
Circulating miRNAs as Potential Biomarkers Distinguishing Relapsing–Remitting from Secondary Progressive Multiple Sclerosis. A Review

 , , ,
, , ,
Abstract
:1. Introduction
2. Biogenesis and Characteristics of miRNA
3. MiRNAs as Potential Biomarkers in MS
4. Potential Candidates for a Panel Distinguishing the RR from SP Phase
5. Limitations in miRNA Biomarker Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, A.J.; Baranzini, S.E.; Geurts, J.; Hemmer, B.; Ciccarelli, O. Multiple Sclerosis. Lancet 2018, 391, 1622–1636. [Google Scholar] [PubMed]
- Walton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; la Rocca, N.; Uitdehaag, B.; van der Mei, I.; et al. Rising Prevalence of Multiple Sclerosis Worldwide: Insights from the Atlas of MS, Third Edition. Mult. Scler. J. 2020, 26, 1816–1821. [Google Scholar] [CrossRef]
- Miclea, A.; Salmen, A.; Zoehner, G.; Diem, L.; Kamm, C.P.; Chaloulos-Iakovidis, P.; Miclea, M.; Briner, M.; Kilidireas, K.; Stefanis, L.; et al. Age-dependent Variation of Female Preponderance across Different Phenotypes of Multiple Sclerosis: A Retrospective Cross-sectional Study. CNS Neurosci. Ther. 2019, 25, 527–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilha, I.G.; Fonseca, A.P.A.; Pettengill, A.L.M.; Fragoso, D.C.; Pacheco, F.T.; Nunes, R.H.; Maia, A.C.M.; da Rocha, A.J. Pediatric Multiple Sclerosis: From Clinical Basis to Imaging Spectrum and Differential Diagnosis. Pediatric Radiol. 2020, 50, 776–792. [Google Scholar]
- Roohani, P.; Emiru, T.; Carpenter, A.; Luzzio, C.; Freeman, J.; Scarberry, S.; Beaver, G.; Davidson, L.; Parry, G. Late Onset Multiple Sclerosis: Is It Really Late Onset? Mult. Scler. Relat. Disord. 2014, 3, 444–449. [Google Scholar] [CrossRef]
- GBD 2016 Multiple Sclerosis Collaborators. Global, Regional, and National Burden of Multiple Sclerosis 1990-2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet. Neurol. 2019, 18, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.V.; Pearson, J.F.; Clarke, G.; Mason, D.F.; Abernethy, D.A.; Willoughby, E.; Sabel, C. MS Prevalence in New Zealand, an Ethnically and Latitudinally Diverse Country. Mult. Scler. 2010, 16, 1422–1431. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Chard, D.; Ciccarelli, O. Clinically Isolated Syndromes. Lancet. Neurol. 2012, 11, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Klineova, S.; Lublin, F.D. Clinical Course of Multiple Sclerosis. Cold Spring Harb. Perspect. Med. 2018, 8, a028928. [Google Scholar] [CrossRef]
- Lassmann, H. Multiple Sclerosis Pathology. Cold Spring Harb. Perspect. Med. 2018, 8, a028936. [Google Scholar] [CrossRef] [Green Version]
- Lucchinetti, C.; Brück, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of Multiple Sclerosis Lesions: Implications for the Pathogenesis of Demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Kuhlmann, T.; Ludwin, S.; Prat, A.; Antel, J.; Brück, W.; Lassmann, H. An Updated Histological Classification System for Multiple Sclerosis Lesions. Acta Neuropathol. 2017, 133, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bitsch, A.; Brück, W. Differentiation of Multiple Sclerosis Subtypes: Implications for Treatment. CNS Drugs 2002, 16, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Goverman, J. Autoimmune T Cell Responses in the Central Nervous System. Nat. Rev. Immunol. 2009, 9, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Kasper, L.H.; Shoemaker, J. Multiple Sclerosis Immunology: The Healthy Immune System vs. the MS Immune System. Neurology 2010, 74, S2. [Google Scholar] [CrossRef]
- Kalra, S.; Lowndes, C.; Durant, L.; Strange, R.; Al-Araji, A.; Hawkins, C.P.; Curnow, S.J. Th17 Cells Increase in RRMS as Well as in SPMS, Whereas Various Other Phenotypes of Th17 Increase in RRMS Only. Mult. Scler. J. Exp. Transl. Clin. 2020, 6, 2055217319899695. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.; Rosenberg, G.A. Matrix Metalloproteinase-Mediated Disruption of Tight Junction Proteins in Cerebral Vessels Is Reversed by Synthetic Matrix Metalloproteinase Inhibitor in Focal Ischemia in Rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Murphy, Á.C.; Lalor, S.J.; Lynch, M.A.; Mills, K.H.G. Infiltration of Th1 and Th17 Cells and Activation of Microglia in the CNS during the Course of Experimental Autoimmune Encephalomyelitis. Brain Behav. Immun. 2010, 24, 641–651. [Google Scholar] [CrossRef]
- Confavreux, C.; Vukusic, S. The Clinical Course of Multiple Sclerosis. Handb. Clin. Neurol. 2014, 122, 343–369. [Google Scholar] [CrossRef]
- Compston, A.; Coles, A. Multiple Sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Frischer, J.M.; Bramow, S.; Dal-Bianco, A.; Lucchinetti, C.F.; Rauschka, H.; Schmidbauer, M.; Laursen, H.; Sorensen, P.S.; Lassmann, H. The Relation between Inflammation and Neurodegeneration in Multiple Sclerosis Brains. Brain 2009, 132, 1175–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrazzano, G.; Crisafulli, S.G.; Baione, V.; Tartaglia, M.; Cortese, A.; Frontoni, M.; Altieri, M.; Pauri, F.; Millefiorini, E.; Conte, A. Early Diagnosis of Secondary Progressive Multiple Sclerosis: Focus on Fluid and Neurophysiological Biomarkers. J. Neurol. 2021, 268, 3626–3645. [Google Scholar] [CrossRef] [PubMed]
- Hollen, C.W.; Soldán, M.M.P.; Rinker, J.R., II; Spain, R.I. The Future of Progressive Multiple Sclerosis Therapies. Fed. Pract. 2020, 37, S43. [Google Scholar] [PubMed]
- de Angelis, F.; Plantone, D.; Chataway, J. Pharmacotherapy in Secondary Progressive Multiple Sclerosis: An Overview. CNS Drugs 2018, 32, 499–526. [Google Scholar] [CrossRef]
- Adamczyk-Sowa, M.; Adamczyk, B.; Kułakowska, A.; Rejdak, K.; Nowacki, P. Secondary Progressive Multiple Sclerosis—From Neuropathology to Definition and Effective Treatment. Neurol. I Neurochir. Pol. 2020, 54, 384–398. [Google Scholar] [CrossRef]
- Gross, H.J.; Watson, C. Characteristics, Burden of Illness, and Physical Functioning of Patients with Relapsing-Remitting and Secondary Progressive Multiple Sclerosis: A Cross-Sectional US Survey. Neuropsychiatr. Dis. Treat. 2017, 13, 1349–1357. [Google Scholar] [CrossRef] [Green Version]
- Inojosa, H.; Proschmann, U.; Akgün, K.; Ziemssen, T. A Focus on Secondary Progressive Multiple Sclerosis (SPMS): Challenges in Diagnosis and Definition. J. Neurol. 2021, 268, 1210–1221. [Google Scholar] [CrossRef] [PubMed]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sørensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the Clinical Course of Multiple Sclerosis: The 2013 Revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ontaneda, D.; Cohen, J.; Amato, M. Clinical Outcome Measures for Progressive MS Trials. Mult. Scler. 2017, 23, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of Multiple Sclerosis: 2017 Revisions of the McDonald Criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Calabrese, M.; Favaretto, A.; Martini, V.; Gallo, P. Grey Matter Lesions in MS: From Histology to Clinical Implications. Prion 2013, 7, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trip, S.; Miller, D. Imaging in Multiple Sclerosis. J. Neurol. Neurosurg. Psychiatry 2005, 76, iii11. [Google Scholar] [CrossRef] [Green Version]
- Housley, W.; Pitt, D.; Hafler, D. Biomarkers in Multiple Sclerosis. Clin. Immunol. 2015, 161, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Preziosa, P.; Rocca, M. Microstructural MR Imaging Techniques in Multiple Sclerosis. Neuroimaging Clin. N. Am. 2017, 27, 313–333. [Google Scholar] [CrossRef]
- Pasquali, L.; Lucchesi, C.; Pecori, C.; Metelli, M.R.; Pellegrini, S.; Iudice, A.; Bonuccelli, U. A Clinical and Laboratory Study Evaluating the Profile of Cytokine Levels in Relapsing Remitting and Secondary Progressive Multiple Sclerosis. J. Neuroimmunol. 2015, 278, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Gresle, M.M.; Liu, Y.; Dagley, L.F.; Haartsen, J.; Pearson, F.; Purcell, A.W.; Laverick, L.; Petzold, A.; Lucas, R.M.; van der Walt, A.; et al. Serum Phosphorylated Neurofilament-Heavy Chain Levels in Multiple Sclerosis Patients. J. Neurol. Neurosurg. Psychiatry 2014, 85, 1209–1213. [Google Scholar] [CrossRef]
- Ayrignac, X.; le Bars, E.; Duflos, C.; Hirtz, C.; Maceski, A.M.; Carra-Dallière, C.; Charif, M.; Pinna, F.; Prin, P.; de Champfleur, N.M.; et al. Serum GFAP in Multiple Sclerosis: Correlation with Disease Type and MRI Markers of Disease Severity. Sci. Rep. 2020, 10, 10923. [Google Scholar] [CrossRef] [PubMed]
- Högel, H.; Rissanen, E.; Barro, C.; Matilainen, M.; Nylund, M.; Kuhle, J.; Airas, L. Serum Glial Fibrillary Acidic Protein Correlates with Multiple Sclerosis Disease Severity. Mult. Scler. 2018, 26, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Harris, V.K.; Sadiq, S.A. Disease Biomarkers in Multiple Sclerosis. Mol. Diagn. Ther. 2012, 13, 225–244. [Google Scholar] [CrossRef]
- Zhou, Z.; Xiong, H.; Xie, F.; Wu, Z.; Feng, Y. A Meta-Analytic Review of the Value of MiRNA for Multiple Sclerosis Diagnosis. Front. Neurol. 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Han, D.; Feng, J. MicroRNA in Multiple Sclerosis. Clin. Chim. Acta 2021, 516, 92–99. [Google Scholar] [CrossRef]
- Ferrante, M.; Conti, G.O. Environment and Neurodegenerative Diseases: An Update on MiRNA Role. MicroRNA 2017, 6. [Google Scholar] [CrossRef]
- Martinez, B.; Peplow, P. MicroRNAs in Blood and Cerebrospinal Fluid as Diagnostic Biomarkers of Multiple Sclerosis and to Monitor Disease Progression. Neural Regen. Res. 2020, 15, 606–619. [Google Scholar] [CrossRef]
- Qiu, L.; Tan, E.; Zeng, L. MicroRNAs and Neurodegenerative Diseases. Adv. Exp. Med. Biol. 2015, 888, 51–70. [Google Scholar] [CrossRef]
- Dolati, S.; Babaloo, Z.; Jadidi-Niaragh, F.; Ayromlou, H.; Sadreddini, S.; Yousefi, M. Multiple Sclerosis: Therapeutic Applications of Advancing Drug Delivery Systems. Biomed. Pharmacother. = Biomed. Pharmacother. 2017, 86, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Ajit, S.K. Circulating MicroRNAs as Biomarkers, Therapeutic Targets, and Signaling Molecules. Sensors 2012, 12, 3359–3369. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating MicroRNAs as Stable Blood-Based Markers for Cancer Detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Sheinerman, K.S.; Umansky, S.R. Circulating Cell-Free MicroRNA as Biomarkers for Screening, Diagnosis, and Monitoring of Neurodegenerative Diseases and Other Neurologic Pathologies. Front. Cell. Neurosci. 2013, 7, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghildiyal, M.; Xu, J.; Seitz, H.; Weng, Z.; Zamore, P.D. Sorting of Drosophila Small Silencing RNAs Partitions MicroRNA* Strands into the RNA Interference Pathway. RNA 2010, 16, 43–56. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu-Gruttadauria, J.; MacRae, I.J. Structural Foundations of RNA Silencing by Argonaute. J. Mol. Biol. 2017, 429, 2619–2639. [Google Scholar] [CrossRef]
- Desvignes, T.; Batzel, P.; Berezikov, E.; Eilbeck, K.; Eppig, J.T.; McAndrews, M.S.; Singer, A.; Postlethwait, J.H. MicroRNA Nomenclature: A View Incorporating Genetic Origins, Biosynthetic Pathways, and Sequence Variants. Trends Genet. TIG 2015, 31, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing Beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef] [Green Version]
- Denzler, R.; McGeary, S.E.; Title, A.C.; Agarwal, V.; Bartel, D.P.; Stoffel, M. Impact of MicroRNA Levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression. Mol. Cell 2016, 64, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.-J.; Leng, X.-M. Dynamic MiRNA–MRNA Paradigms: New Faces of MiRNAs. Biochem. Biophys. Rep. 2015, 4, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread Changes in Protein Synthesis Induced by MicroRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E. Targeting of MRNAs by Multiple MiRNAs: The next Step. Oncogene 2010, 29, 2161–2164. [Google Scholar] [CrossRef] [Green Version]
- Dedeoğlu, B.G. High-Throughput Approaches for MicroRNA Expression Analysis. Methods Mol. Biol. 2014, 1107, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA Dysregulation in Cancer: Diagnostics, Monitoring and Therapeutics. A Comprehensive Review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-Time Quantification of MicroRNAs by Stem–Loop RT–PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zheng, J.; Chen, Z.; Liu, Y.; Dura, B.; Kwak, M.; Xavier-Ferrucio, J.; Lu, Y.-C.; Zhang, M.; Roden, C.; et al. Single-Cell MicroRNA-MRNA Co-Sequencing Reveals Non-Genetic Heterogeneity and Mechanisms of MicroRNA Regulation. Nat. Commun. 2019, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Liu, C.; Tong, H.; Chen, Y.; Liu, K. Principles of Digital PCR and Its Applications in Current Obstetrical and Gynecological Diseases. Am. J. Transl. Res. 2019, 11, 7209–7222. [Google Scholar] [PubMed]
- Redis, R.S.; Calin, S.; Yang, Y.; You, M.J.; Calin, G.A. Cell-to-Cell MiRNA Transfer: From Body Homeostasis to Therapy. Pharmacol. Ther. 2012, 136, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [Green Version]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Pirola, C.J.; Gianotti, T.F.; Castaño, G.O.; Mallardi, P.; Martino, J.S.; Ledesma, M.M.G.L.; Flichman, D.; Mirshahi, F.; Sanyal, A.J.; Sookoian, S. Circulating MicroRNA Signature in Non-Alcoholic Fatty Liver Disease: From Serum Non-Coding RNAs to Liver Histology and Disease Pathogenesis. Gut 2015, 64, 800–812. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Shen, Z. Exosomal MiRNAs as Biomarkers for Diagnostic and Prognostic in Lung Cancer. Cancer Med. 2020, 9, 6909–6922. [Google Scholar] [CrossRef]
- Manna, I.; Iaccino, E.; Dattilo, V.; Barone, S.; Vecchio, E.; Mimmi, S.; Filippelli, E.; Demonte, G.; Polidoro, S.; Granata, A.; et al. Exosome-Associated MiRNA Profile as a Prognostic Tool for Therapy Response Monitoring in Multiple Sclerosis Patients. FASEB J. 2018, 32, 4241–4246. [Google Scholar] [CrossRef] [Green Version]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum MicroRNAs Are Promising Novel Biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Nakajima, G.; Gavin, E.; Morris, C.G.; Kudo, K.; Hayashi, K.; Ju, J. Systematic Analysis of MicroRNA Expression of RNA Extracted from Fresh Frozen and Formalin-Fixed Paraffin-Embedded Samples. RNA 2007, 13, 1668–1674. [Google Scholar] [CrossRef] [Green Version]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. MiRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Correia, C.N.; Nalpas, N.C.; McLoughlin, K.E.; Browne, J.A.; Gordon, S.V.; MacHugh, D.E.; Shaughnessy, R.G. Circulating MicroRNAs as Potential Biomarkers of Infectious Disease. Front. Immunol. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating High Confidence MicroRNAs Using Deep Sequencing Data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, V.; Winkler, J. MiRNA-Based Therapies: Strategies and Delivery Platforms for Oligonucleotide and Non-Oligonucleotide Agents. Future Med. Chem. 2014, 6, 1967. [Google Scholar] [CrossRef] [Green Version]
- HMDD v3.2. Available online: https://www.cuilab.cn/hmdd (accessed on 6 September 2021).
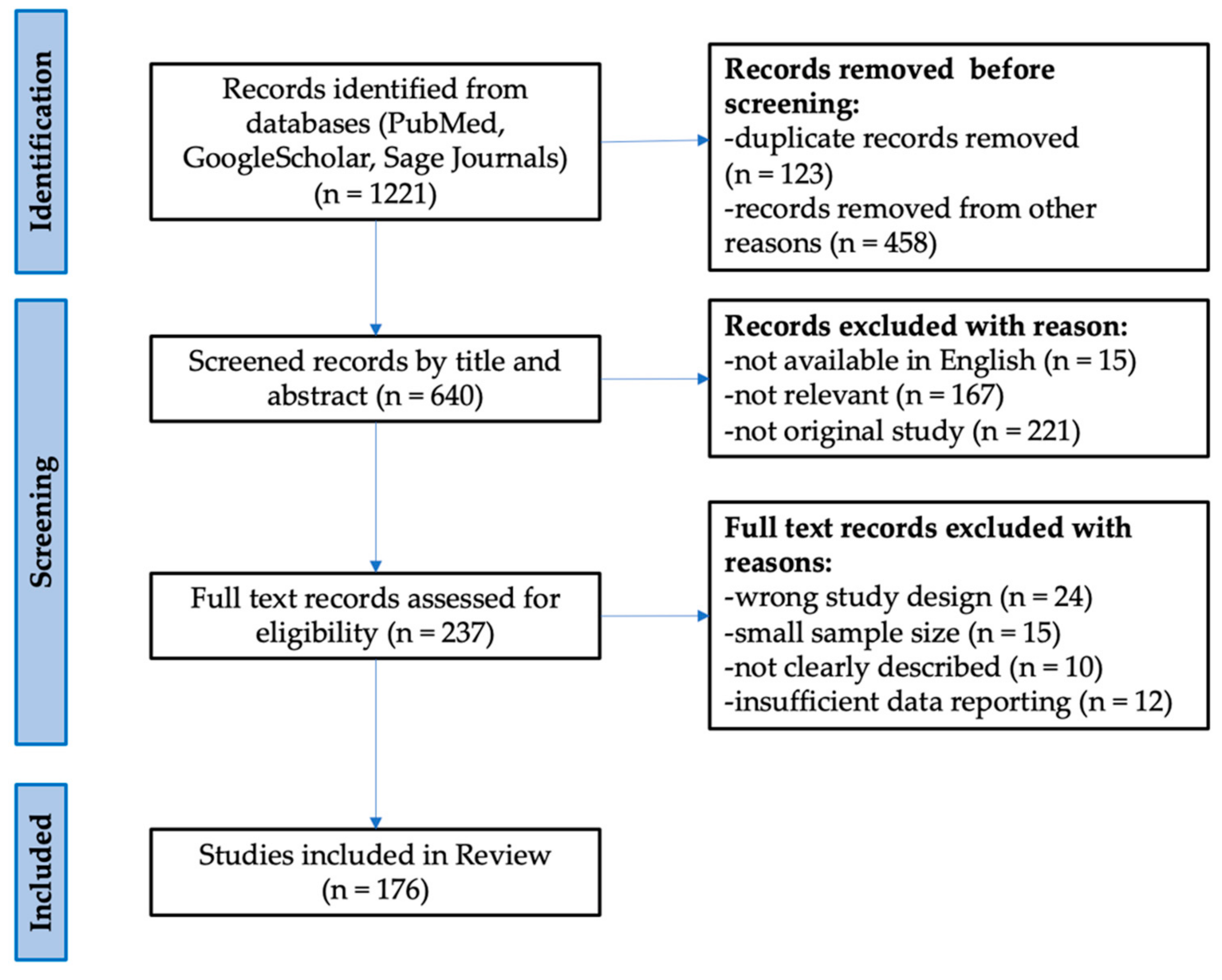
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Nejad, C.; Stunden, H.J.; Gantier, M.P. A Guide to MiRNAs in Inflammation and Innate Immune Responses. FEBS J. 2018, 285, 3695–3716. [Google Scholar] [CrossRef]
- Mendell, J.T.; Olson, E.N. MicroRNAs in Stress Signaling and Human Disease. Cell 2012, 148, 1172–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar] [PubMed]
- Li, Y.; Kowdley, K. MicroRNAs in Common Human Diseases. Genom. Proteom. Bioinform. 2012, 10, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Sheinerman, K.S.; Toledo, J.B.; Tsivinsky, V.G.; Irwin, D.; Grossman, M.; Weintraub, D.; Hurtig, H.I.; Chen-Plotkin, A.; Wolk, D.A.; McCluskey, L.F.; et al. Circulating Brain-Enriched MicroRNAs as Novel Biomarkers for Detection and Differentiation of Neurodegenerative Diseases. Alzheimer’s Res. Ther. 2017, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.-Y. MicroRNAs in Human Diseases: From Autoimmune Diseases to Skin, Psychiatric and Neurodegenerative Diseases. Immune Netw. 2011, 11, 227–244. [Google Scholar] [CrossRef]
- Patterson, M.; Gaeta, X.; Loo, K.; Edwards, M.; Smale, S.; Cinkornpumin, J.; Xie, Y.; Listgarten, J.; Azghadi, S.; Douglass, S.M.; et al. Let-7 MiRNAs Can Act through Notch to Regulate Human Gliogenesis. Stem Cell Rep. 2014, 3, 758–773. [Google Scholar] [CrossRef] [Green Version]
- Emery, B. Regulation of Oligodendrocyte Differentiation and Myelination. Science 2010, 330, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Janowska, J.; Gargas, J.; Ziemka-Nalecz, M.; Zalewska, T.; Buzanska, L.; Sypecka, J. Directed Glial Differentiation and Transdifferentiation for Neural Tissue Regeneration. Exp. Neurol. 2019, 319, 112813. [Google Scholar] [CrossRef] [PubMed]
- Vistbakka, J.; Elovaara, I.; Lehtimäki, T.; Hagman, S. Circulating MicroRNAs as Biomarkers in Progressive Multiple Sclerosis. Mult. Scler. 2016, 23, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Piket, E.; Zheleznyakova, G.; Kular, L.; Jagodic, M. Small Non-Coding RNAs as Important Players, Biomarkers and Therapeutic Targets in Multiple Sclerosis: A Comprehensive Overview. J. Autoimmun. 2019, 101, 17–25. [Google Scholar] [CrossRef]
- Gandhi, R. MiRNA in Multiple Sclerosis: Search for Novel Biomarkers. Mult. Scler. J. 2015, 21, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Otaegui, D.; Baranzini, S.E.; Armañanzas, R.; Calvo, B.; Muñoz-Culla, M.; Khankhanian, P.; Inza, I.; Lozano, J.A.; Castillo-Triviño, T.; Asensio, A.; et al. Differential Micro RNA Expression in PBMC from Multiple Sclerosis Patients. PLoS ONE 2009, 4, e6309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regev, K.; Paul, A.; Healy, B.; von Glenn, F.; Diaz-Cruz, C.; Gholipour, T.; Antonietta Mazzola, M.; Raheja, R.; Nejad, P.; Glanz, B.I.; et al. Comprehensive Evaluation of Serum MicroRNAs as Biomarkers in Multiple Sclerosis. Neurol. Neuroimmunol. NeuroInflammation 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Mandolesi, G.; Rizzo, F.R.; Balletta, S.; Bassi, M.S.; Gilio, L.; Guadalupi, L.; Nencini, M.; Moscatelli, A.; Ryan, C.P.; Licursi, V.; et al. The MicroRNA Let-7b-5p Is Negatively Associated with Inflammation and Disease Severity in Multiple Sclerosis. Cells 2021, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Perdaens, O.; Dang, H.A.; D’Auria, L.; van Pesch, V. CSF MicroRNAs Discriminate MS Activity and Share Similarity to Other Neuroinflammatory Disorders. Neurol. Neuroimmunol. Neuroinflammation 2020, 7, e673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baulina, N.M.; Kulakova, O.G.; Favorova, O.O. MicroRNAs: The Role in Autoimmune Inflammation. Acta Nat. 2016, 8, 21–33. [Google Scholar] [CrossRef]
- Maciak, K.; Dziedzic, A.; Miller, E.; Saluk-Bijak, J. MiR-155 as an Important Regulator of Multiple Sclerosis Pathogenesis. A Review. Int. J. Mol. Sci. 2021, 22, 4332. [Google Scholar] [CrossRef]
- McCoy, C.E. MiR-155 Dysregulation and Therapeutic Intervention in Multiple Sclerosis. Adv. Exp. Med. Biol. 2017, 1024, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Hong, W.; Wang, X.; Zhang, P.; Körner, H.; Tu, J.; Wei, W. MicroRNAs in Microglia: How Do MicroRNAs Affect Activation, Inflammation, Polarization of Microglia and Mediate the Interaction Between Microglia and Glioma? Front. Mol. Neurosci. 2019, 12, 125. [Google Scholar] [CrossRef]
- Tarassishin, L.; Loudig, O.; Bauman, A.; Shafit-Zagardo, B.; Suh, H.-S.; Lee, S.C. Interferon Regulatory Factor 3 Inhibits Astrocyte Inflammatory Gene Expression through Suppression of the Proinflammatory MiR-155 and MiR-155. Glia 2011, 59, 1911–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.R.; Kang, Y.H.; Jeon, H.; Lee, S.; Park, S.-J.; Song, D.-Y.; Min, S.S.; Yoo, S.-M.; Lee, M.-S.; Lee, S.-H. Differential Expression of MiRNAs and Behavioral Change in the Cuprizone-Induced Demyelination Mouse Model. Int. J. Mol. Sci. 2020, 21, 646. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Zhang, G.; Li, S. Astrogliosis and Axonal Regeneration. Neural Regen. 2015, 181–196. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, B.; Sun, Y.; Xie, X. MiR-155 Modulates the Progression of Neuropathic Pain through Targeting SGK3. Int. J. Clin. Exp. Pathol. 2015, 8, 14374. [Google Scholar]
- Kacperska, M.J.; Jastrzebski, K.; Tomasik, B.; Walenczak, J.; Konarska-Krol, M.; Glabinski, A. Selected Extracellular MicroRNA as Potential Biomarkers of Multiple Sclerosis Activity—Preliminary Study. J. Mol. Neurosci. 2015, 56, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, R.; Healy, B.; Gholipour, T.; Egorova, S.; Musallam, A.; Hussain, M.S.; Nejad, P.; Patel, B.; Hei, H.; Khoury, S.; et al. Circulating MicroRNAs as Biomarkers for Disease Staging in Multiple Sclerosis. Ann. Neurol. 2013, 73, 729–740. [Google Scholar] [CrossRef]
- Bernstein, D.L.; Jiang, X.; Rom, S. Let-7 MicroRNAs: Their Role in Cerebral and Cardiovascular Diseases, Inflammation, Cancer, and Their Regulation. Biomedicines 2021, 9, 606. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Song, J.; Oh, Y.; Lee, J.E. MicroRNA-Let-7a Regulates the Function of Microglia in Inflammation. Mol. Cell. Neurosci. 2015, 68, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Falcone, M.; Rajan, A.; Bloom, B.; Brosnan, C. A Critical Role for IL-4 in Regulating Disease Severity in Experimental Allergic Encephalomyelitis as Demonstrated in IL-4-Deficient C57BL/6 Mice and BALB/c Mice. J. Immunol. 1998, 160, 4822–4830. [Google Scholar]
- Talebi, F.; Ghourbani, S.; Vojgani, M.; Noorbakhsh, F. MiR-320 and Inflammation Regulation in Experimental Autoimmune Encephalomyelitis through Interference With Tumor Growth Factor-β Signaling Pathway. Immunoregulation 2020, 2, 111–120. [Google Scholar] [CrossRef]
- Lee, P.W.; Severin, M.E.; Lovett-Racke, A.E. TGF-β Regulation of T Cells in Multiple Sclerosis. Eur. J. Immunol. 2017, 47, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Aung, L.L.; Mouradian, M.M.; Dhib-Jalbut, S.; Balashov, K.E. MMP-9 Expression Is Increased in B Lymphocytes during Multiple Sclerosis Exacerbation and Is Regulated by MicroRNA-320a. J. Neuroimmunol. 2015, 278, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Fainardi, E.; Castellazzi, M.; Bellini, T.; Manfrinato, M.C.; Baldi, E.; Casetta, I.; Paolino, E.; Granieri, E.; Dallocchio, F. Cerebrospinal Fluid and Serum Levels and Intrathecal Production of Active Matrix Metalloproteinase-9 (MMP-9) as Markers of Disease Activity in Patients with Multiple Sclerosis. Mult. Scler. 2016, 12, 294–301. [Google Scholar] [CrossRef]
- Castellazzi, M.; Ligi, D.; Contaldi, E.; Quartana, D.; Fonderico, M.; Borgatti, L.; Bellini, T.; Trentini, A.; Granieri, E.; Fainardi, E.; et al. Multiplex Matrix Metalloproteinases Analysis in the Cerebrospinal Fluid Reveals Potential Specific Patterns in Multiple Sclerosis Patients. Front. Neurol. 2018, 9, 1080. [Google Scholar] [CrossRef] [PubMed]
- A Pilot Study to Assess MicroRNA Biomarkers in Early and Later Stage Multiple Sclerosis—Full Text View—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01737372 (accessed on 6 September 2021).
- Zou, X.; Wei, J.; Huang, Z.; Zhou, X.; Lu, Z.; Zhu, W.; Miao, Y. Identification of a Six-miRNA Panel in Serum Benefiting Pancreatic Cancer Diagnosis. Cancer Med. 2019, 8, 2810–2822. [Google Scholar] [CrossRef]
- Yan, L.; Zhao, W.; Yu, H.; Wang, Y.; Liu, Y.; Xie, C. A Comprehensive Meta-Analysis of MicroRNAs for Predicting Colorectal Cancer. Medicine 2016, 95, e2738. [Google Scholar] [CrossRef] [PubMed]
- Pogribny, I.P. MicroRNAs as Biomarkers for Clinical Studies. Exp. Biol. Med. 2018, 243, 283. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; Haghikia, A.; Hellwig, K.; Baraniskin, A.; Holzmann, A.; Décard, B.F.; Thum, T.; Gold, R. Regulated MicroRNAs in the CSF of Patients with Multiple Sclerosis. Neurology 2012, 79, 2166–2170. [Google Scholar] [CrossRef]
- Kramer, S.; Haghikia, A.; Bang, C.; Scherf, K.; Pfanne, A.; Duscha, A.; Kaisler, J.; Gisevius, B.; Gold, R.; Thum, T.; et al. Elevated Levels of MiR-181c and MiR-633 in the CSF of Patients with MS: A Validation Study. Neurol. Neuroimmunol. Neuroinflammation 2019, 6, e623. [Google Scholar] [CrossRef] [Green Version]
- Ahlbrecht, J.; Martino, F.; Pul, R.; Skripuletz, T.; Sühs, K.-W.; Schauerte, C.; Yildiz, Ö.; Trebst, C.; Tasto, L.; Thum, S.; et al. Deregulation of MicroRNA-181c in Cerebrospinal Fluid of Patients with Clinically Isolated Syndrome Is Associated with Early Conversion to Relapsing–Remitting Multiple Sclerosis. Mult. Scler. 2015, 22, 1202–1214. [Google Scholar] [CrossRef]
- Kos, A.; Loohuis, N.O.; Meinhardt, J.; van Bokhoven, H.; Kaplan, B.B.; Martens, G.; Aschrafi, A. MicroRNA-181 Promotes Synaptogenesis and Attenuates Axonal Outgrowth in Cortical Neurons. Cell. Mol. Life Sci. CMLS 2016, 73, 3555–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Xue, Z.; Liu, Y.; Liu, H.; Guo, X.; Li, Y.; Yang, H.; Zhang, L.; Da, Y.; Yao, Z.; et al. MicroRNA-181c Promotes Th17 Cell Differentiation and Mediates Experimental Autoimmune Encephalomyelitis. Brain Behav. Immun. 2018, 70, 305–314. [Google Scholar] [CrossRef]
- Mackey, M.F.; Barth, R.J.; Noelle, R.J. The Role of CD40/CD154 Interactions in the Priming, Differentiation, and Effector Function of Helper and Cytotoxic T Cells. J. Leukoc. Biol. 1998, 63, 418–428. [Google Scholar] [CrossRef]
- Karnell, J.L.; Rieder, S.A.; Ettinger, R.; Kolbeck, R. Targeting the CD40-CD40L Pathway in Autoimmune Diseases: Humoral Immunity and Beyond. Adv. Drug Deliv. Rev. 2019, 141, 92–103. [Google Scholar] [CrossRef]
- Aarts, S.A.B.M.; Seijkens, T.T.P.; van Dorst, K.J.F.; Dijkstra, C.D.; Kooij, G.; Lutgens, E. The CD40–CD40L Dyad in Experimental Autoimmune Encephalomyelitis and Multiple Sclerosis. Front. Immunol. 2017, 8, 1791. [Google Scholar] [CrossRef] [Green Version]
- Balashov, K.E.; Smith, D.R.; Khoury, S.J.; Hafler, D.A.; Weiner, H.L. Increased Interleukin 12 Production in Progressive Multiple Sclerosis: Induction by Activated CD4+ T Cells via CD40 Ligand. Proc. Natl. Acad. Sci. USA 1997, 94, 599–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei, N.; Talebi, F.; Ghorbani, S.; Rezaei, A.; Esmaeili, A.; Noorbakhsh, F.; Hakemi, M.G. MicroRNA-92a Drives Th1 Responses in the Experimental Autoimmune Encephalomyelitis. Inflammation 2018, 42, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Xie, N.; Cui, H.; Tan, Z.; Yang, S.; Icyuz, M.; Abraham, E.; Liu, G. MicroRNA Let-7c Regulates Macrophage Polarization. J. Immunol. 2013, 190, 6542–6549. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Gregersen, P.K.; Diamond, B. Regulation of Dendritic Cell Activation by MicroRNA Let-7c and BLIMP1. J. Clin. Investig. 2013, 123, 823–833. [Google Scholar] [CrossRef]
- Liu, H.Y.; Huang, C.M.; Hung, Y.F.; Hsueh, Y.P. The MicroRNAs Let7c and MiR21 Are Recognized by Neuronal Toll-like Receptor 7 to Restrict Dendritic Growth of Neurons. Exp. Neurol. 2015, 269, 202–212. [Google Scholar] [CrossRef]
- Søndergaard, H.B.; Hesse, D.; Krakauer, M.; Sørensen, P.S.; Sellebjerg, F. Differential MicroRNA Expression in Blood in Multiple Sclerosis. Mult. Scler. 2013, 19, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- De, R.; Malefyt, W.; Abrams, J.; Bennett, B.; Figdor, C.G.; de Vries, J.E. Interleukin 10(EL,.10) Inhibits Cytokine Synthesis by Human Monocytes: An Autoregulatory Role of IL-10 Produced by Monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Al-Faraj, S. Neurothrophins. Molecular Links between the Immune and Nervous Systems. Neurosci. J. 2001, 6, 23–25. [Google Scholar]
- Sarchielli, P.; Greco, L.; Stipa, A.; Floridi, A.; Gallai, V. Brain-Derived Neurotrophic Factor in Patients with Multiple Sclerosis. J. Neuroimmunol. 2002, 132, 180–188. [Google Scholar] [CrossRef]
- Saha, R.N.; Jana, M.; Pahan, K. MAPK P38 Regulates Transcriptional Activity of NF-ΚB in Primary Human Astrocytes via Acetylation of P65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef] [Green Version]
- Torres, J.; Enríquez-de-Salamanca, A.; Fernández, I.; Rodríguez-Ares, M.T.; Quadrado, M.J.; Murta, J.; del Castillo, J.M.B.; Stern, M.E.; Calonge, M. Activation of MAPK Signaling Pathway and NF-ΚB Activation in Pterygium and Ipsilateral Pterygium-Free Conjunctival Specimens. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5842–5852. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhou, N.; Yang, P.; Deng, L.; Liu, G. MicroRNA-27a-3p Downregulation Inhibits Inflammatory Response and Hippocampal Neuronal Cell Apoptosis by Upregulating Mitogen-Activated Protein Kinase 4 (MAP2K4) Expression in Epilepsy: In Vivo and In Vitro Studies. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 8499–8508. [Google Scholar] [CrossRef]
- Ahmadian-Elmi, M.; Bidmeshki Pour, A.; Naghavian, R.; Ghaedi, K.; Tanhaei, S.; Izadi, T.; Nasr-Esfahani, M.H. MiR-27a and MiR-214 Exert Opposite Regulatory Roles in Th17 Differentiation via Mediating Different Signaling Pathways in Peripheral Blood CD4+ T Lymphocytes of Patients with Relapsing–Remitting Multiple Sclerosis. Immunogenetics 2016, 68, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.A.; Howell, M.; Page, C.S.; Daly, A.; Bates, P.A.; Hill, C.S. Recognition of Phosphorylated-Smad2-Containing Complexes by a Novel Smad Interaction Motif. Mol. Cell. Biol. 2004, 24, 1106–1121. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Lopes, J.E.; Chong, M.M.W.; Ivanov, I.I.; Min, R.; Victora, G.D.; Shen, Y.; Du, J.; Rubtsov, Y.P.; Rudensky, A.Y.; et al. TGF-β-Induced Foxp3 Inhibits Th17 Cell Differentiation by Antagonizing RORγt Function. Nature 2008, 453, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Volsko, C.; Garcia, J.P.; Agirre, E.; Allan, K.C.; Tesar, P.J.; Trapp, B.D.; Castelo-Branco, G.; Sim, F.J.; Dutta, R. Oligodendrocyte Intrinsic MiR-27a Controls Myelination and Remyelination. Cell Rep. 2019, 29, 904–919. [Google Scholar] [CrossRef] [Green Version]
- Regev, K.; Healy, B.C.; Paul, A.; Diaz-Cruz, C.; Mazzola, M.A.; Raheja, R.; Glanz, B.I.; Kivisäkk, P.; Chitnis, T.; Jagodic, M.; et al. Identification of MS-Specific Serum MiRNAs in an International Multicenter Study. Neurol. Neuroimmunol. NeuroInflammation 2018, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, B.; Kappos, L. Natalizumab: Targeting A4-Integrins in Multiple Sclerosis. Neurodegener. Dis. 2008, 5, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Salinas, G.F.; Krausz, S.; Dontje, W.; Evavold, B.D.; Tak, P.P.; Baeten, D.L.; Reedquist, K.A. Sustained Rap1 Activation in Autoantigen-Specific T Lymphocytes Attenuates Experimental Autoimmune Encephalomyelitis. J. Neuroimmunol. 2012, 250, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Sharaf-Eldin, W.E.; Kishk, N.A.; Gad, Y.Z.; Hassan, H.; Ali, M.A.M.; Zaki, M.S.; Mohamed, M.R.; Essawi, M.L. Extracellular MiR-145, MiR-223 and MiR-326 Expression Signature Allow for Differential Diagnosis of Immune-Mediated Neuroinflammatory Diseases. J. Neurol. Sci. 2017, 383, 188–198. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The Meaning and Use of the Area under a Receiver Operating Characteristic (ROC) Curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornfeld, S.F.; Cummings, S.E.; Fathi, S.; Bonin, S.R.; Kothary, R. MiRNA-145-5p Prevents Differentiation of Oligodendrocyte Progenitor Cells by Regulating Expression of Myelin Gene Regulatory Factor. J. Cell. Physiol. 2021, 236, 997–1012. [Google Scholar] [CrossRef] [PubMed]
- Dharap, A.; Bowen, K.; Place, R.; Li, L.-C.; Vemuganti, R. Transient Focal Ischemia Induces Extensive Temporal Changes in Rat Cerebral MicroRNAome. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2009, 29, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Peng, L.; Zhu, J.; Zhou, Y.; Li, L.; Chen, Y.; Yu, S.; Zhao, Y. MiR-145-5p/Nurr1/TNF-α Signaling-Induced Microglia Activation Regulates Neuron Injury of Acute Cerebral Ischemic/Reperfusion in Rats. Front. Mol. Neurosci. 2017, 10, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Barcelos, I.P.; Troxell, R.M.; Graves, J.S. Mitochondrial Dysfunction and Multiple Sclerosis. Biology 2019, 8, 37. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wei, Z.-C.; Xu, L.-L.; Yu, S.-Y.; Li, C. Inhibition of MiR-145-5p Reduces Spinal Cord Injury-Induced Inflammatory and Oxidative Stress Responses via Affecting Nurr1-TNF-α Signaling Axis. Cell Biochem. Biophys. 2021. [Google Scholar] [CrossRef]
- Johnnidis, J.B.; Harris, M.H.; Wheeler, R.T.; Stehling-Sun, S.; Lam, M.H.; Kirak, O.; Brummelkamp, T.R.; Fleming, M.D.; Camargo, F.D. Regulation of Progenitor Cell Proliferation and Granulocyte Function by MicroRNA-223. Nature 2008, 451, 1125–1129. [Google Scholar] [CrossRef]
- Ifergan, I.; Chen, S.; Zhang, B.; Miller, S.D. Cutting Edge: MicroRNA-223 Regulates Myeloid Dendritic Cell–Driven Th17 Responses in Experimental Autoimmune Encephalomyelitis. J. Immunol. 2016, 196, 1455–1459. [Google Scholar] [CrossRef]
- Galloway, D.A.; Blandford, S.N.; Berry, T.; Williams, J.B.; Stefanelli, M.; Ploughman, M.; Moore, C.S. MiR-223 Promotes Regenerative Myeloid Cell Phenotype and Function in the Demyelinated Central Nervous System. Glia 2019, 67, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Morgan, M.J.; Choksi, S.; Zhang, Y.; Kim, Y.S.; Liu, Z.G. MicroRNAs Modulate the Noncanonical Transcription Factor NF-ΚB Pathway by Regulating Expression of the Kinase IKKα during Macrophage Differentiation. Nat. Immunol. 2010, 11, 799–805. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Pal, A.S.; Hsu, A.Y.H.; Gurol, T.; Zhu, X.; Wirbisky-Hershberger, S.E.; Freeman, J.L.; Kasinski, A.L.; Deng, Q. MicroRNA-223 Suppresses the Canonical NF-ΚB Pathway in Basal Keratinocytes to Dampen Neutrophilic Inflammation. Cell Rep. 2018, 22, 1810–1823. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wang, H.; Liu, Y.; Song, Y.; Lai, L.; Han, Q.; Cao, X.; Wang, Q. Inducible MicroRNA-223 down-Regulation Promotes TLR-Triggered IL-6 and IL-1β Production in Macrophages by Targeting STAT3. PLoS ONE 2012, 7, e42971. [Google Scholar] [CrossRef] [Green Version]
- Barclay, W.; Shinohara, M.L. Inflammasome Activation in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis (EAE). Brain Pathol. 2017, 27, 213–219. [Google Scholar] [CrossRef]
- Bauernfeind, F.; Rieger, A.; Schildberg, F.A.; Knolle, P.A.; Schmid-Burgk, J.L.; Hornung, V. NLRP3 Inflammasome Activity Is Negatively Controlled by MiR-223. J. Immunol. 2012, 189, 4175–4181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, S.; Costa, C.; Eixarch, H.; Keller, C.W.; Amman, L.; Martínez-Banaclocha, H.; Midaglia, L.; Sarró, E.; Machín-Díaz, I.; Villar, L.M.; et al. NLRP3 Inflammasome as Prognostic Factor and Therapeutic Target in Primary Progressive Multiple Sclerosis Patients. Brain 2020, 143, 1414–1430. [Google Scholar] [CrossRef] [PubMed]
- Morquette, B.; Juźwik, C.A.; Drake, S.S.; Charabati, M.; Zhang, Y.; Lécuyer, M.A.; Galloway, D.A.; Dumas, A.; de Faria Junior, O.; Paradis-Isler, N.; et al. MicroRNA-223 Protects Neurons from Degeneration in Experimental Autoimmune Encephalomyelitis. Brain 2019, 142, 2979–2995. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Vissel, B. The Essential Role of AMPA Receptor GluA2 Subunit RNA Editing in the Normal and Diseased Brain. Front. Mol. Neurosci. 2012, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Liu, C.; Kang, J.; Zhao, G.; Ye, Z.; Huang, S.; Li, Z.; Wu, Z.; Pei, G. MicroRNA MiR-326 Regulates TH-17 Differentiation and Is Associated with the Pathogenesis of Multiple Sclerosis. Nat. Immunol. 2009, 10, 1252–1259. [Google Scholar] [CrossRef]
- Moisan, J.; Grenningloh, R.; Bettelli, E.; Oukka, M.; Ho, I.-C. Ets-1 Is a Negative Regulator of Th17 Differentiation. J. Exp. Med. 2007, 204, 2825–2835. [Google Scholar] [CrossRef] [PubMed]
- Honardoost, M.A.; Kiani-Esfahani, A.; Ghaedi, K.; Etemadifar, M.; Salehi, M. MiR-326 and MiR-26a, Two Potential Markers for Diagnosis of Relapse and Remission Phases in Patient with Relapsing–Remitting Multiple Sclerosis. Gene 2014, 544, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Woo, J.H.; Jou, I.; Joe, E. PINK1 Deficiency Decreases Expression Levels of Mir-326, Mir-330, and Mir-3099 during Brain Development and Neural Stem Cell Differentiation. Exp. Neurobiol. 2016, 25, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Junker, A.; Krumbholz, M.; Eisele, S.; Mohan, H.; Augstein, F.; Bittner, R.; Lassmann, H.; Wekerle, H.; Hohlfeld, R.; Meinl, E. MicroRNA Profiling of Multiple Sclerosis Lesions Identifies Modulators of the Regulatory Protein CD47. Brain 2009, 132, 3342–3352. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; Beadnall, H.; Barnett, M.H.; Suter, C.M.; Buckland, M.E. Exosomal MicroRNA Signatures in Multiple Sclerosis Reflect Disease Status. Sci. Rep. 2017, 7, 14293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Shao, R.; Li, M.; Yan, Q.; Hu, H. MiR-485-3p Modulates Neural Stem Cell Differentiation and Proliferation via Regulating TRIP6 Expression. J. Cell. Mol. Med. 2020, 24, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Li, H.; Liu, W.; Zhang, L.; Tian, Q.; Li, H.; Li, M. MiR-485-3p Serves as a Biomarker and Therapeutic Target of Alzheimer’s Disease via Regulating Neuronal Cell Viability and Neuroinflammation by Targeting AKT3. Mol. Genet. Genom. Med. 2021, 9, e1548. [Google Scholar] [CrossRef]
- Lin, X.; Wang, R.; Li, R.; Tao, T.; Zhang, D.; Qi, Y. Diagnostic Performance of MiR-485-3p in Patients with Parkinson’s Disease and Its Relationship with Neuroinflammation. Neuromolecular Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fordham, J.B.; Naqvi, A.R.; Nares, S. Regulation of MiR-24, MiR-30b, and MiR-142-3p during Macrophage and Dendritic Cell Differentiation Potentiates Innate Immunity. J. Leukoc. Biol. 2015, 98, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Brennan, S.; Keon, M.; Liu, B.; Su, Z.; Saksena, N.K. Panoramic Visualization of Circulating MicroRNAs Across Neurodegenerative Diseases in Humans. Mol. Neurobiol. 2019, 56, 7380–7407. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Li, Z.; Zhang, G.-X.; Guan, Y. Wnt Signaling in Remyelination in Multiple Sclerosis: Friend or Foe? Mol. Neurobiol. 2013, 49, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, Z.; Tu, Y.; Shen, H.; Dai, Z.; Lin, J.; Zhou, Z. MiR-101a and MiR-30b Contribute to Inflammatory Cytokine-Mediated β-Cell Dysfunction. Lab. Investig. 2015, 95, 1387–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharief, M.K.; Matthews, H.; Noori, M.A. Expression Ratios of the Bcl-2 Family Proteins and Disease Activity in Multiple Sclerosis. J. Neuroimmunol. 2003, 134, 158–165. [Google Scholar] [CrossRef]
- Zettl, U.K.; Kuhlmann, T.; Brück, W. Bcl-2 Expressing T Lymphocytes in Multiple Sclerosis Lesions. Neuropathol. Appl. Neurobiol. 1998, 24, 202–208. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
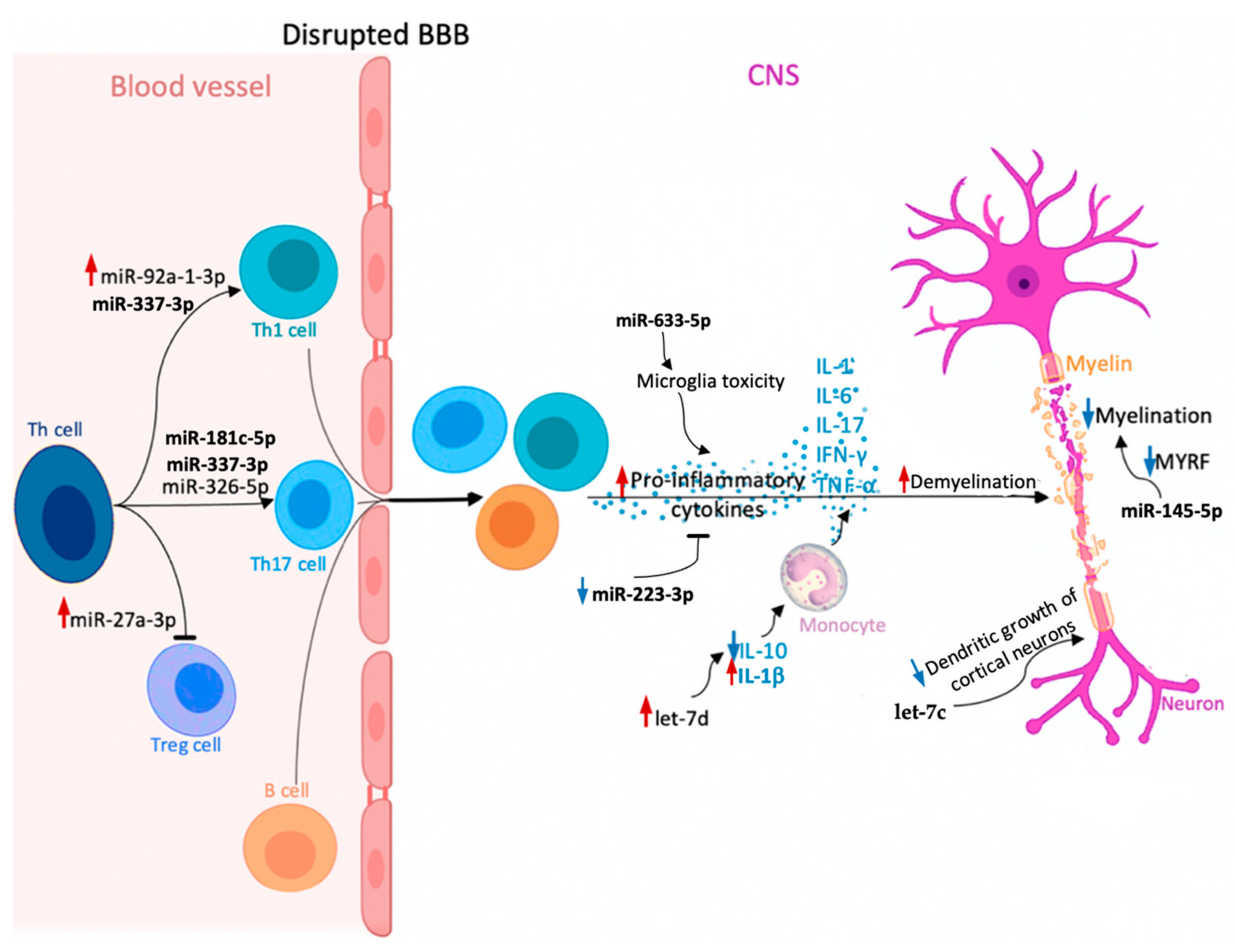
| miRNA | Function | SPMS vs. RRMS | Biological Material | Patients Number | miRNAs Number Included in the Analysis | Ref. |
|---|---|---|---|---|---|---|
| miR-181c-5p | Regulates neuronal maturation and synaptogenesis in the cortex | Upregulated | CSF | 81 RRMS 106 SPMS 0 HCs 211 OND | 3 | [118] |
| Downregulated | 17 RRMS 30 SPMS 0 HCs 39 OND | 760 | [117] | |||
| miR-633-5p | Targets mRNAs transcripts involved in neuroinflammatory pathways | Downregulated | CSF | 17 RRMS 30 SPMS 0 HCs 39 OND | 760 | [117] |
| miR-27a-3p | Regulates the differentiation of Th1 and Th17 cells and the accumulation of Tregs Regulates oligodendrocytes differentiation | Downregulated | Serum | 29 RRMS 19 SPMS 30 HCs | 652 | [92] |
| miR-337-3p | Initiates through Rap1 signaling pathogenic character of T cells | Downregulated | Serum | 115 RRMS 51 SPMS 88 HCs | 652 | [141] |
| miR-92a-1-3p | Targets CD40-CD40L pathway Regulates cell cycle and cell signaling Promotes Th1 differentiation | Downregulated | Plasma | 51 SPMS 50 RRMS 32 HCs | 368 | [104] |
| miR-145-5p | Regulates the expression of myelin gene regulatory factor | |||||
| let-7c | Plays a role in suppressing the dendritic growth of cortical neuron | |||||
| let-7d | Regulates immunological and inflammatory responses | |||||
| Combination of: miR-223-3p + miR-326-5p + miR-145-5p; miR-326-5p + miR-145-5p; miR-223-3p + miR-326-5p | miR-223-3p: Modulates the nuclear factor κB pathway Regulates inflammatory responses in macrophages | No statistical significance | Serum | 18 RRMS 19 SPMS 23 HCs | 3 | [144] |
| miR-326-5p: Regulates Th17 differentiation | Downregulated | |||||
| miR-145-5p: Regulates the expression of myelin gene regulatory factor | No statistical significance |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrasik, S.; Dziedzic, A.; Miller, E.; Starosta, M.; Saluk-Bijak, J. Circulating miRNAs as Potential Biomarkers Distinguishing Relapsing–Remitting from Secondary Progressive Multiple Sclerosis. A Review. Int. J. Mol. Sci. 2021, 22, 11887. https://doi.org/10.3390/ijms222111887
Pietrasik S, Dziedzic A, Miller E, Starosta M, Saluk-Bijak J. Circulating miRNAs as Potential Biomarkers Distinguishing Relapsing–Remitting from Secondary Progressive Multiple Sclerosis. A Review. International Journal of Molecular Sciences. 2021; 22(21):11887. https://doi.org/10.3390/ijms222111887
Chicago/Turabian StylePietrasik, Sylwia, Angela Dziedzic, Elzbieta Miller, Michal Starosta, and Joanna Saluk-Bijak. 2021. "Circulating miRNAs as Potential Biomarkers Distinguishing Relapsing–Remitting from Secondary Progressive Multiple Sclerosis. A Review" International Journal of Molecular Sciences 22, no. 21: 11887. https://doi.org/10.3390/ijms222111887