
Genome-Wide Analysis of the Apple CBL Family Reveals That Mdcbl10.1 Functions Positively in Modulating Apple Salt Tolerance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Retrieval and Identification of Apple CBL Family Proteins
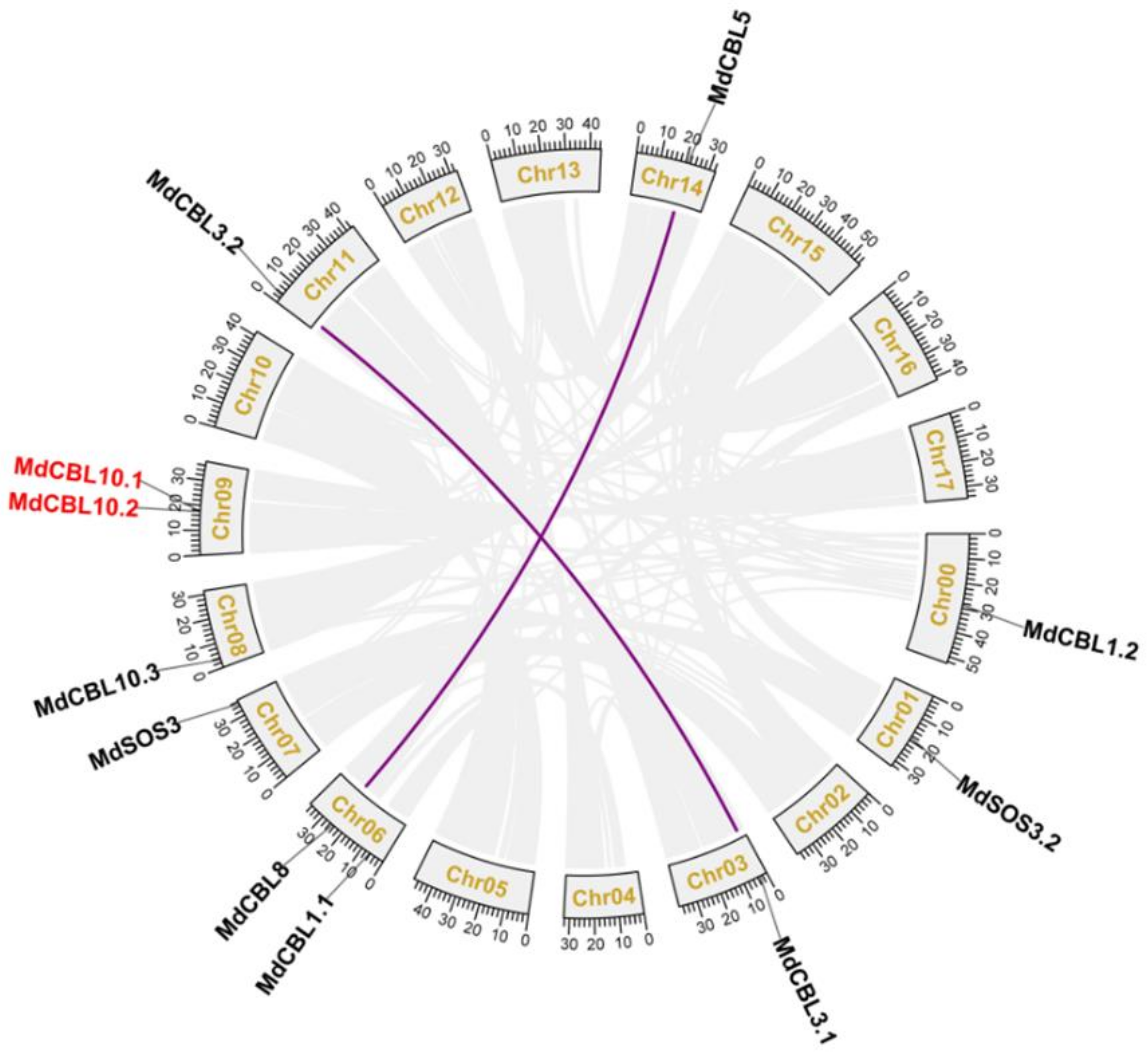
2.2. Collinearity Analysis and Characterization of Apple CBL Family Genes
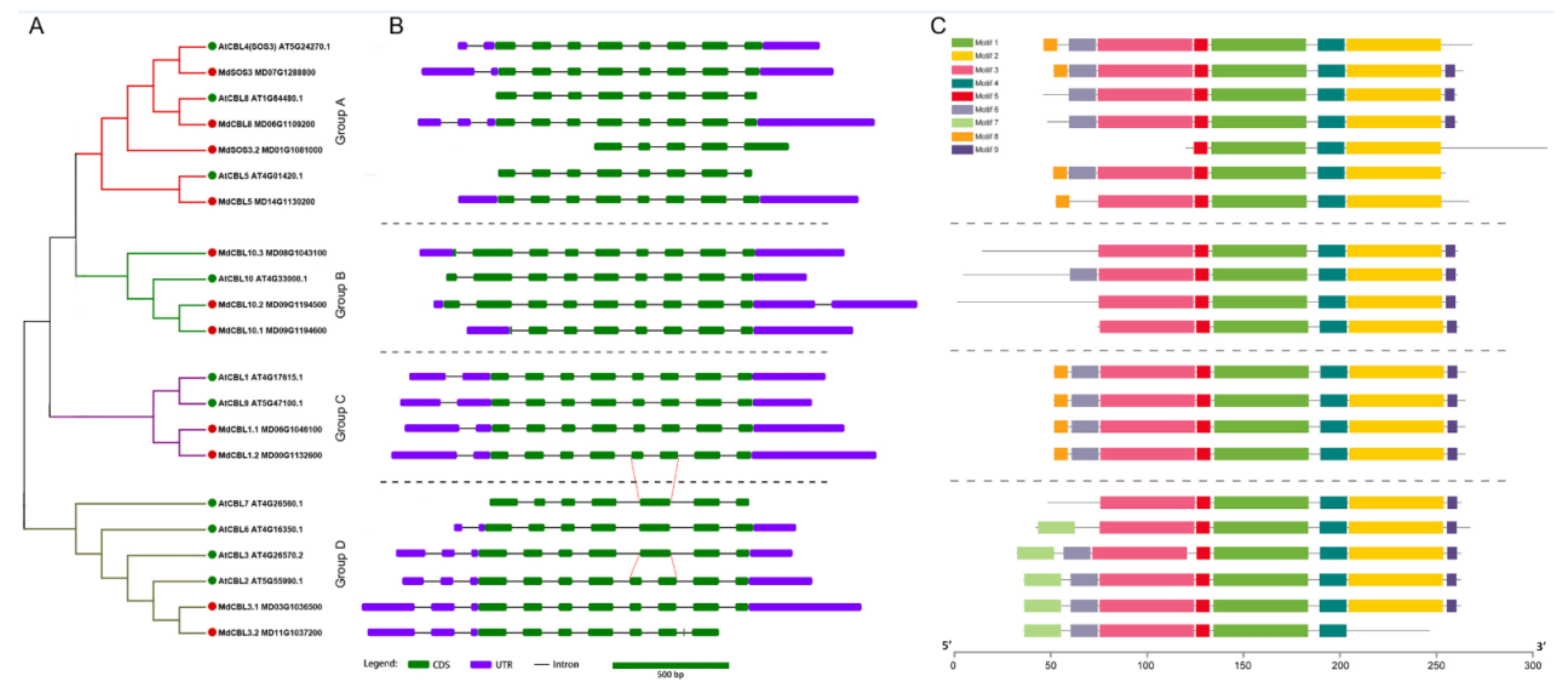
2.3. Phylogenetic Relationships, Gene Structure, and Conserved Motif Analysis
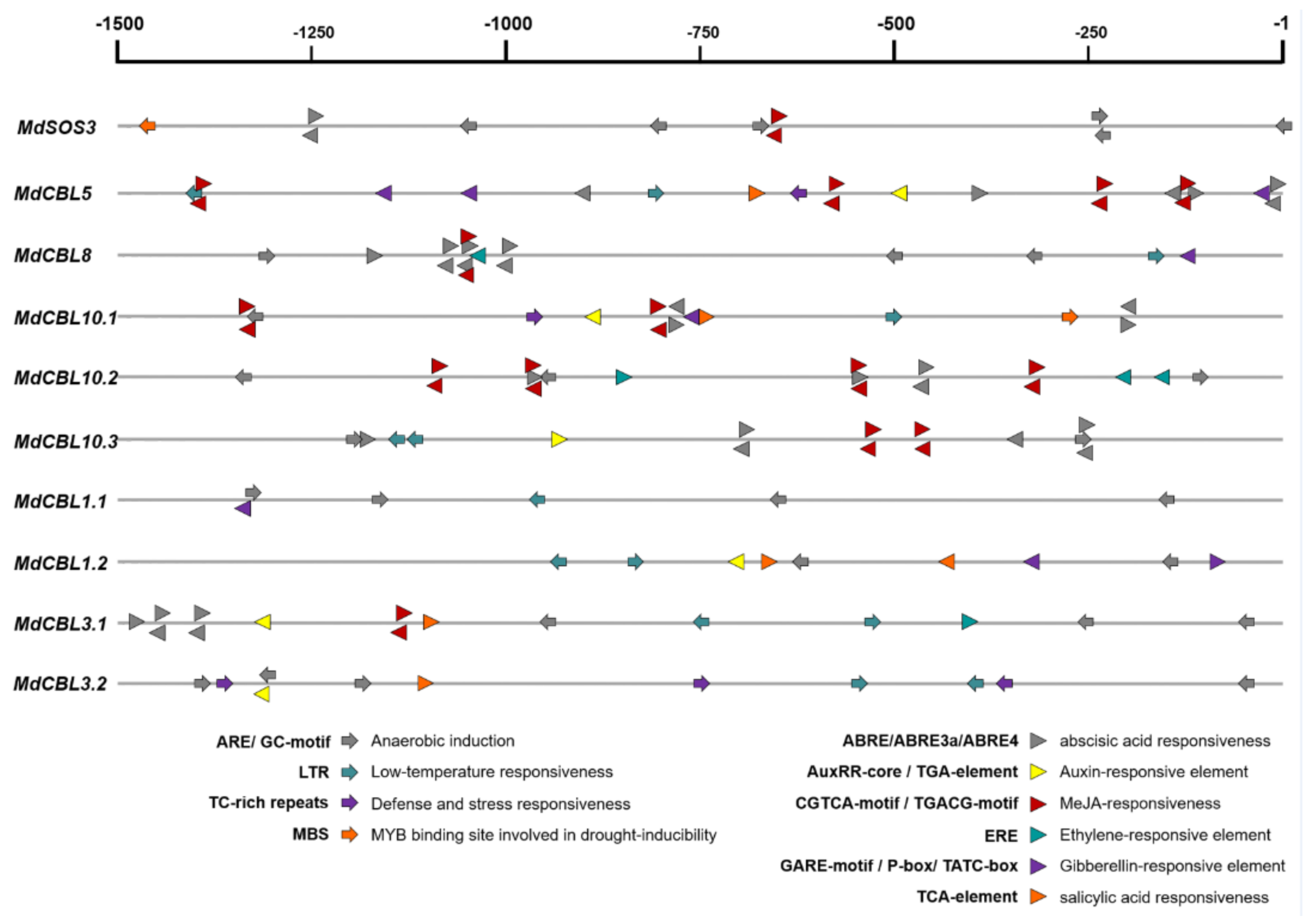
2.4. Cis-Acting Elements in the Promoters of MdCBL Genes
2.5. Vector Construction, Genetic Transformation, and Stress Treatment
2.6. Measurement of Stress-Related Physiological Parameters
2.7. Statistical Analysis
3. Results
3.1. Identification, Characterization, and Gene Duplication of Apple CBL Family Genes
3.2. Phylogenetic Analysis, Gene Structure Display, Prediction of Conserved Motifs, and Cloning of Apple CBL Family Genes
3.3. Promoter Analysis of MdCBL Genes
3.4. Functional Identification of Mdcbls in Regulating Salt Tolerance in Yeast
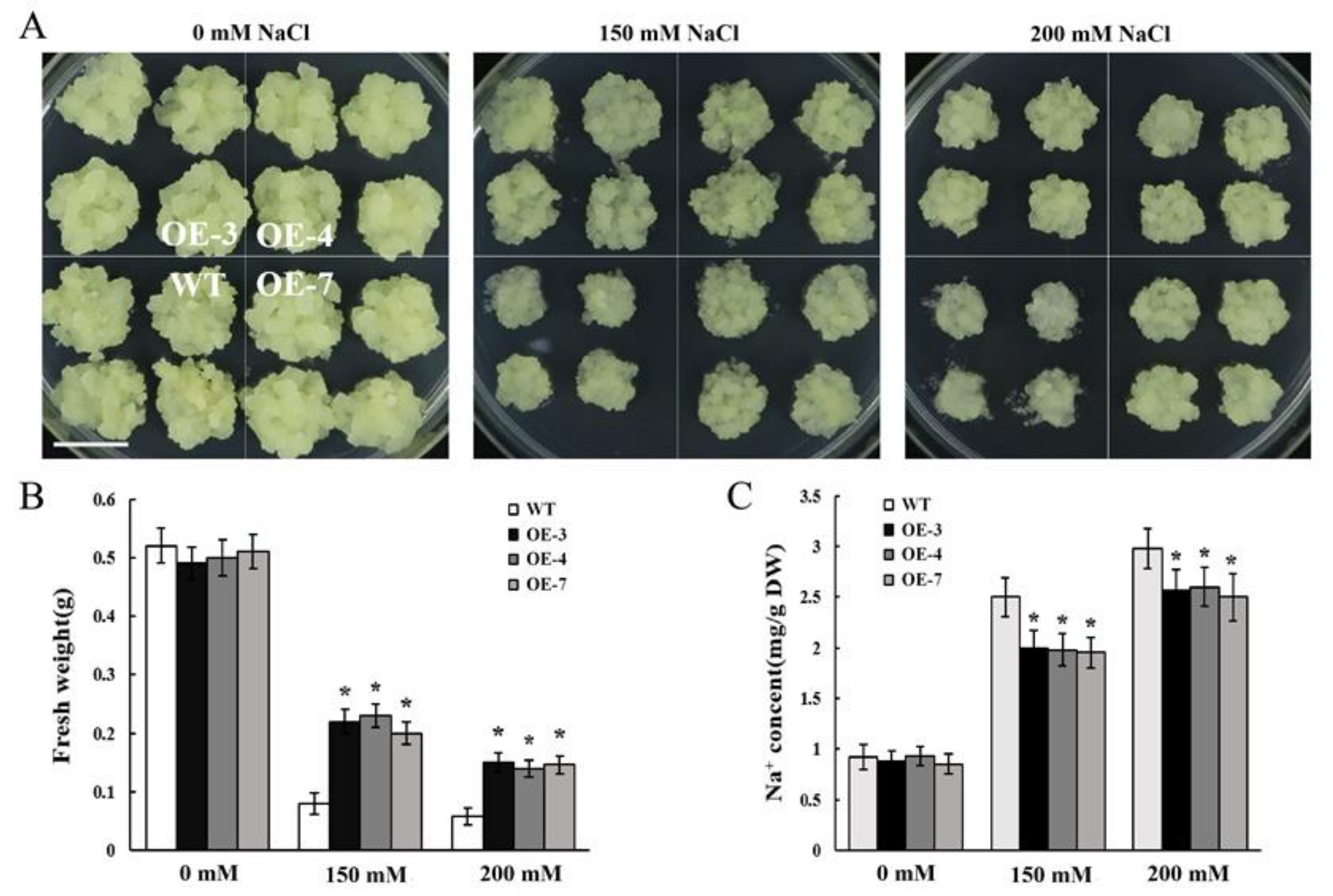
3.5. Overexpression of MdCBL10.1 Improved Salt Tolerance of Cisgenic Apple Calli
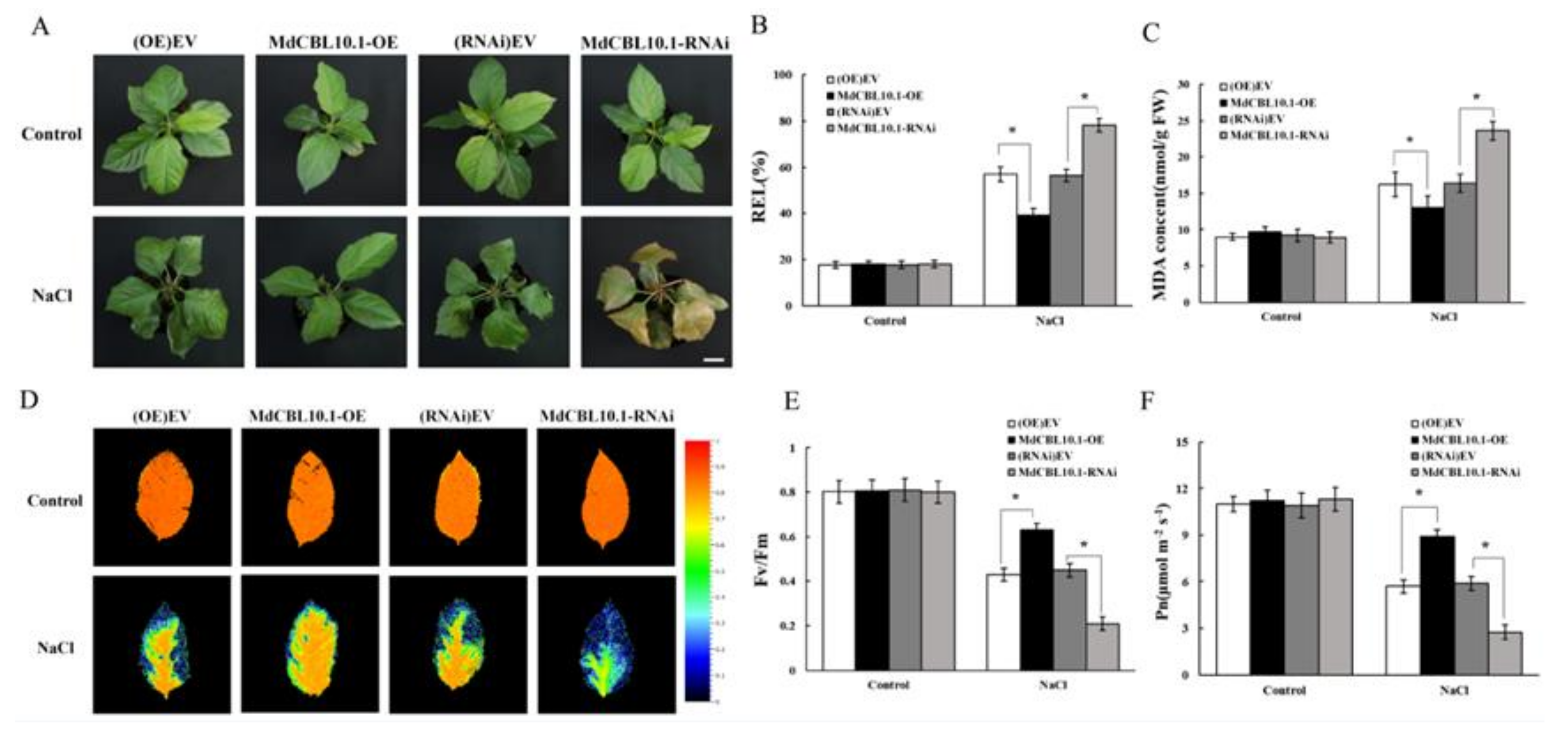
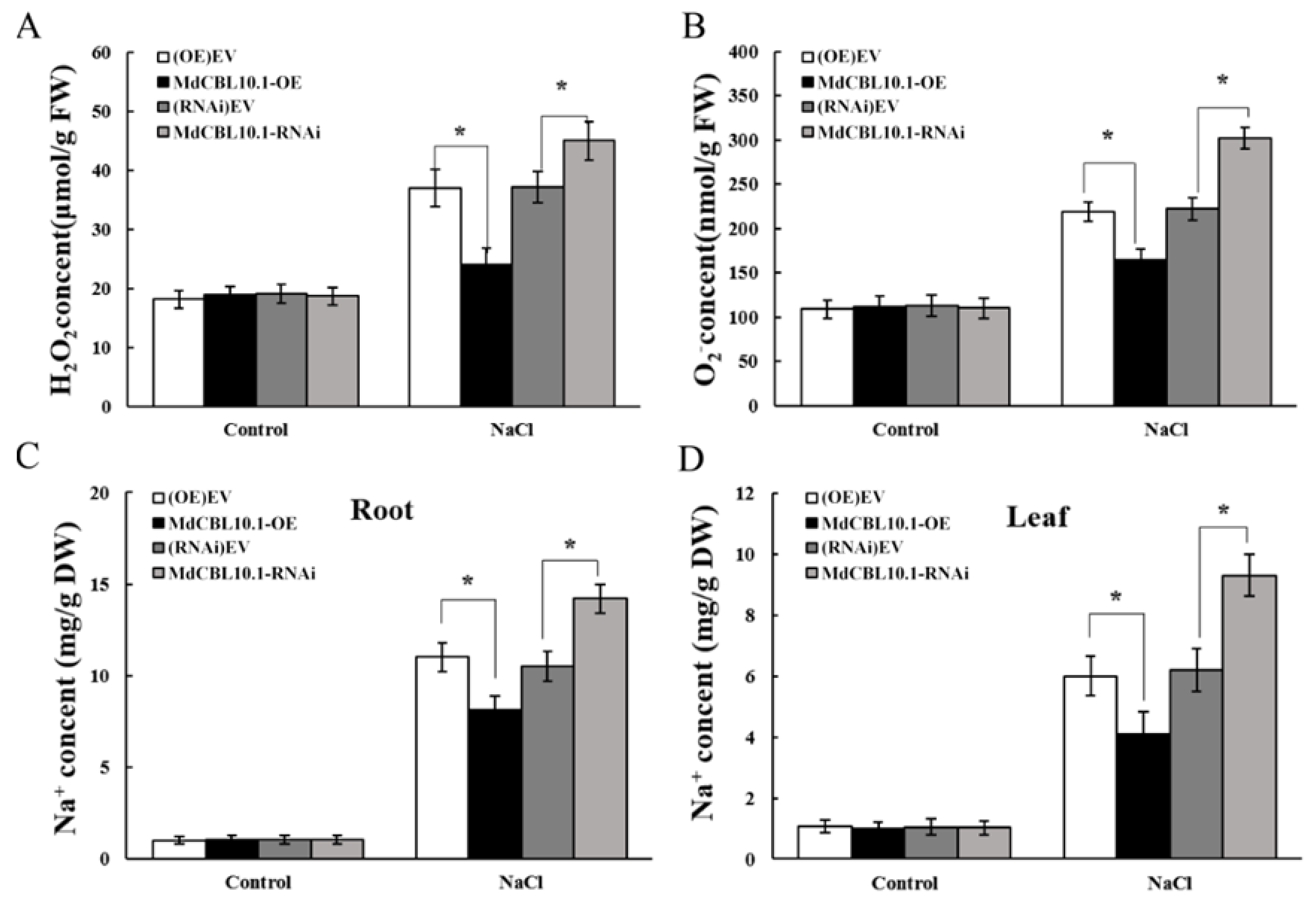
3.6. Overexpression of MdCBL10.1 in Roots Enhanced the Salt Tolerance of Apple Plants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, T.T.; Zhang, G.F.; Sun, L.R.; Wang, J.; Hao, F.S. Genome-wide identification of CBL family and expression analysis of CBLs in response to potassium deficiency in cotton. PeerJ 2017, 5, e3653. [Google Scholar] [CrossRef] [Green Version]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B-like proteins: Calcium sensors for specific signal response coupling in plants. Plant Cell 2002, 14, S389–S400. [Google Scholar] [CrossRef] [Green Version]
- Cheong, Y.H.; Pandey, G.K.; Grant, J.J.; Batistic, O.; Li, L.; Kim, B.G.; Lee, S.C.; Kudla, J.; Luan, S. Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. Cell Mol. Biol. 2007, 52, 223–239. [Google Scholar] [CrossRef]
- Mazars, C.; Bourque, S.; Mithofer, A.; Pugin, A.; Ranjeva, R. Calcium homeostasis in plant cell nuclei. New Phytol. 2009, 181, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Bethke, P.C.; Jones, R.L. Calcium homeostasis in plants. J. Cell Sci. 1993, 106, 453–462. [Google Scholar] [CrossRef]
- Kumar, M.; Sharma, K.; Yadav, A.K.; Kanchan, K.; Baghel, M.; Kateriya, S.; Pandey, G.K. Genome-wide identification and biochemical characterization of calcineurin B-like calcium sensor proteins in Chlamydomonas reinhardtii. Biochem. J. 2020, 477, 1879–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, S.K.; Pandey, A.; Pandey, G.K. The CBL-CIPK signaling module in plants: A mechanistic perspective. Physiol. Plant. 2015, 155, 89–108. [Google Scholar] [CrossRef]
- Xi, Y.; Liu, J.Y.; Dong, C.; Cheng, Z.M. The CBL and CIPK gene family in grapevine (Vitis vinifera): Genome-wide analysis and expression profiles in response to various abiotic stresses. Front. Plant Sci. 2017, 8, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.N.; Zhang, B.; Deng, J.W.; Chen, L.; Ullah, A.; Yang, X.Y. Genome-wide analysis of CBL and CIPK family genes in cotton: Conserved structures with divergent interactions and expression. Physiol. Mol. Biol. Plant 2021, 27, 359–368. [Google Scholar] [CrossRef]
- Ma, X.; Gai, W.X.; Qiao, Y.M.; Ali, M.; Wei, A.M.; Luo, D.X.; Li, Q.H.; Gong, Z.H. Identification of CBL and CIPK gene families and functional characterization of CaCIPK1 under Phytophthora capsici in pepper (Capsicum annuum L.). BMC Genom. 2019, 20, 775. [Google Scholar] [CrossRef]
- Aslam, M.; Fakher, B.; Jakada, B.H.; Zhao, L.H.; Cao, S.J.; Cheng, Y.; Qin, Y. Genome-wide identification and expression profiling of CBL-CIPK gene family in pineapple (Ananas comosus) and the role of AcCBL1 in abiotic and biotic stress response. Biomolecules 2019, 9, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Yan, Y.; Tie, W.W.; Ding, Z.H.; Wu, C.L.; Ding, X.P.; Wang, W.Q.; Xia, Z.Q.; Guo, J.C.; Peng, M. Genome-wide analyses of calcium sensors reveal their involvement in drought stress response and storage roots deterioration after harvest in Cassava. Genes 2018, 9, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.F.; Xu, C.; Cao, H.H.; Shi, Y.; Chen, J.; Chai, Y.; Li, Z.G. Tomato calmodulin-like protein SlCML37 is a calcium (Ca2+) sensor that interacts with proteasome maturation factor SlUMP1 and plays a role in tomato fruit chilling stress tolerance. J. Plant Physiol. 2021, 258, 153373. [Google Scholar] [CrossRef]
- Jung, H.; Chung, P.J.; Park, S.H.; Redillas, M.C.F.R.; Kim, Y.S.; Suh, J.W.; Kim, J.K. Overexpression of OsERF48 causes regulation of OsCML16, a calmodulin-like protein gene that enhances root growth and drought tolerance. Plant Biotechnol. J. 2017, 15, 1295–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Verde, V.; Trande, M.; D’Onofrio, M.; Dominici, P.; Astegno, A. Binding of calcium and target peptide to calmodulin-like protein CML19, the centrin 2 of Arabidopsis thaliana. Int. J. Biol. Macromol. 2018, 108, 1289–1299. [Google Scholar] [CrossRef]
- Trande, M.; Pedretti, M.; Bonza, M.C.; Di Matteo, A.; D’Onofrio, M.; Dominici, P.; Astegno, A. Cation and peptide binding properties of CML7, a calmodulin-like protein from Arabidopsis thaliana. J. Inorg. Biochem. 2019, 199, 110796. [Google Scholar] [CrossRef]
- Zhao, P.C.; Liu, Y.J.; Kong, W.Y.; Ji, J.Y.; Cai, T.Y.; Guo, Z.F. Genome-wide identification and characterization of calcium-dependent protein kinase (CDPK) and CDPK-related kinase (CRK) gene families in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 1044. [Google Scholar] [CrossRef]
- Bi, Z.Z.; Wang, Y.H.; Li, P.C.; Sun, C.; Qin, T.Y.; Bai, J.P. Evolution and expression analysis of CDPK genes under drought stress in two varieties of potato. Biotechnol. Lett. 2021, 43, 511–521. [Google Scholar] [CrossRef]
- Crizel, R.L.; Perin, E.C.; Vighi, I.L.; Woloski, R.; Seixas, A.; Pinto, L.D.; Rombaldi, C.V.; Galli, V. Genome-wide identification, and characterization of the CDPK gene family reveal their involvement in abiotic stress response in Fragaria × ananassa. Sci. Rep. 2020, 10, 11040. [Google Scholar] [CrossRef]
- Chu, L.C.; Offenborn, J.N.; Steinhorst, L.; Wu, X.N.; Xi, L.; Li, Z.; Jacquot, A.; Lejay, L.; Kudla, J.; Schulze, W.X. Plasma membrane calcineurin B-like calcium-ion sensor proteins function in regulating primary root growth and nitrate uptake by affecting global phosphorylation patterns and microdomain protein distribution. New Phytol. 2021, 229, 2223–2237. [Google Scholar] [CrossRef]
- Lu, L.; Chen, X.Y.; Zhu, L.M.; Li, M.J.; Zhang, J.B.; Yang, X.Y.; Wang, P.K.; Lu, Y.; Cheng, T.L.; Shi, J.S.; et al. NtCIPK9: A calcineurin B-like protein-interacting protein kinase from the Halophyte Nitraria tangutorum, enhances arabidopsis salt tolerance. Front. Plant Sci. 2020, 11, 1112. [Google Scholar] [CrossRef]
- Albrecht, V.; Weinl, S.; Blazevic, D.; D’Angelo, C.; Batistic, O.; Kolukisaoglu, U.; Bock, R.; Schulz, B.; Harter, K.; Kudla, J. The calcium sensor CBL1 integrates plant responses to abiotic stresses. Plant J. Cell Mol. Biol. 2003, 36, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wang, C.; Li, L.; Fu, D.; Zhang, Y.; Yang, P.; Zhang, T.; Wang, C. A novel role of the calcium sensor CBL1 in response to phosphate deficiency in Arabidopsis thaliana. J. Plant Physiol. 2020, 253, 153266. [Google Scholar] [CrossRef]
- Yan, Y.; He, M.; Guo, J.; Zeng, H.; Wei, Y.; Liu, G.; Hu, W.; Shi, H. The CBL1/9-CIPK23-AKT1 complex is essential for low potassium response in cassava. Plant Physiol. Biochem. PPB 2021, 167, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, K.; Pascaud, F.; Eckert, C.; Gajdanowicz, P.; Hashimoto, K.; Corratge-Faillie, C.; Offenborn, J.N.; Lacombe, B.; Dreyer, I.; Thibaud, J.B.; et al. Calcium-dependent modulation and plasma membrane targeting of the AKT2 potassium channel by the CBL4/CIPK6 calcium sensor/protein kinase complex. Cell Res. 2011, 21, 1116–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.G.; Waadt, R.; Cheong, Y.H.; Pandey, G.K.; Dominguez-Solis, J.R.; Schultke, S.; Lee, S.C.; Kudla, J.; Luan, S. The calcium sensor CBL10 mediates salt tolerance by regulating ion homeostasis in Arabidopsis. Plant J. Cell Mol. Biol. 2007, 52, 473–484. [Google Scholar] [CrossRef]
- Quan, R.; Lin, H.; Mendoza, I.; Zhang, Y.; Cao, W.; Yang, Y.; Shang, M.; Chen, S.; Pardo, J.M.; Guo, Y. SCABP8/CBL10, a putative calcium sensor, interacts with the protein kinase SOS2 to protect Arabidopsis shoots from salt stress. Plant Cell 2007, 19, 1415–1431. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Y.X.; Li, H.; Teng, R.M.; Wang, Y.; Zhuang, J. Genome-wide identification and expression analysis of calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase family genes in tea plant. DNA Cell Biol. 2019, 38, 824–839. [Google Scholar] [CrossRef] [PubMed]
- Kolukisaoglu, U.; Weinl, S.; Blazevic, D.; Batistic, O.; Kudla, J. Calcium sensors and their interacting protein kinases: Genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Yue, D.; Wei, W.; Hu, Y.; Feng, J.; Zou, Z. Characterization and functional analysis of calmodulin and calmodulin-like genes in Fragaria vesca. Front. Plant Sci. 2016, 7, 1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Hou, X.; Xia, Z.; Yan, Y.; Wei, Y.; Wang, L.; Zou, M.; Lu, C.; Wang, W.; Peng, M. Genome-wide survey and expression analysis of the calcium-dependent protein kinase gene family in cassava. Mol. Genet. Genom. MGG 2016, 291, 241–253. [Google Scholar] [CrossRef]
- Li, J.; Jiang, M.M.; Ren, L.; Liu, Y.; Chen, H.Y. Identification and characterization of CBL and CIPK gene families in eggplant (Solanum melongena L.). Mol. Genet. Genom. MGG 2016, 291, 1769–1781. [Google Scholar] [CrossRef]
- Mao, K.; Dong, Q.; Li, C.; Liu, C.; Ma, F. Genome wide identification and characterization of apple bHLH Transcription factors and expression analysis in response to drought and salt stress. Front. Plant Sci. 2017, 8, 480. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Yang, J.; Wang, M.; Liu, H.; Guo, X.; Zhao, S.; Dong, Q.; Ma, F. Genome-wide analysis of the apple CaCA superfamily reveals that MdCAX proteins are involved in the abiotic stress response as calcium transporters. BMC Plant Biol. 2021, 21, 81. [Google Scholar] [CrossRef]
- Yang, J.; Guo, X.; Li, W.; Chen, P.; Cheng, Y.; Ma, F.; Mao, K. MdCCX2 of apple functions positively in modulation of salt tolerance. Environ. Exp. Bot. 2021, 192, 104663. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.; Guo, X.; Chen, P.; Cheng, Y.; Mao, K.; Ma, F. Cation/Ca2+ Exchanger 1 (MdCCX1), a plasma membrane-localized Na+ transporter, enhances plant salt tolerance by inhibiting excessive accumulation of Na+ and reactive oxygen species. Front. Plant Sci. 2021, 12, 46189. [Google Scholar] [CrossRef]
- Yin, X.; Xia, Y.; Xie, Q.; Cao, Y.; Wang, Z.; Hao, G.; Song, J.; Zhou, Y.; Jiang, X. The protein kinase complex CBL10-CIPK8-SOS1 functions in Arabidopsis to regulate salt tolerance. J. Exp. Bot. 2020, 71, 1801–1814. [Google Scholar] [CrossRef]
- Tang, R.J.; Wang, C.; Li, K.L.; Luan, S. The CBL-CIPK Calcium signaling network: Unified paradigm from 20 years of discoveries. Trends Plant Sci. 2020, 25, 604–617. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Samiotaki, M.; Tsiolas, G.; Sarrou, E.; Stamatakis, G.; Ganopoulos, I.; Martens, S.; Argiriou, A.; et al. Novel insights into the calcium action in cherry fruit development revealed by high-throughput mapping. Plant Mol. Biol. 2020, 104, 597–614. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Karamanoli, K.; Lazaridou, A.; Matsi, T.; Molassiotis, A. Metabolomic and physico-chemical approach unravel dynamic regulation of calcium in sweet cherry fruit physiology. Plant Physiol. Biochim. 2017, 116, 68–79. [Google Scholar] [CrossRef]
- Yu, J.; Zhu, M.T.; Wang, M.J.; Xu, Y.S.; Chen, W.T.; Yang, G.S. Transcriptome analysis of calcium-induced accumulation of anthocyanins in grape skin. Sci. Hortic. 2020, 260, 108871. [Google Scholar] [CrossRef]
- Martins, V.; Billet, K.; Garcia, A.; Lanoue, A.; Geros, H. Exogenous calcium deflects grape berry metabolism towards the production of more stilbenoids and less anthocyanins. Food Chem. 2020, 313, 126123. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Sarrou, E.; Stavridou, E.; Ganopoulos, I.; Karamanoli, K.; Madesis, P.; Martens, S.; Molassiotis, A. An integrated metabolomic and gene expression analysis identifies heat and calcium metabolic networks underlying postharvest sweet cherry fruit senescence. Planta 2019, 250, 2009–2022. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Mohanta, N.; Mohanta, Y.K.; Parida, P.; Bae, H.H. Genome-wide identification of Calcineurin B-Like (CBL) gene family of plants reveals novel conserved motifs and evolutionary aspects in calcium signaling events. BMC Plant Biol. 2015, 15, 189. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.H.; Dong, B.Y.; Yang, Q.; Niu, L.L.; Li, H.H.; Cao, H.Y.; Amin, R.; Meng, D.; Fu, Y.J. Screening of CBL genes in pigeon pea with focus on the functional analysis of CBL4 in abiotic stress tolerance and flavonoid biosynthesis. Environ. Exp. Bot. 2020, 177, 104102. [Google Scholar] [CrossRef]
- Xiong, Y.; Li, R.M.; Lin, X.J.; Zhou, Y.J.; Tang, F.L.; Yao, Y.; Liu, J.; Wang, L.X.; Yin, X.M.; Liu, Y.X.; et al. Genome-wide analysis and expression tendency of banana (Musa acuminata L.) calcineurin B-like (MaCBL) genes under potassium stress. Horticulturae 2021, 7, 70. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, H.; Luo, J.; Wang, H.; Dong, Q.; Wang, Y.; Yang, K.; Mao, K.; Ma, F. Genome-wide analysis of the light-harvesting chlorophyll a/b-binding gene family in apple (Malus domestica) and functional characterization of MdLhcb4.3, which confers tolerance to drought and osmotic stress. Plant Physiol. Biochem. PPB 2020, 154, 517–529. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhao, C.L.; Zhao, M.F.; Li, Y.Z.; Wen, G.S. Phylogeny and evolution of calcineurin B-like (CBL) gene family in grass and functional analyses of rice CBLs. J. Plant Biol. 2020, 63, 117–130. [Google Scholar] [CrossRef]
- Hu, D.G.; Ma, Q.J.; Sun, C.H.; Sun, M.H.; You, C.X.; Hao, Y.J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plant. 2016, 156, 201–214. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Gene Name | Gene Locus (GDDH13) | Genomic Location (GDDH13) | Deduced Polypeptide | Best Hits | |||
|---|---|---|---|---|---|---|---|---|
| Length (aa) | Mass Weight (kDa) | pI | Charge at PH 7.0 | |||||
| A | MdSOS3 | MD07G1288800 | Chr07: 34980079–34983632 | 212 | 24.39 | 4.52 | −16.44 | AT5G24270 |
| MdSOS3.2 | MD01G1081000 | Chr01: 18815406–18816403 | 187 | 21.38 | 4.53 | −9.88 | AT5G24270 | |
| MdCBL5 | MD14G1130200 | Chr14: 20784655–20786608 | 214 | 24.53 | 4.42 | −15.87 | AT4G26570 | |
| MdCBL8 | MD06G1109200 | Chr06: 24877949–24881781 | 212 | 24.42 | 4.60 | −13.48 | AT1G64480 | |
| B | MdCBL10.1 | MD09G1194600 | Chr09: 17493583–17495782 | 187 (266) | 21.57 (30.51) | 4.48 (4.88) | −14.55 (−11.27) | AT4G33000 |
| MdCBL10.2 | MD09G1194500 | Chr09: 17485706–17488584 | 259 (245) | 29.53 (27.88) | 4.54 (4.63) | −17.30 (−14.31) | AT4G33000 | |
| MdCBL10.3 | MD08G1043100 | Chr08: 3254626–3256803 | 246 | 28.28 | 4.65 | −17.03 | AT4G33000 | |
| C | MdCBL1.1* | MD06G1046100 | Chr06: 6240265–6244670 | 213 | 24.52 | 4.63 | −10.78 | AT4G17615 |
| MdCBL1.2 | MD00G1132600 | Chr00: 28662543–28667386 | 213 | 24.52 | 4.63 | −10.78 | AT4G17615 | |
| D | MdCBL3.1 | MD03G1036500 | Chr03: 2901761–2905532 | 226 | 26.06 | 4.69 | −14.17 | AT4G26570 |
| MdCBL3.2 | MD11G1037200 | Chr11: 3199328–3202169 | 210 (226) | 24.20 (25.96) | 4.89 (4.75) | −10.34 (−13.17) | AT4G26570 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.; Yang, J.; Mei, Q.; Liu, H.; Cheng, Y.; Ma, F.; Mao, K. Genome-Wide Analysis of the Apple CBL Family Reveals That Mdcbl10.1 Functions Positively in Modulating Apple Salt Tolerance. Int. J. Mol. Sci. 2021, 22, 12430. https://doi.org/10.3390/ijms222212430
Chen P, Yang J, Mei Q, Liu H, Cheng Y, Ma F, Mao K. Genome-Wide Analysis of the Apple CBL Family Reveals That Mdcbl10.1 Functions Positively in Modulating Apple Salt Tolerance. International Journal of Molecular Sciences. 2021; 22(22):12430. https://doi.org/10.3390/ijms222212430
Chicago/Turabian StyleChen, Peihong, Jie Yang, Quanlin Mei, Huayu Liu, Yunpeng Cheng, Fengwang Ma, and Ke Mao. 2021. "Genome-Wide Analysis of the Apple CBL Family Reveals That Mdcbl10.1 Functions Positively in Modulating Apple Salt Tolerance" International Journal of Molecular Sciences 22, no. 22: 12430. https://doi.org/10.3390/ijms222212430