
Encapsulin Based Self-Assembling Iron-Containing Protein Nanoparticles for Stem Cells MRI Visualization

 , and
, and 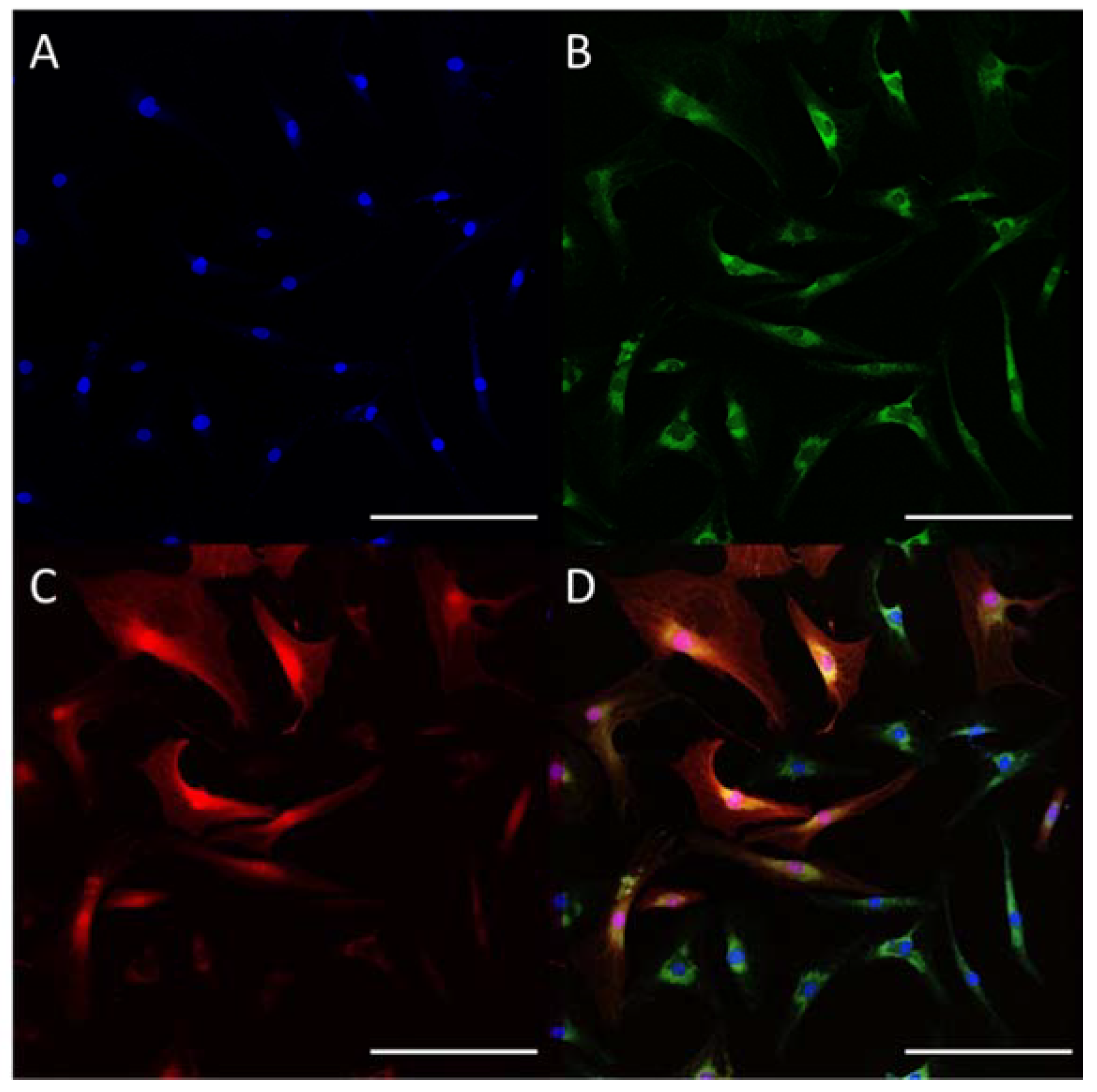{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Encapsulin Expression
2.2. Iron Biomineralization
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Flow Cytometry
4.3. Differentiation of the Cells
4.4. Construction of Lentiviruses and Lentivirus Transduction of Cells
4.5. Laser Scanning Confocal Microscopy
4.6. PCR with Reverse Transcription
4.7. Western Blot Analysis
4.8. Cytotoxicity Study of FAS In Vitro
4.9. Prussian Blue Staining
4.10. TEM
4.11. MRI
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simmons, J.K.; Hildreth, B.; Supsavhad, W.; Elshafae, S.M.; Hassan, B.B.; Dirksen, W.P.; Toribio, R.E.; Rosol, T.J. Animal Models of Bone Metastasis. Veter. Pathol. 2015, 52, 827–841. [Google Scholar] [CrossRef]
- Zhang, S.; Xie, R.; Zhao, T.; Yang, X.; Han, L.; Ye, F.; Lei, T.; Wan, F. Neural stem cells preferentially migrate to glioma stem cells and reduce their stemness phenotypes. Int. J. Oncol. 2014, 45, 1989–1996. [Google Scholar] [CrossRef] [Green Version]
- Michalet, X.; Pinaud, F.F.; Bentolila, L.A.; Tsay, J.M.; Doose, S.; Li, J.J.; Sundaresan, G.; Wu, A.M.; Gambhir, S.S.; Weiss, S. Quantum Dots for Live Cells, in Vivo Imaging, and Diagnostics. Science 2005, 307, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Xie, X.; Patel, M.R.; Yang, Y.-H.; Li, Z.; Cao, F.; Gheysens, O.; Zhang, Y.; Gambhir, S.S.; Rao, J.H.; et al. Quantum dot imaging for embryonic stem cells. BMC Biotechnol. 2007, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Shah, B.S.; Mao, J.J. Labeling of Mesenchymal Stem Cells with Bioconjugated Quantum Dots. Methods Mol. Biol. 2011, 680, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Tamura, R.; Jiang, F.; Xie, J.; Kamiyama, D. Multiplexed labeling of cellular proteins with split fluorescent protein tags. Commun. Biol. 2021, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.C.; Nabel, E.M.; Murdock, M.H.; Lao-Peregrin, C.; Tsoulfas, P.; Blackmore, M.G.; Lee, F.S.; Liston, C.; Morishita, H.; Petsko, G.A. mGreenLantern: A bright monomeric fluorescent protein with rapid expression and cell filling properties for neuronal imaging. Proc. Natl. Acad. Sci. USA 2020, 117, 30710–30721. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.; Xu, T.; Kirkpatrick, A.; Ripp, S.; Sayler, G.; Close, D. Real-time tracking of stem cell viability, proliferation, and differentiation with autonomous bioluminescence imaging. BMC Biol. 2020, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Park, B.N.; Shim, W.; Lee, G.; Bang, O.Y.; An, Y.S.; Yoon, J.K.; Ahn, Y.H. Early Distribution of Intravenously Injected Mesenchymal Stem Cells in Rats with Acute Brain Trauma Evaluated by 99mTc-HMPAO Labeling. Nucl. Med. Biol. 2011, 38, 1175–1182. [Google Scholar] [CrossRef]
- Gholamrezanezhad, A.; Mirpour, S.; Bagheri, M.; Mohamadnejad, M.; Alimoghaddam, K.; Abdolahzadeh, L.; Saghari, M.; Malekzadeh, R. In vivo tracking of 111In-oxine labeled mesenchymal stem cells following infusion in patients with advanced cirrhosis. Nucl. Med. Biol. 2011, 38, 961–967. [Google Scholar] [CrossRef]
- Wolfs, E.; Struys, T.; Notelaers, T.; Roberts, S.J.; Sohni, A.; Bormans, G.; Van Laere, K.; Luyten, F.P.; Gheysens, O.; Lambrichts, I.; et al. 18F-FDG Labeling of Mesenchymal Stem Cells and Multipotent Adult Progenitor Cells for PET Imaging: Effects on Ultrastructure and Differentiation Capacity. J. Nucl. Med. 2013, 54, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Tarantal, A.F.; Lee, C.C.I.; Batchelder, C.A.; Christensen, J.E.; Prater, D.; Cherry, S.R. Radiolabeling and in vivo imaging of transplanted renal lineages differentiated from human embryonic stem cells in fetal rhesus monkeys. Mol. Imaging Biol. 2011, 14, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Ponomarev, V.; Doubrovin, M.; Shavrin, A.; Serganova, I.; Beresten, T.; Ageyeva, L.; Cai, C.; Balatoni, J.; Alauddin, M.; Gelovani, J. A Human-Derived Reporter Gene for Noninvasive Imaging in Humans: Mitochondrial Thymidine Kinase Type 2. J. Nucl. Med. 2007, 48, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-H.; Chen, F.-D.; Wang, H.-E.; Chang, C.-W.; Lee, Y.-J.; Gelovani, J.G.; Liu, R.-S. Generation of Destabilized Herpes Simplex Virus Type 1 Thymidine Kinase as Transcription Reporter for PET Reporter Systems in Molecular–Genetic Imaging. J. Nucl. Med. 2008, 49, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Yaghoubi, S.S.; Campbell, D.O.; Radu, C.G.; Czernin, J. Positron Emission Tomography Reporter Genes and Reporter Probes: Gene and Cell Therapy Applications. Theranostics 2012, 2, 374–391. [Google Scholar] [CrossRef] [Green Version]
- Ittrich, H.; Lange, C.; Tögel, F.; Zander, A.R.; Dahnke, H.; Westenfelder, C.; Adam, G.; Nolte-Ernsting, C. In vivo magnetic resonance imaging of iron oxide–labeled, arterially-injected mesenchymal stem cells in kidneys of rats with acute ischemic kidney injury: Detection and monitoring at 3T. J. Magn. Reson. Imaging 2007, 25, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Namestnikova, D.D.; Gubskiy, I.L.; Revkova, V.A.; Sukhinich, K.K.; Melnikov, P.A.; Gabashvili, A.N.; Cherkashova, E.A.; Vishnevskiy, D.A.; Kurilo, V.V.; Burunova, V.V.; et al. Intra-Arterial Stem Cell Transplantation in Experimental Stroke in Rats: Real-Time MR Visualization of Transplanted Cells Starting with Their First Pass Through the Brain With Regard to the Therapeutic Action. Front. Neurosci. 2021, 15, 15. [Google Scholar] [CrossRef]
- Jiang, L.; Li, R.; Tang, H.; Zhong, J.; Sun, H.; Tang, W.; Wang, H.; Zhu, J. MRI Tracking of iPS Cells-Induced Neural Stem Cells in Traumatic Brain Injury Rats. Cell Transplant. 2019, 28, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Vallabani, S.S.A.A.K.N.V.S.; Singh, S.; Karakoti, A. Magnetic Nanoparticles: Current Trends and Future Aspects in Diagnostics and Nanomedicine. Curr. Drug Metab. 2019, 20, 457–472. [Google Scholar] [CrossRef]
- Guldris, N.; Argibay, B.; Gallo, J.; Iglesias-Rey, R.; Carbó-Argibay, E.; Kolen’Ko, Y.V.; Campos, F.; Sobrino, T.; Salonen, L.M.; Bañobre-López, M.; et al. Magnetite Nanoparticles for Stem Cell Labeling with High Efficiency and Long-Term in Vivo Tracking. Bioconjug. Chem. 2016, 28, 362–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Zafar, H.; Zia, M.; ul Haq, I.; Phull, A.R.; Ali, J.S.; Hussain, A. Synthesis, characterization, applications, and challenges of iron oxide nanoparticles. Nanotechnol. Sci. Appl. 2016, 9, 49–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.; Chandrasekaran, R. Biogenic nanoparticles: A comprehensive perspective in synthesis, characterization, application and its challenges. J. Genet. Eng. Biotechnol. 2020, 18, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Suzuki, Y.; Huang, M.; Cao, F.; Xie, X.; Connolly, A.J.; Yang, P.C.; Wu, J.C. Comparison of Reporter Gene and Iron Particle Labeling for Tracking Fate of Human Embryonic Stem Cells and Differentiated Endothelial Cells in Living Subjects. Stem Cells 2008, 26, 864–873. [Google Scholar] [CrossRef] [Green Version]
- Naumova, A.V.; Velde, G.V. Genetically encoded iron-associated proteins as MRI reporters for molecular and cellular imaging. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2018, 10, e1482. [Google Scholar] [CrossRef]
- Chasteen, N.D.; Harrison, P.M. Mineralization in Ferritin: An Efficient Means of Iron Storage. J. Struct. Biol. 1999, 126, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdés-Stauber, N.; Scherer, S. Isolation and characterization of Linocin M18, a bacteriocin produced by Brevibacterium linens. Appl. Environ. Microbiol. 1994, 60, 3809–3814. [Google Scholar] [CrossRef] [Green Version]
- Rosenkrands, I.; Rasmussen, P.B.; Carnio, M.; Jacobsen, S.; Theisen, M.; Andersen, P. Identification and Characterization of a 29-Kilodalton Protein from Mycobacterium tuberculosis Culture Filtrate Recognized by Mouse Memory Effector Cells. Infect. Immun. 1998, 66, 2728–2735. [Google Scholar] [CrossRef] [Green Version]
- Hicks, P.M.; Rinker, K.D.; Baker, J.R.; Kelly, R.M. Homomultimeric Protease in the Hyperthermophilic Bacterium Thermotoga Maritima Has Structural and Amino Acid Sequence Homology to Bacteriocins in Mesophilic Bacteria. FEBS Lett. 1998, 440, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Giessen, T.; Silver, P.A. Converting a Natural Protein Compartment into a Nanofactory for the Size-Constrained Synthesis of Antimicrobial Silver Nanoparticles. ACS Synth. Biol. 2016, 5, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Rahmanpour, R.; Bugg, T.D.H. Assembly in Vitro of Rhodococcus Jostii RHA1 Encapsulin and Peroxidase DypB to Form a Nanocompartment. FEBS J. 2013, 280, 2097–2104. [Google Scholar] [CrossRef]
- Gabashvili, A.N.; Chmelyuk, N.S.; Efremova, M.V.; Malinovskaya, J.A.; Semkina, A.S.; Abakumov, M.A. Encapsulins—Bacterial Protein Nanocompartments: Structure, Properties, and Application. Biomolecules 2020, 10, 966. [Google Scholar] [CrossRef] [PubMed]
- McHugh, C.A.; Fontana, J.; Nemecek, D.; Cheng, N.; Aksyuk, A.A.; Heymann, B.; Winkler, D.; Lam, A.S.; Wall, J.S.; Steven, A.C.; et al. A virus capsid-like nanocompartment that stores iron and protects bacteria from oxidative stress. EMBO J. 2014, 33, 1896–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigmund, F.; Pettinger, S.; Kube, M.; Schneider, F.; Schifferer, M.; Schneider, S.; Efremova, M.V.; Pujol-Martí, J.; Aichler, M.; Walch, A.; et al. Iron-Sequestering Nanocompartments as Multiplexed Electron Microscopy Gene Reporters. ACS Nano 2019, 13, 8114–8123. [Google Scholar] [CrossRef] [PubMed]
- Efremova, M.; Bodea, S.-V.; Sigmund, F.; Semkina, A.; Westmeyer, G.; Abakumov, M. Genetically Encoded Self-Assembling Iron Oxide Nanoparticles as a Possible Platform for Cancer-Cell Tracking. Pharmaceutics 2021, 13, 397. [Google Scholar] [CrossRef]
- Sigmund, F.; Massner, C.; Erdmann, P.; Stelzl, A.; Rolbieski, H.; Desai, M.; Bricault, S.; Wörner, T.P.; Snijder, J.; Geerlof, A.; et al. Bacterial encapsulins as orthogonal compartments for mammalian cell engineering. Nat. Commun. 2018, 9, 1990. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, K.; Tammik, C.; Rosendahl, K.; Zetterberg, E.; Ringdén, O. HLA Expression and Immunologic Properties of Differentiated and Undifferentiated Mesenchymal Stem Cells. Exp. Hematol. 2003, 31, 890–896. [Google Scholar] [CrossRef]
- Park, Y.-B.; Ha, C.-W.; Lee, C.-H.; Yoon, Y.C. Cartilage Regeneration in Osteoarthritic Patients by a Composite of Allogeneic Umbilical Cord Blood-Derived Mesenchymal Stem Cells and Hyaluronate Hydrogel: Results from a Clinical Trial for Safety and Proof-of-Concept with 7 Years of Extended Follow-Up. Stem cells Transl. Med. 2016, 6, 613–621. [Google Scholar] [CrossRef]
- Human Mesenchymal Stromal Cells for Acute Respiratory Distress Syndrome (START). Available online: https://clinicaltrials.gov/ct2/show/NCT02097641 (accessed on 9 November 2021).
- Investigation of Mesenchymal Stem Cell Therapy for the Treatment of Osteoarthritis of the Knee. Available online: https://clinicaltrials.gov/ct2/show/NCT02958267 (accessed on 9 November 2021).
- Autologous Mesenchymal Bone Marrow Stromal Cells Secreting Neurotrophic Factors (MSC-NTF), in Patients with Amyotrophic Lateral Sclerosis (ALS). Available online: https://clinicaltrials.gov/ct2/show/NCT01777646 (accessed on 9 November 2021).
- Autologous Cultured Mesenchymal Bone Marrow Stromal Cells Secreting Neurotrophic Factors (MSC-NTF), in ALS Patients. Available online: https://clinicaltrials.gov/ct2/show/NCT01051882 (accessed on 9 November 2021).
- Hamann, A.; Nguyen, A.; Pannier, A.K. Nucleic acid delivery to mesenchymal stem cells: A review of nonviral methods and applications. J. Biol. Eng. 2019, 13, 1–16. [Google Scholar] [CrossRef]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, X.; Lin, F.; Xiong, M.; Fan, D.; Yuan, X.; Lu, Y.; Song, Y.; Zhang, Y.; Hao, M.; et al. Bispecific CD3-HAC carried by E1A-engineered mesenchymal stromal cells against metastatic breast cancer by blocking PD-L1 and activating T cells. J. Hematol. Oncol. 2019, 12, 46. [Google Scholar] [CrossRef]
- Suresh, S.C.; Selvaraju, V.; Thirunavukkarasu, M.; Goldman, J.; Husain, A.; Palesty, J.A.; Sanchez, J.A.; McFadden, D.W.; Maulik, N. Thioredoxin-1 (Trx1) engineered mesenchymal stem cell therapy increased pro-angiogenic factors, reduced fibrosis and improved heart function in the infarcted rat myocardium. Int. J. Cardiol. 2015, 201, 517–528. [Google Scholar] [CrossRef]
- Relation, T.; Yi, T.; Guess, A.J.; La Perle, K.; Otsuru, S.; Hasgur, S.; Dominici, M.; Breuer, C.; Horwitz, E.M. Intratumoral Delivery of Interferonγ-Secreting Mesenchymal Stromal Cells Repolarizes Tumor-Associated Macrophages and Suppresses Neuroblastoma Proliferation In Vivo. Stem Cells 2018, 36, 915–924. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.H.; Kim, K.S.; Park, S.H.; Suh, Y.S.; Kim, S.J.; Jeun, S.-S.; Sung, Y.C. The effects of mesenchymal stem cells injected via different routes on modified IL-12-mediated antitumor activity. Gene Ther. 2011, 18, 488–495. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.; Iolascon, A.; Taher, A.; Cappellini, M.D. Clinical management of iron deficiency anemia in adults: Systemic review on advances in diagnosis and treatment. Eur. J. Intern. Med. 2017, 42, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Yiannikourides, A.; Latunde-Dada, G.O. A Short Review of Iron Metabolism and Pathophysiology of Iron Disorders. Medicines 2019, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabashvili, A.N.; Vodopyanov, S.S.; Chmelyuk, N.S.; Sarkisova, V.A.; Fedotov, K.A.; Efremova, M.V.; Abakumov, M.A. Encapsulin Based Self-Assembling Iron-Containing Protein Nanoparticles for Stem Cells MRI Visualization. Int. J. Mol. Sci. 2021, 22, 12275. https://doi.org/10.3390/ijms222212275
Gabashvili AN, Vodopyanov SS, Chmelyuk NS, Sarkisova VA, Fedotov KA, Efremova MV, Abakumov MA. Encapsulin Based Self-Assembling Iron-Containing Protein Nanoparticles for Stem Cells MRI Visualization. International Journal of Molecular Sciences. 2021; 22(22):12275. https://doi.org/10.3390/ijms222212275
Chicago/Turabian StyleGabashvili, Anna N., Stepan S. Vodopyanov, Nelly S. Chmelyuk, Viktoria A. Sarkisova, Konstantin A. Fedotov, Maria V. Efremova, and Maxim A. Abakumov. 2021. "Encapsulin Based Self-Assembling Iron-Containing Protein Nanoparticles for Stem Cells MRI Visualization" International Journal of Molecular Sciences 22, no. 22: 12275. https://doi.org/10.3390/ijms222212275