradenitis suppurativa: Evidence for a distinctive proinflammatory role and a key factor in the development of an inflammatory loop. Br. J. Dermatol. 2018, 178, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo Villavicencio, J.C.; Agudelo Arbelaez, N.; Lastra, B.R.; Ramirez, I.; Quezada, F.; Ponte, M.C.; Cañola, L.; Smith, J.M. Primary Outcomes of Patients with Chronic Angle-Closure Glaucoma Treated with Combined Phacoemulsification, Viscogoniosynechialysis, and Endocyclophotocoagulation. J. Ophthalmol. 2019, 2019, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnadas, R. Current management options in primary angle closure disease. Indian J. Ophthalmol. 2019, 67, 321. [Google Scholar] [CrossRef]
- Zhang, J.; Song, X.; Chen, Y.; Nguyen, T.H.A.; Zhang, J.; Zhang, Y. Novel inflammatory cytokines (IL-36, 37, 38) in the aqueous humor from patients with chronic primary angle closure glaucoma. Int. Immunopharmacol. 2019, 71, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Fruttiger, M. Oxygen-induced retinopathy: A model for vascular pathology in the retina. Eye 2010, 24, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.-D.; Huang, A.-F. Role of Interleukin-38 in Chronic Inflammatory Diseases: A Comprehensive Review. Front. Immunol. 2018, 9, 1462. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Ganeshkumar, P.; Prakash, M.V.S.; Christopher, B.J.; Amalraj, V. Prevalence of overweight/obesity among the medical students, Malaysia. Med. J. Malays. 2012, 67, 442–444. [Google Scholar]
- Xu, K.; Sun, J.; Chen, S.; Li, Y.; Peng, X.; Li, M. Hydrodynamic delivery of IL-38 gene alleviates obesity-induced inflammation and insulin resistance. Biochem. Biophys. Res. Commun. 2019, 508, 198–202. [Google Scholar] [CrossRef]
- Malone, J.I.; Hansen, B.C. Does obesity cause type 2 diabetes mellitus (T2DM)? Or is it the opposite? Pediatr. Diabetes 2018, 20, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Pickup, J.C. Inflammation and Activated Innate Immunity in the Pathogenesis of Type 2 Diabetes. Diabetes Care 2004, 27, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Ballak, D.B.; Stienstra, R.; Tack, C.J.; Dinarello, C.A.; van Diepen, J.A. IL-1 family members in the pathogenesis and treatment of metabolic disease: Focus on adipose tissue inflammation and insulin resistance. Cytokine 2015, 75, 280–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurău, F.; Silvestrini, A.; Matacchione, G.; Fazioli, F.; Bonfigli, A.R.; Olivieri, F.; Sabbatinelli, J. Plasma levels of interleukin-38 in healthy aging and in type 2 diabetes. Diabetes Res. Clin. Pr. 2021, 171, 108585. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, S.; Sun, J.; Yu, Y.; Li, M. Interleukin-38 inhibits adipogenesis and inflammatory cytokine production in 3T3-L1 preadipocytes. Cell Biol. Int. 2020, 44, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, T.; Zhou, F.; Mu, D.; Liu, S. Interleukin-38 increases the insulin sensitivity in children with the type 2 diabetes. Int. Immunopharmacol. 2020, 82, 106264. [Google Scholar] [CrossRef]
- Abid, S.M.; Alaaraji, S.F.; Alrawi, K.F. Study of Interleukins 13, 18, 27, 32 and 38 Levels in Iraqi Type 2 diabetes with Insulin Resistance. J. Educ. Sci. Stud. 2019, 4, 14. [Google Scholar]
- Wang, C.; Guelfi, K.; Yang, H.-X. Exercise and its role in gestational diabetes mellitus. Chronic Dis. Transl. Med. 2016, 2, 208–214. [Google Scholar] [CrossRef]
- Southcombe, J.H.; Redman, C.W.G.; Sargent, I.L.; Granne, I. Interleukin-1 family cytokines and their regulatory proteins in normal pregnancy and pre-eclampsia. Clin. Exp. Immunol. 2015, 181, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhang, W.; Liu, X.; Zhang, W.; Li, Y. Interrelationship between diabetes and periodontitis: Role of hyperlipidemia. Arch. Oral Biol. 2015, 60, 667–674. [Google Scholar] [CrossRef]
- Yang, N.; Song, Y.; Dong, B.; Li, Y.; Kou, L.; Yang, J.; Qin, Q. Elevated Interleukin-38 Level Associates with Clinical Re-sponse to Atorvastatin in Patients with Hyperlipidemia. Cell. Physiol. Biochem. 2018, 49, 653–661. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The inflammatory response in myocardial injury, repair, and remodelling. Nat. Rev. Cardiol. 2014, 11, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Yu, K.; Wang, X.; Wang, X.; Ji, Q.; Zeng, Q. Elevated Plasma IL-38 Concentrations in Patients with Acute ST-Segment Elevation Myocardial Infarction and Their Dynamics after Reperfusion Treatment. Mediat. Inflamm. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Lan, Y.; Zhong, Y.; Yu, K.; Xu, W.; Zhu, R.; Sun, H.; Ding, Y.; Wang, Y.; Zeng, Q. Interleukin-38 alleviates cardiac remodelling after myocardial infarction. J. Cell. Mol. Med. 2021, 24, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Graaf, D.M.; Jaeger, M.; van den Munckhof, I.C.; Ter Horst, R.; Schraa, K.; Zwaag, J.; Kox, M.; Fujita, M.; Yamauchi, T.; Mercurio, L.; et al. Reduced concentrations of the B cell cytokine interleukin 38 are associated with cardiovascular disease risk in overweight subjects. Eur. J. Immunol. 2021, 51, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Fan, J.; Zhang, X.; Huang, Q.; Han, J.; Wu, F.; Hu, G.; Guo, M.; Jin, Y. Integrative genomic analysis of interleukin-36RN and its prognostic value in cancer. Mol. Med. Rep. 2016, 13, 1404–1412. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Huard, A.; Mora, J.; da Silva, P.; Brüne, B.; Weigert, A. IL-36 family cytokines in protective versus destructive inflammation. Cell. Signal. 2020, 75, 109773. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, W.; Wu, T.; Chu, H. Reduced interleukin-38 in non-small cell lung cancer is associated with tumour progression. Royal Soc. Open Biol. 2018, 8, 180132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Zhang, F.; Tan, Z.; Hambly, B.D.; Bao, S.; Tao, K. Interleukin-38 in colorectal cancer: A potential role in precision medicine. Cancer Immunol. Immunother. 2020, 69, 69–79. [Google Scholar] [CrossRef]
- Bao, S.; Hu, R.; Hambly, B.D. IL-34, IL-36 and IL-38 in colorectal cancer—Key immunoregulators of carcinogenesis. Biophys. Rev. 2020, 12, 925–930. [Google Scholar] [CrossRef]
- Tominaga, M.; Okamoto, M.; Kawayama, T.; Matsuoka, M.; Kaieda, S.; Sakazaki, Y.; Kinoshita, T.; Mori, D.; Inoue, A.; Hoshino, T. Overexpression of IL-38 protein in anticancer drug-induced lung injury and acute exacerbation of idiopathic pulmonary fibrosis. Respir. Investig. 2017, 55, 293–299. [Google Scholar] [CrossRef]
- Xu, Z.; Yuan, X.; Gao, Q.; Li, Y.; Li, M. Interleukin-38 overexpression prevents bleomycin-induced mouse pulmo-nary fibrosis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 391–399. [Google Scholar] [CrossRef]
- Sun, Y.-H.; Liu, Z.-X.; Bao, W.-H.; Wang, Y.-X.; Niu, Y.-Y. Role of IL-38 and MIP-2 in pulmonary fibrosis in rats. Chin. J. Immunol. 2019, 12, 151–155. [Google Scholar]
- Gao, X.; Chan, P.K.S.; Lui, G.C.Y.; Hui, D.S.C.; Chu, I.M.-T.; Sun, X.; Tsang, M.S.-M.; Chan, B.C.L.; Lam, C.W.-K.; Wong, C.-K. Interleukin-38 ameliorates poly(I:C) induced lung inflammation: Therapeutic implications in respiratory viral infections. Cell Death Dis. 2021, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Conti, P. COVID-19 induced by SARS-CoV-2 causes Kawasaki-like disease in children: Role of pro-inflammatory and anti-inflammatory cytokines. J. Biol. Regul. Homeost. Agents. 2020, 34, 767–773. [Google Scholar]
- Yalcin, A.D.; Yalcin, A.N. Future perspective: Biologic agents in patients with severe COVID-19. Immunopharmacol. Immunotoxicol. 2021, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Takada, K.; Okamoto, T.; Tominaga, M.; Teraishi, K.; Akamine, T.; Takamori, S.; Katsura, M.; Toyokawa, G.; Shoji, F.; Okamoto, M.; et al. Clinical implications of the novel cytokine IL-38 expressed in lung adenocarcinoma: Possible association with PD-L1 expression. PLoS ONE 2017, 12, e0181598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anborgh, P.H.; Kolotilin, I.; Owens, N.; Mahboob, A.A. Prolific Induction of IL-6 in Human Cells by SARS-CoV-2-derived Peptide is Attenuated by Recombinant Human Anti-inflammatory Cytokines made in planta. bioRxiv. 2021. [Google Scholar] [CrossRef]
- Kinoshita, F.; Tagawa, T.; Akamine, T.; Takada, K.; Yamada, Y.; Oku, Y.; Kosai, K.; Ono, Y.; Tanaka, K.; Wakasu, S.; et al. Interleukin-38 promotes tumor growth through regulation of CD8+ tumor-infiltrating lymphocytes in lung cancer tumor microenvironment. Cancer Immunol. Immunother. 2021, 70, 123–135. [Google Scholar] [CrossRef]
- Zhu, Y.Q.; Ma, S.P.; Li, B.; Zheng, C.Y.; Ma, J. [Interleukin-38 expression and clinical significance in serum of patients with chronic obstructive pulmonary disease]. Zhonghua Yi Xue Za Zhi 2018, 98, 759–762. (In Chinese) [Google Scholar] [CrossRef]
- Wei, H.D.; Hong, J.; De, S.L. Correlation Analysis of Serum Galectin-3, IL-38 Levels with Pulmonary Func-tion in Patients with Chronic Obstructive Pulmonary Disease. J. Clin. Transfus. Lab. Med. 2020, 22, 152. [Google Scholar]
- Kinoshita, T.; Sakazaki, Y.; Matsuoka, M.; Tokunaga, Y.; Tominaga, M.; Kaieda, S.; Okamoto, M.; Kawayama, T.; Hoshino, T. The Role of Interleukin-38 In Elastase-Induced Mouse Model. B80-A Mechanisms and Models of Acute Lung Injury; American Thoracic Society: New York, NY, USA, 2017; p. A4410-A. [Google Scholar]
- Matsuoka, M.; Kawayama, T.; Tominaga, M.; Kaieda, S.; Tokunaga, Y.; Kaku, Y.; Imaoka, H.; Kinoshita, T.; Okamoto, T.; Akiba, J.; et al. Attenuated Air-way Eosinophilic Inflammations in IL-38 Knockout Mouse Model. Kurume Med. J. 2019, 65, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, M.; Kawayama, T.; Tominaga, M.; Kaieda, S.; Tokunaga, Y.; Kaku, Y.; Imaoka, H.; Kinoshita, T.; Okamoto, M.; Hoshino, T. Il-38 May Enhance Ova-Induced Eosinophilic Inflammation in Mice. In C35 Asthma and Allergy Cellular Investigations; American Thoracic Society: New York, NY, USA, 2017; p. A5308-A. [Google Scholar]
- Zhu, Y.; Ma, S.; Dong, W.; Zhang, J.; Zhang, Y. Correlation between serum interleukin-38 and acute exacerba-tion of chronic obstructive pulmonary disease with pulmonary embolism. Zhonghua Yi Xue Za Zhi 2019, 99, 929–933. (In Chinese) [Google Scholar] [PubMed]
- Richeldi, L.; Launders, N.; Martinez, F.; Walsh, S.L.; Myers, J.; David-Wang, A.; Jones, M.; Chisholm, A.; Flaherty, K.R. The characterisation of interstitial lung disease multidisciplinary team meetings: A global study. ERJ Open Res. 2019, 5, 00209–02018. [Google Scholar] [CrossRef] [Green Version]
- Cottin, V.; Hirani, N.A.; Hotchkin, D.L.; Nambiar, A.M.; Ogura, T.; Otaola, M.; Skowasch, D.; Park, J.S.; Poonyagariyagorn, H.K.; Wuyts, W.; et al. Presentation, diagnosis and clinical course of the spectrum of progressive-fibrosing interstitial lung diseases. Eur. Respir. Rev. 2018, 27, 180076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heukels, P.; Moor, C.; von der Thusen, J.; Wijsenbeek, M.; Kool, M. Inflammation and immunity in IPF pathogenesis and treatment. Respir. Med. 2019, 147, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Okamoto, M.; Kawayama, T.; Kaieda, S.; Matsuoka, M.; Sakazaki, Y.; Kinoshita, T.; Hoshino, T. Overexpression of Il-38 protein in the Lungs of drug induced Lung Injury. In C67 Acute Lung Injury; American Thoracic Society: New York, NY, USA, 2016; p. A5786-A. [Google Scholar]
- Tominaga, M.; Okamoto, M.; Kinoshita, T.; Sakazaki, Y.; Matsuoka, M.; Kaieda, S.; Hoshino, T. New IL-1 family IL-38 is highly expressed in alveolar cells of drug-induced lung injury and idiopathic pulmonary fibrosis. Eur. Respir. J. 2016, 48, PA3088. [Google Scholar] [CrossRef]
- Chai, Y.; Lin, S.; Zhang, M.; Deng, L.; Chen, Y.; Xie, K.; Wang, C.J.; Xu, F. IL-38 is a biomarker for acute respiratory distress syn-drome in humans and down-regulates Th17 differentiation in vivo. Clin. Immunol. 2019, 210, 108315. [Google Scholar] [CrossRef] [PubMed]
- Terrault, N.A.; Lok, A.S.; McMahon, B.J.; Chang, K.-M.; Hwang, J.; Jonas, M.M.; Brown, R.S., Jr.; Bzowej, N.H.; Wong, J.B. Update on prevention, diagnosis, and treatment of chronic hepatitis B: AASLD 2018 hepatitis B guidance. Hepatology 2018, 67, 1560–1599. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-J.; Jiang, Y.-F.; Wang, X.-R.; Zhang, M.-L.; Gao, P.-J. Elevated serum interleukin-38 level at baseline predicts virological response in telbivudine-treated patients with chronic hepatitis B. World J. Gastroenterol. 2016, 22, 4529–4537. [Google Scholar] [CrossRef] [PubMed]
- Alaaraji, S.F.T. Exploration of the Relationship between Interleukins 17, 37 and 38 with Vitamin E in Iraqi Men with CHB. J. Phys. Ser. 2019, 1294, 052047. [Google Scholar] [CrossRef]
- Yuan, X.; Li, Y.; Pan, X.; Peng, X.; Song, G.; Jiang, W.; Gao, Q.; Li, M. IL-38 alleviates concanavalin A-induced liver injury in mice. Int. Immunopharmacol. 2016, 40, 452–457. [Google Scholar] [CrossRef]
- Xu, F.; Lin, S.; Yan, X.; Wang, C.; Tu, H.; Yin, Y.; Cao, J. Interleukin-38 is a Protective Cytokine for Lethal Sepsis. J. Infect. Dis. 2018, 8, 24. [Google Scholar]
- Wang, C.; Chai, Y.; Xie, K.; Lin, S.; Xu, F. Exploring Interleukin-38 As A Potential Therapeutic Tar-Get for Acute Lung Injury Induced by Sepsis. Chest J. 2019, 155, 89A. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Wu, Y.; Dong, N.; Yao, Y.M. Interleukin-38 protects against sepsis by augmenting immunosuppressive activity of CD4+CD25+ regulatory T cells. J. Cell. Molecul. Med. 2020, 2, 2027–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lissoni, P.; Messina, G.; Pelizzoni, F.; Rovelli, F.; Brivio, F.; Monzon, A.; Crivelli, N.; Lissoni, A.; Tassoni, S.; Sassola, A.; et al. The Fascination of Cytokine Immuno-logical Science. J. Infectiology 2020, 3, 18–28. [Google Scholar] [CrossRef]
- Conti, P.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Kritas, S.K.; Frydas, I.; Younes, A.; Di Emidio, P.; Ronconi, G.; Pandolfi, F.; et al. Powerful anti-inflammatory action of luteolin: Potential increase with IL-38. BioFactors 2021, 47, 165–169. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, M.K. A Prospective Study to Assess The Role of Interleukin-38 in Inflammatory Bowel Disease. Ph.D. Thesis, Monash University, Melbourne, Australia. [CrossRef]
- Han, M.-M.; Yuan, X.-R.; Shi, X.; Zhu, X.-Y.; Su, Y.; Xiong, D.-K.; Zhang, X.-M.; Zhou, H.; Wang, J.-N. The Pathological Mechanism and Potential Application of IL-38 in Autoimmune Diseases. Front. Pharmacol. 2021, 12, 2306. [Google Scholar] [CrossRef]
- Wang, M. The Role of IL-37 and IL-38 in Obstetrics Abnormalities. Front. Med. 2021, 8, 1465. [Google Scholar] [CrossRef]
- Wei, Q.; Chen, X.; Wang, C. Contribution of IL-38 in Lung Immunity During Pseudomonas Aeruginosa-Induced Pneumonia. Res. Square 2021, 76, 639. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, H.; Zhao, D.; Hu, H.; Lu, Z. Interleukin-38 Suppresses Cell Migration and Proliferation and Promotes Apoptosis of Colorectal Cancer Cell Through Negatively Regulating Extracellular Signal-Regulated Kinases Signaling. J. Interferon Cytokine Res. 2021, 15, 47. [Google Scholar] [CrossRef]
- Xu, W.D.; Su, L.C.; Liu, X.Y.; Wang, J.M.; Yuan, Z.C.; Qin, Z.; Zhou, X.P.; Huang, A.F. IL-38: A novel cytokine in systemic lupus erythematosus pathogenesis. J. Cell. Mol. Med. 2020, 24, 12379–12389. [Google Scholar] [CrossRef] [PubMed]
- Klück, V.; Liu, R.; Joosten, L.A. The role of interleukin-1 family members in hyperuricemia and gout. Jt. Bone Spine 2021, 88, 105092. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, M.A.; Roshan, S.; Singh, H.; Rawat, S.; Pathak, S.; Mishra, A.; Gupta, G. Role of Various Gene Expressions in Etiopathogenesis of Type 2 Diabetes Mellitus. Adv. Mind Body Med. 2021, 35, 31–39. [Google Scholar]
- Li, J.-M.; Lu, R.; Zhang, Y.; Lin, J.; Hua, X.; Pflugfelder, S.C.; Li, D.-Q. IL-36α/IL-36RA/IL-38 signaling mediates inflam-mation and barrier disruption in human corneal epithelial cells under hyperosmotic stress. Ocul. Surf. 2021, 22, 163–171. [Google Scholar] [CrossRef]
- Salvi, R.; Patankar, P. Emerging pharmacotherapies for COVID-19. Biomed. Pharmacother. 2020, 128, 110267. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune-Mediated Diseases | Immunological Roles of IL-38 | Reference |
|---|---|---|
| Inflammatory Bowel diseases (IBD) | Protective effect in IBD, through production of pro-inflammatory cytokines from macrophages, and a promising immunotherapeutic target in IBD. | [91] |
| Acne Vulgaris | Exacerbation of skin inflammation. | [92] |
| Behcet’s Disease (BD) | Exacerbation of eye involvement, and a protective anti-inflammatory role in BS. | [93,94] |
| Intervertebral Disc Degeneration (IVDD) | Therapeutic roles through alleviation of the inflammatory responses and the degeneration of nucleus pulpous cells via inhibition of the NF-κB signaling pathway. | [95] |
| Alzheimer | Novel biochemical marker with anti-inflammatory activities. | [96] |
| Ischemic stroke | Novel early predictor factor for ischemic stroke prognosis. | [97] |
| Autism spectrum disorder | Therapeutic role through inhibition of activation of human microglia. | [98] |
| Thyroid-associated ophthalmopathy (TAO) | Protective role in TAO, a promising marker of TAO disease activity, and a potential target for TAO therapy. | [99] |
| Multiple sclerosis (MS) | Development of through attenuated inflammatory conditions in early stages of MS. | [100] |
| Experimental autoimmune encephalomyelitis (EAE) | Promotion of inflammation in the central nervous system (CNS). | [101] |
| Candidiasis | Dose-response reduction in Candida-induced T helper 17 responses. | [102] |
| Arthritis | Significant reduction in clinical inflammation and attenuated severity in mouse models of arthritis. | [103] |
| Systemic sclerosis | Role in the pathogenesis of systemic sclerosis. | [104] |
| Atopic dermatitis (eczema) | Prognostication of atopic severity and its inflammatory state in atopic sufferers. | [105] |
| Osteoporosis | Inhibited proliferation of BMSCs and inhibited apoptosis of osteoblasts by regulating the PI3K/Akt/GSK3β/NFATc1 signaling pathway. | [106] |
| Asthma | Development of a regulatory cytokine-based treatment for allergic asthma. | [107] |
| Chronic inflammatory demyelinating polyneuropathy (CIDP) | Making a compensatory mechanism to reduce inflammatory processes in these patients. | [108] |
| Atopic dermatitis, allergic asthma, and allergic rhinitis | Therapeutic potential in the regulation of allergy asthma, and allergic rhinitis. | [109] |
| Brucellosis | Progression from acute into the chronic forms of brucellosis. | [110] |
| inflammatory diseases (psoriasis, rheumatoid arthritis, gout, systemic lupus erythematosus, and Crohn’s disease) | Involved in the pathogenesis of inflammatory diseases. | [111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esmaeilzadeh, A.; Bahmaie, N.; Nouri, E.; Hajkazemi, M.J.; Zareh Rafie, M. Immunobiological Properties and Clinical Applications of Interleukin-38 for Immune-Mediated Disorders: A Systematic Review Study. Int. J. Mol. Sci. 2021, 22, 12552. https://doi.org/10.3390/ijms222212552
Esmaeilzadeh A, Bahmaie N, Nouri E, Hajkazemi MJ, Zareh Rafie M. Immunobiological Properties and Clinical Applications of Interleukin-38 for Immune-Mediated Disorders: A Systematic Review Study. International Journal of Molecular Sciences. 2021; 22(22):12552. https://doi.org/10.3390/ijms222212552
Chicago/Turabian StyleEsmaeilzadeh, Abdolreza, Nazila Bahmaie, Elham Nouri, Mohammad Javad Hajkazemi, and Maryam Zareh Rafie. 2021. "Immunobiological Properties and Clinical Applications of Interleukin-38 for Immune-Mediated Disorders: A Systematic Review Study" International Journal of Molecular Sciences 22, no. 22: 12552. https://doi.org/10.3390/ijms222212552