
Fibulin 2 Is Hypermethylated and Suppresses Tumor Cell Proliferation through Inhibition of Cell Adhesion and Extracellular Matrix Genes in Non-Small Cell Lung Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
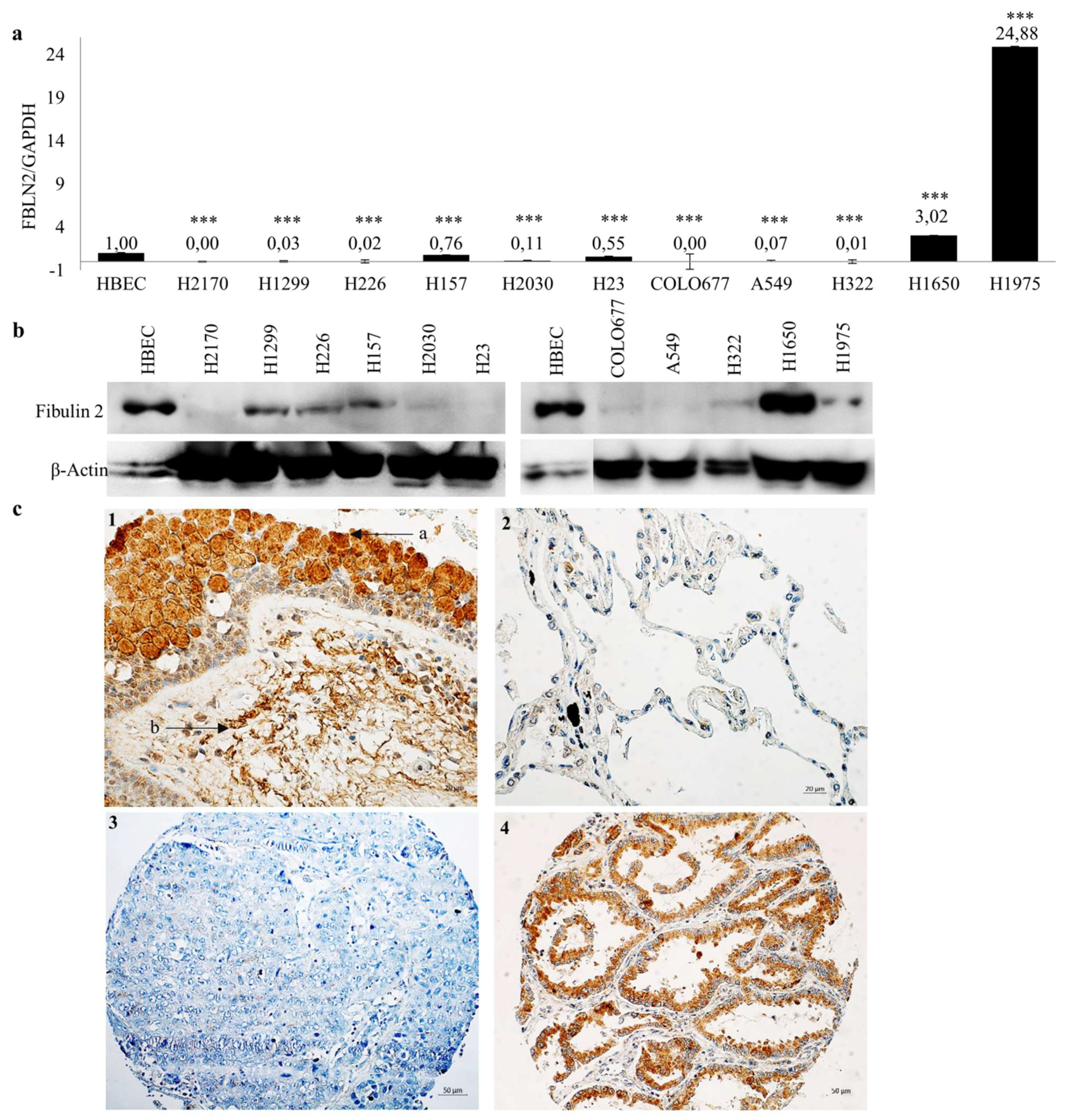
2.1. FBLN2 Is Downregulated in Lung Cancer
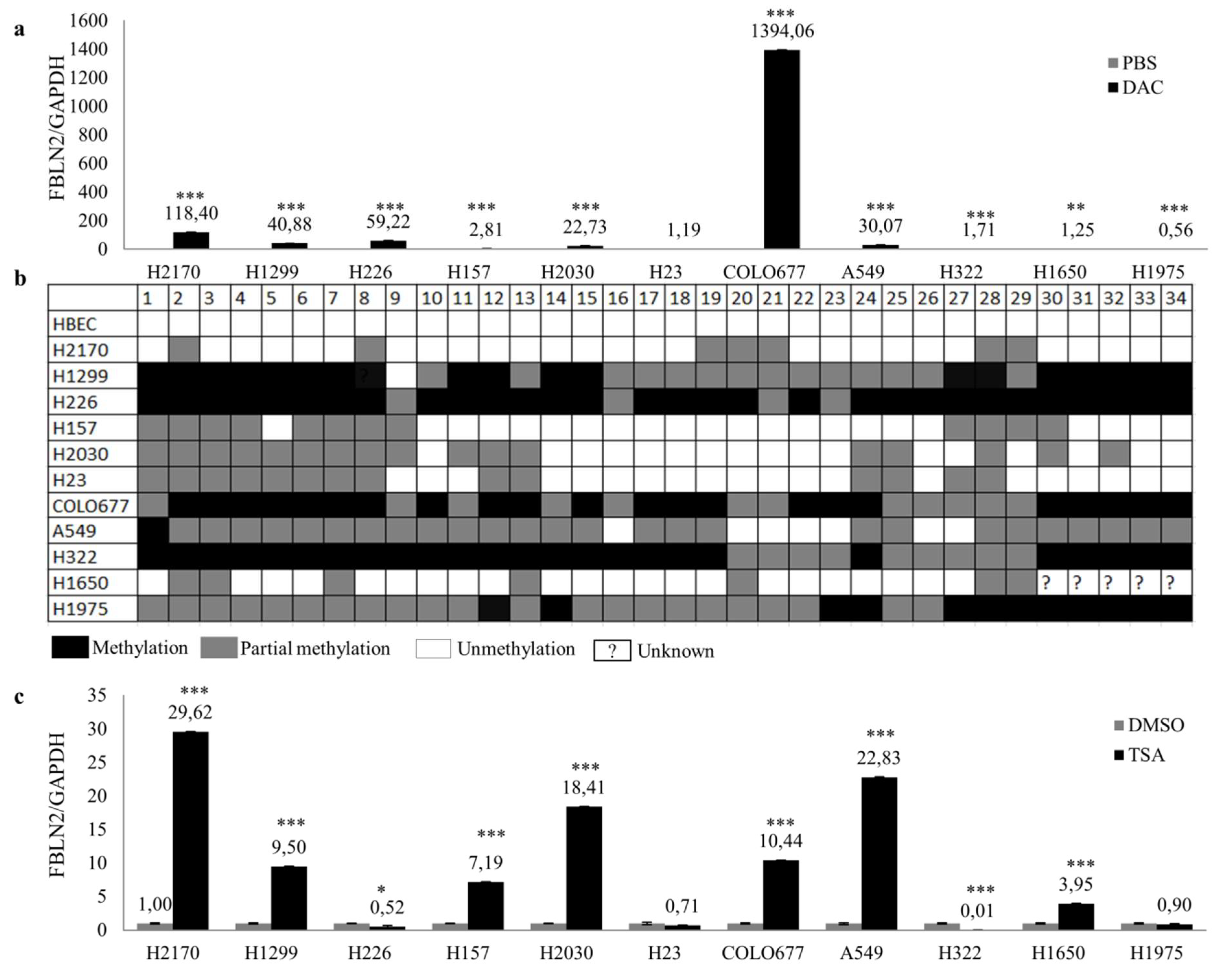
2.2. Epigenetic Regulation Contributes to the Downregulation of FBLN2
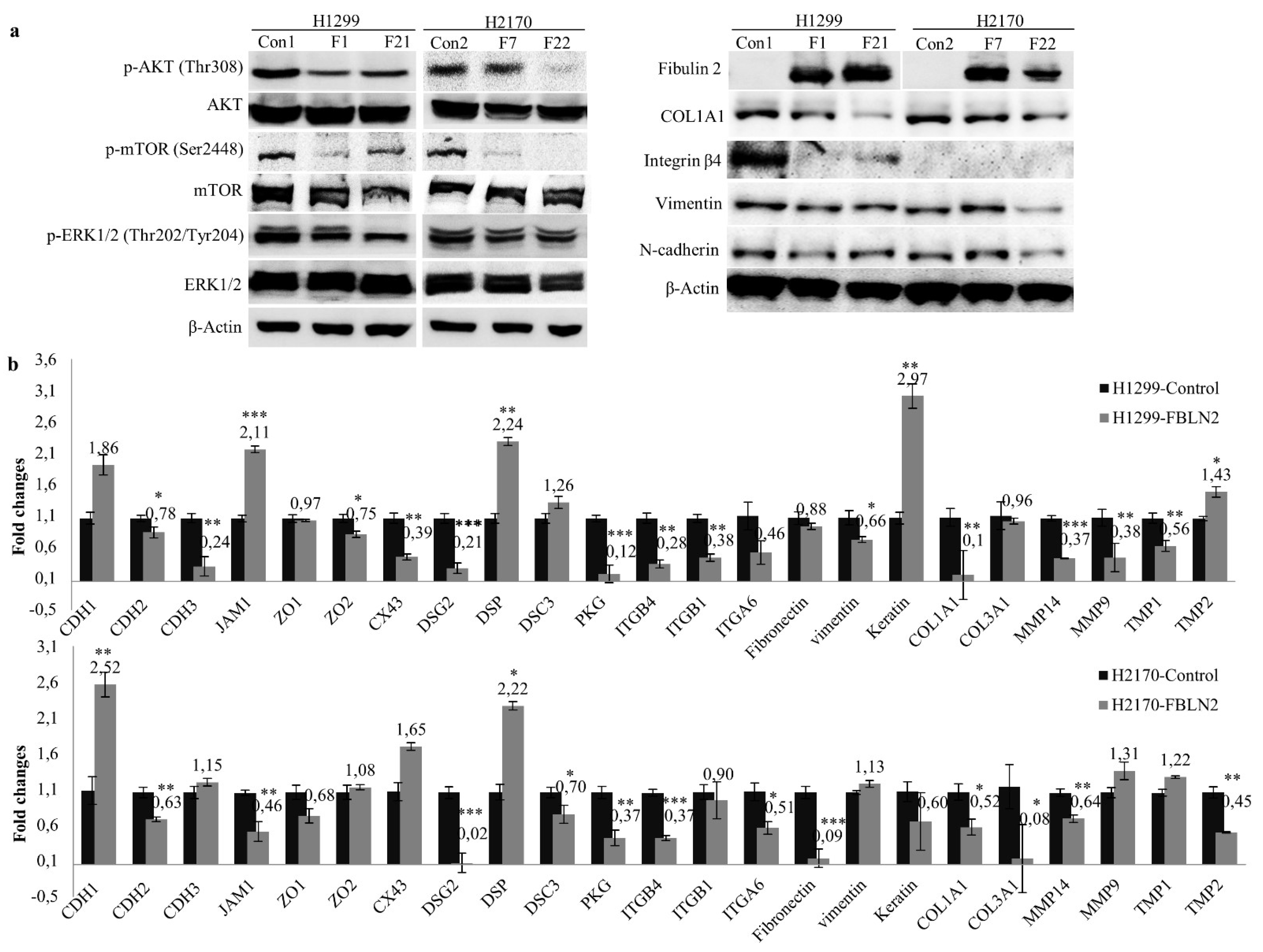
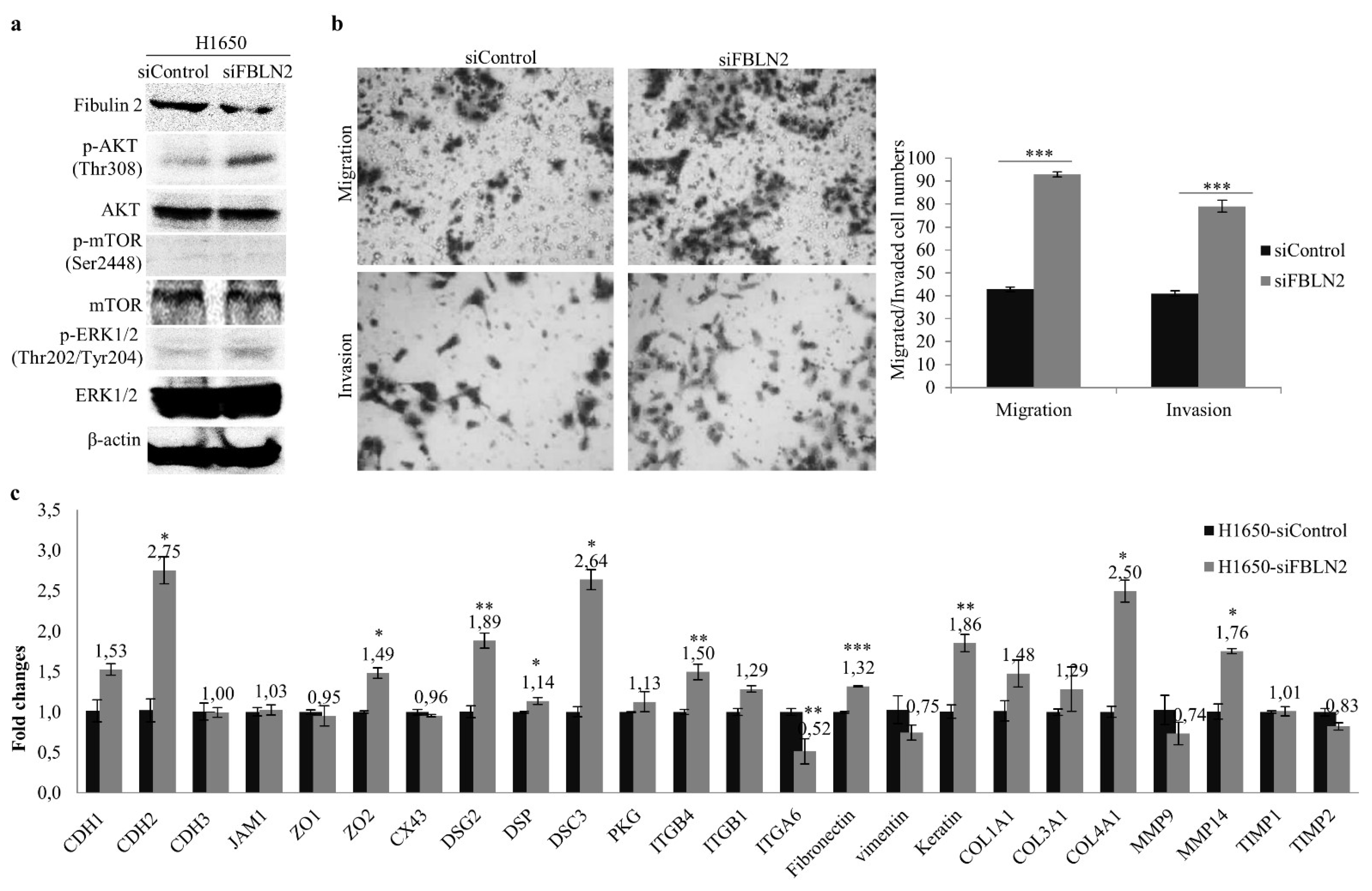
2.3. FBLN2 Suppresses Cell Colony Formation, Invasion and Migration through Inhibiting the MAPK/ERK and PI3K/AKT/mTOR Pathways
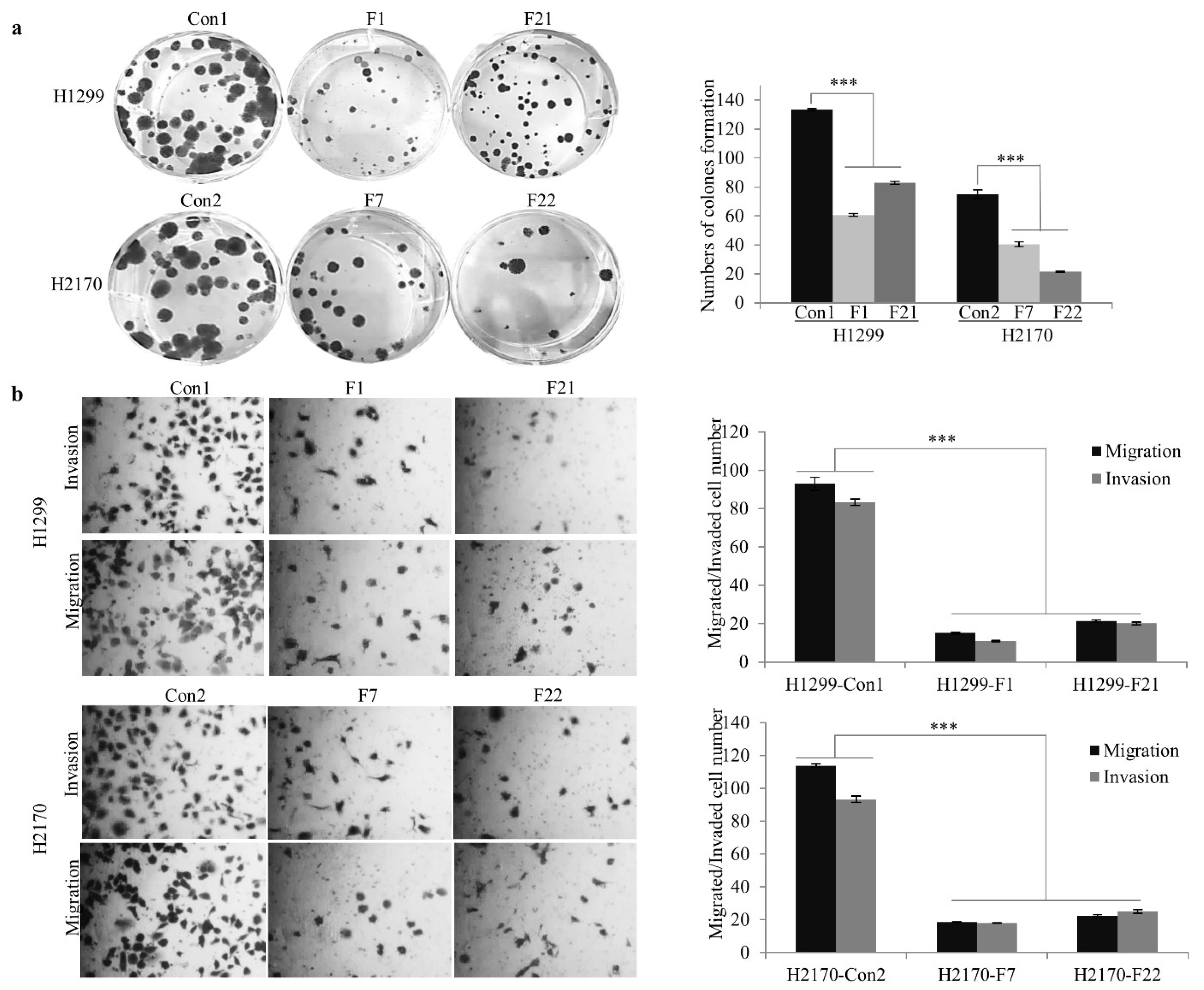
2.4. FBLN2 Suppresses Tumor Cell Proliferation by Regulation of Cell Adhesion Molecules and ECM Associated Genes
2.5. FBLN2 Knockdown Promotes Migratory and Invasive Phonotypes
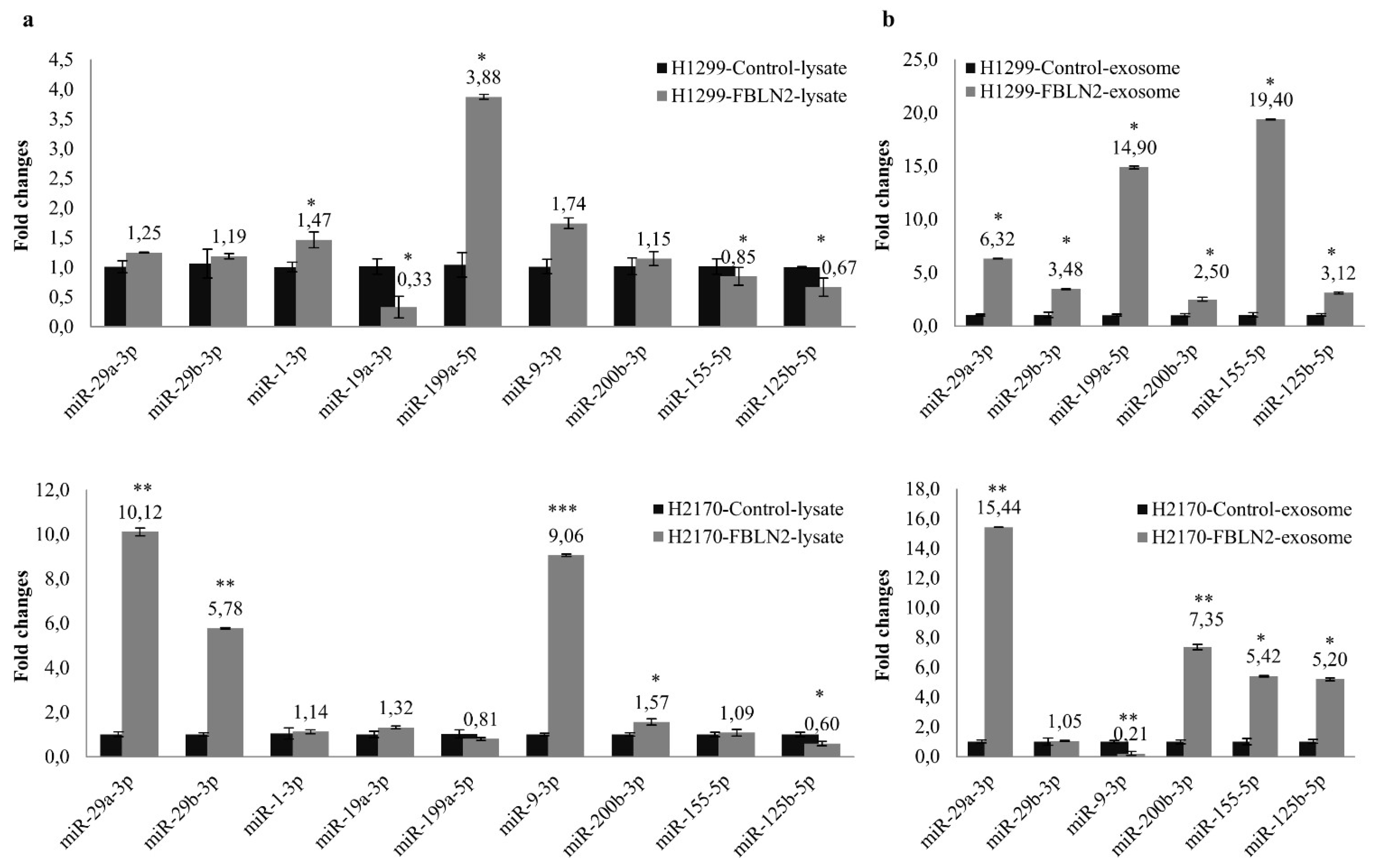
2.6. FBLN2 Modulates miRNAs in Regulation of Cell Adhesion and ECM Molecules
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Drug Treatment
4.2. RNA Isolation, cDNA Synthesis and Real-Time RT-PCR
4.3. DNA Methylation Analysis
4.4. Patient Samples and Immunohistochemistry (IHC)
4.5. FBLN2 Expression Vector Construction, Stable Transfection, and siRNA Knockdown
4.6. Colony Formation, Migration, and Invasion
4.7. Western Blot (WB) Analysis
4.8. Exosome Isolation, miRNA Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| MET | Mesenchymal–epithelial transition |
| CDH1 | E-cadherin |
| CDH2 | N-cadherin |
| CDH3 | P-cadherin |
| JAM | Junctional adhesion molecule |
| ZO | Zonula occludens |
| CX | Connexin 43 (Gap junction alpha-1) |
| ITGB | Integrin beta |
| ITGA | Integrin alpha |
| ELN | Elastin |
| COL1A1 | Collagen Type 1 alpha 1 |
| MMP | Matrix metallopeptidase |
| TIMP | Metallopeptidase inhibitor |
| DSP | Desmoplakin |
| DSG2 | Desmoglein 2 |
| DSC2 | Desmocollin 3 |
| PKG | Plakoglobin |
References
- Obaya, A.J.; Rua, S.; Moncada-Pazos, A.; Cal, S. The dual role of fibulins in tumorigenesis. Cancer Lett. 2012, 325, 132–138. [Google Scholar] [CrossRef]
- Papke, C.L.; Yanagisawa, H. Fibulin-4 and fibulin-5 in elastogenesis and beyond: Insights from mouse and human studies. Matrix Biol. 2014, 37, 142–149. [Google Scholar] [CrossRef]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell Sci. 2002, 115, 2817–2828. [Google Scholar] [CrossRef]
- Atlas, Human Protein. Available online: https://www.proteinatlas.org/ENSG00000163520-FBLN2/tissue (accessed on 29 September 2021).
- Sicot, F.X.; Tsuda, T.; Markova, D.; Klement, J.F.; Arita, M.; Zhang, R.Z.; Pan, T.C.; Mecham, R.P.; Birk, D.E.; Chu, M.L. Fibulin-2 is dispensable for mouse development and elastic fiber formation. Mol. Cell. Biol. 2008, 28, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, S.L.; Sicot, F.X.; Davis, E.C.; Huang, J.; Sasaki, T.; Chu, M.L.; Yanagisawa, H. Fibulin-2 and fibulin-5 cooperatively function to form the internal elastic lamina and protect from vascular injury. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Olijnyk, D.; Ibrahim, A.M.; Ferrier, R.K.; Tsuda, T.; Chu, M.L.; Gusterson, B.A.; Stein, T.; Morris, J.S. Fibulin-2 is involved in early extracellular matrix development of the outgrowing mouse mammary epithelium. Cell. Mol. Life Sci. 2014, 71, 3811–3828. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Sabet, S.; El-Ghor, A.A.; Kamel, N.; Anis, S.E.; Morris, J.S.; Stein, T. Fibulin-2 is required for basement membrane integrity of mammary epithelium. Sci. Rep. 2018, 8, 14139. [Google Scholar] [CrossRef] [PubMed]
- Missan, D.S.; Chittur, S.V.; DiPersio, C.M. Regulation of fibulin-2 gene expression by integrin α3β1 contributes to the invasive phenotype of transformed keratinocytes. J. Investig. Dermatol. 2014, 134, 2418–2427. [Google Scholar] [CrossRef] [Green Version]
- Longmate, W.M.; Monichan, R.; Chu, M.L.; Tsuda, T.; Mahoney, M.G.; DiPersio, C.M. Reduced fibulin-2 contributes to loss of basement membrane integrity and skin blistering in mice lacking integrin α3β1 in the epidermis. J. Investig. Dermatol. 2014, 134, 1609–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. Br. J. Pharmacol. 2019, 176, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, W.M.; Currid, C.A.; Whelan, L.C. Fibulins and cancer: Friend or foe? Trends Mol. Med. 2005, 11, 336–340. [Google Scholar] [CrossRef]
- Atlas, Genetics and Cytogenetics in Oncology and Haematology. Available online: http://atlasgeneticsoncology.org/Genes/GC_FBLN2.html (accessed on 29 September 2021).
- Grässel, S.; Sicot, F.X.; Gotta, S.; Chu, M.L. Mouse fibulin-2 gene. Complete exon-intron organization and promoter characterization. Eur. J. Biochem. 1999, 263, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Law, E.W.; Cheung, A.K.; Kashuba, V.I.; Pavlova, T.V.; Zabarovsky, E.R.; Lung, H.L.; Cheng, Y.; Chua, D.; Lai-Wan Kwong, D.; Tsao, S.W.; et al. Anti-angiogenic and tumor-suppressive roles of candidate tumor-suppressor gene, Fibulin-2, in nasopharyngeal carcinoma. Oncogene 2012, 31, 728–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danan-Gotthold, M.; Golan-Gerstl, R.; Eisenberg, E.; Meir, K.; Karni, R.; Levanon, E.Y. Identification of recurrent regulated alternative splicing events across human solid tumors. Nucleic Acids Res. 2015, 43, 5130–5144. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.S.; Son, H.; Kim, S.; Kim, S. ISOexpresso: A web-based platform for isoform-level expression analysis in human cancer. BMC Genom. 2016, 17, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senapati, S.; Gnanapragassam, V.S.; Moniaux, N.; Momi, N.; Batra, S.K. Role of MUC4-NIDO domain in the MUC4-mediated metastasis of pancreatic cancer cells. Oncogene 2012, 31, 3346–3356. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Lian, C.; Song, Y. Fibulin-2 inhibits development of gastric cancer by downregulating β-catenin. Oncol. Lett. 2019, 18, 2799–2804. [Google Scholar] [CrossRef] [PubMed]
- Atlas, Human Protein. Available online: https://www.proteinatlas.org/ENSG00000163520-FBLN2 (accessed on 29 September 2021).
- Sofela, A.A.; Hilton, D.A.; Ammoun, S.; Baiz, D.; Adams, C.L.; Ercolano, E.; Jenkinson, M.D.; Kurian, K.M.; Teo, M.; Whitfield, P.C.; et al. Fibulin-2: A Novel Biomarker for Differentiating Grade II from Grade I Meningiomas. Int. J. Mol. Sci. 2021, 22, 560. [Google Scholar] [CrossRef]
- Baird, B.N.; Schliekelman, M.J.; Ahn, Y.H.; Chen, Y.; Roybal, J.D.; Gill, B.J.; Mishra, D.K.; Erez, B.; O’Reilly, M.; Yang, Y.; et al. Fibulin-2 is a driver of malignant progression in lung adenocarcinoma. PLoS ONE 2013, 8, e67054. [Google Scholar] [CrossRef] [Green Version]
- Valastyan, S.; Weinberg, R.A. Roles for microRNAs in the regulation of cell adhesion molecules. J. Cell Sci. 2011, 124, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Piccinini, A.M.; Midwood, K.S. Illustrating the interplay between the extracellular matrix and microRNAs. Int. J. Exp. Pathol. 2014, 95, 158–180. [Google Scholar] [CrossRef] [Green Version]
- Boon, R.A.; Seeger, T.; Heydt, S.; Fischer, A.; Hergenreider, E.; Horrevoets, A.J.; Vinciguerra, M.; Rosenthal, N.; Sciacca, S.; Pilato, M.; et al. MicroRNA-29 in aortic dilation: Implications for aneurysm formation. Circ. Res. 2011, 109, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Hill, V.K.; Hesson, L.B.; Dansranjavin, T.; Dallol, A.; Bieche, I.; Vacher, S.; Tommasi, S.; Dobbins, T.; Gentle, D.; Euhus, D.; et al. Identification of 5 novel genes methylated in breast and other epithelial cancers. Mol. Cancer 2010, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Cully, M.; Downward, J. SnapShot: Ras Signaling. Cell 2008, 133, 1292. [Google Scholar] [CrossRef] [Green Version]
- Krutakova, M.; Sarlinova, M.; Matakova, T.; Dzian, A.; Hamzik, J.; Pec, M.; Javorkova, S.; Halasova, E. The Role of Dysregulated MicroRNA Expression in Lung Cancer. Adv. Exp. Med. Biol. 2016, 911, 1–8. [Google Scholar] [CrossRef]
- Yang, Q.; Diamond, M.P.; Al-Hendy, A. The emerging role of extracellular vesicle-derived miRNAs: Implication in cancer progression and stem cell related diseases. J. Clin. Epigenet. 2016, 2, 13. [Google Scholar] [PubMed]
- Chen, Y.; Yang, L.; Cui, T.; Pacyna-Gengelbach, M.; Petersen, I. HOPX is methylated and exerts tumour-suppressive function through Ras-induced senescence in human lung cancer. J. Pathol. 2015, 235, 397–407. [Google Scholar] [CrossRef]
- Chen, Y.; Pacyna-Gengelbach, M.; Deutschmann, N.; Niesporek, S.; Petersen, I. Homeobox gene HOP has a potential tumor suppressive activity in human lung cancer. Int. J. Cancer 2007, 121, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Schröder, D.C.; Nenkov, M.; Rizwan, M.N.; Abubrig, M.; Sonnemann, J.; Murrieta-Coxca, J.M.; Morales-Prieto, D.M.; Westermann, M.; Gaßler, N.; et al. Epithelial Membrane Protein 2 Suppresses Non-Small Cell Lung Cancer Cell Growth by Inhibition of MAPK Pathway. Int. J. Mol. Sci. 2021, 22, 2944. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.H.; Smith, D.J.; West, W.W.; Hollingsworth, M.A. Loss of fibulin-2 expression is associated with breast cancer progression. Am. J. Pathol. 2007, 170, 1535–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decapmaker, Z. The Role of Fibulin-2 in ECM-Driven Resistance to Targeted Therapy of Head and Neck Squamous Cell Carcinoma. Master’s Thesis, Ghent University, Ghent, Belgium, 2018. [Google Scholar]
- Lakshminarasimhan, R.; Liang, G. The Role of DNA Methylation in Cancer. Adv. Exp. Med. Biol. 2016, 945, 151–172. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Laird, P.W. Cancer epigenetics comes of age. Nat. Genet. 1999, 21, 163–167. [Google Scholar] [CrossRef]
- Dunwell, T.L.; Hesson, L.B.; Pavlova, T.; Zabarovska, V.; Kashuba, V.; Catchpoole, D.; Chiaramonte, R.; Brini, A.T.; Griffiths, M.; Maher, E.R.; et al. Epigenetic analysis of childhood acute lymphoblastic leukemia. Epigenetics 2009, 4, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Guo, J.; Wang, S.; Zhu, L.; Shen, Z. Identification of novel methylated targets in colorectal cancer by microarray analysis and construction of co-expression network. Oncol. Lett. 2017, 14, 2643–2648. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Myers, J.S.; Wang, F.; Wang, K.; Lucas, J.; Rosfjord, E.; Lucas, J.; Hooper, A.T.; Yang, S.; Lemon, L.A.; et al. Comparison of the molecular and cellular phenotypes of common mouse syngeneic models with human tumors. BMC Genom. 2020, 21, 2. [Google Scholar] [CrossRef] [Green Version]
- de Jong, M.; Maina, T. Of mice and humans: Are they the same?—Implications in cancer translational research. J. Nucl. Med. 2010, 51, 501–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Hui, D.; Fu, X. Roles of Fibulin-2 in Carcinogenesis. Med. Sci. Monit. 2020, 26, e918099. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, Y.; Sun, Y.B.; Wang, H.Y.; Sun, C.B.; Zhang, X. Role of fibulin-3 in lung cancer: In vivo and in vitro analyses. Oncol. Rep. 2014, 31, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Qi, C.; Liu, X.; Li, C.; Chen, J.; Shi, M. Fibulin-3 knockdown inhibits cervical cancer cell growth and metastasis in vitro and in vivo. Sci. Rep. 2018, 8, 10594. [Google Scholar] [CrossRef]
- Tan, H.; Zhang, J.; Fu, D.; Zhu, Y. Loss of fibulin-2 expression is involved in the inhibition of breast cancer invasion and forms a new barrier in addition to the basement membrane. Oncol. Lett. 2017, 14, 2663–2668. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, S.; Chen, J.; Liu, H.; Lu, J.; Jiang, H.; Huang, A.; Chen, Y. Fibulin-4 promotes osteosarcoma invasion and metastasis by inducing epithelial to mesenchymal transition via the PI3K/Akt/mTOR pathway. Int. J. Oncol. 2017, 50, 1513–1530. [Google Scholar] [CrossRef]
- Kinoshita, T.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Yamamoto, N.; Yoshino, H.; Itesako, T.; Enokida, H.; Nakagawa, M.; Okamoto, Y.; et al. Tumour-suppressive microRNA-29s inhibit cancer cell migration and invasion by targeting laminin-integrin signalling in head and neck squamous cell carcinoma. Br. J. Cancer 2013, 109, 2636–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshizuka, K.; Kikkawa, N.; Hanazawa, T.; Yamada, Y.; Okato, A.; Arai, T.; Katada, K.; Okamoto, Y.; Seki, N. Inhibition of integrin β1-mediated oncogenic signalling by the antitumor microRNA-29 family in head and neck squamous cell carcinoma. Oncotarget 2018, 9, 3663–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [Green Version]
- Schliekelman, M.J.; Gibbons, D.L.; Faca, V.M.; Creighton, C.J.; Rizvi, Z.H.; Zhang, Q.; Wong, C.H.; Wang, H.; Ungewiss, C.; Ahn, Y.H.; et al. Targets of the tumor suppressor miR-200 in regulation of the epithelial-mesenchymal transition in cancer. Cancer Res. 2011, 71, 7670–7682. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Yu, T.; Liu, D.; Shi, Y.; Yang, P.; Zhang, J.; Wang, J.; Liu, Y.; Zhang, X. Sepsis plasma-derived exosomal miR-1-3p induces endothelial cell dysfunction by targeting SERP1. Clin. Sci. 2021, 135, 347–365. [Google Scholar] [CrossRef]
- Liu, Q.; Peng, F.; Chen, J. The Role of Exosomal MicroRNAs in the Tumor Microenvironment of Breast Cancer. Int. J. Mol. Sci. 2019, 20, 3884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, P.; Almeida, F. Role of Exosomal miRNAs and the Tumor Microenvironment in Drug Resistance. Cells 2020, 9, 1450. [Google Scholar] [CrossRef] [PubMed]
- Nik Mohamed Kamal, N.N.S.B.; Shahidan, W.N.S. Non-Exosomal and Exosomal Circulatory MicroRNAs: Which Are More Valid as Biomarkers? Front. Pharmacol. 2019, 10, 1500. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Zhang, W.; Tan, J.; Ma, J.; Zhao, J. MiR-200b-3p Functions as an Oncogene by Targeting ABCA1 in Lung Adenocarcinoma. Technol. Cancer Res. Treat. 2019, 18, 1533033819892590. [Google Scholar] [CrossRef]
- Li, G.; Wu, F.; Yang, H.; Deng, X.; Yuan, Y. MiR-9-5p promotes cell growth and metastasis in non-small cell lung cancer through the repression of TGFBR2. Biomed. Pharmacother. 2017, 96, 1170–1178. [Google Scholar] [CrossRef]
- Wang, J.Y.; Zhang, Q.; Wang, D.D.; Yan, W.; Sha, H.H.; Zhao, J.H.; Yang, S.J.; Zhang, H.D.; Hou, J.C.; Xu, H.Z.; et al. MiR-29a: A potential therapeutic target and promising biomarker in tumors. Biosci. Rep. 2018, 38, BSR20171265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Wang, X.; Jing, L.; Li, Y. MiR-1-3p Inhibits Lung Adenocarcinoma Cell Tumorigenesis via Targeting Protein Regulator of Cytokinesis 1. Front. Oncol. 2019, 9, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, D.; Li, X.; Shao, Y.; He, Y.; Yu, H.; Ma, Z. MiR-199a-5p suppresses non-small cell lung cancer via targeting MAP3K11. J. Cancer 2019, 10, 2472–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Jin, K.; Xie, X.; Wang, K.; Zhang, H. MicroRNA-19a-3p inhibits the cellular proliferation and invasion of non-small cell lung cancer by downregulating UBAP2L. Exp. Ther. Med. 2020, 20, 2252–2261. [Google Scholar] [CrossRef]
- Lin, J.; Chen, Y.; Liu, L.; Shen, A.; Zheng, W. MicroRNA-155-5p suppresses the migration and invasion of lung adenocarcinoma A549 cells by targeting Smad2. Oncol. Lett. 2018, 16, 2444–2452. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Sun, S.; Shi, J.; Cao, F.; Han, X.; Chen, Z. MicroRNA-125a-5p plays a role as a tumor suppressor in lung carcinoma cells by directly targeting STAT3. Tumour Biol. 2017, 39, 1010428317697579. [Google Scholar] [CrossRef] [Green Version]
- Schmidmaier, R.; Baumann, P. ANTI-ADHESION evolves to a promising therapeutic concept in oncology. Curr. Med. Chem. 2008, 15, 978–990. [Google Scholar] [CrossRef]
- Nandhu, M.S.; Behera, P.; Bhaskaran, V.; Longo, S.L.; Barrera-Arenas, L.M.; Sengupta, S.; Rodriguez-Gil, D.J.; Chiocca, E.A.; Viapiano, M.S. Development of a Function-Blocking Antibody Against Fibulin-3 as a Targeted Reagent for Glioblastoma. Clin. Cancer Res. 2018, 24, 821–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Chen, Y.; Li, Y.; Grün, K.; Berndt, A.; Zhou, Z.; Petersen, I. Cystatin A suppresses tumor cell growth through inhibiting epithelial to mesenchymal transition in human lung cancer. Oncotarget 2018, 9, 14084–14098. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hühn, D.; Knösel, T.; Pacyna-Gengelbach, M.; Deutschmann, N.; Petersen, I. Downregulation of connexin 26 in human lung cancer is related to promoter methylation. Int. J. Cancer 2005, 113, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fibulin 2 | |||||
|---|---|---|---|---|---|
| Negative | Positive | p-Value | |||
| Type | ADC | 28 | 24 | ||
| SCC | 14 | 28 | 0.047 | ||
| Methylation status | Unmethylation | 4 | 3 | Fisher’s | |
| Partial methylation and methylation | 15 | 15 | 1 | ||
| Gender | Male | 29 | 39 | ||
| Female | 14 | 14 | 0.51 | ||
| Age | ≤65 | 34 | 42 | ||
| >65 | 9 | 11 | 0.983 | ||
| pT | 1–2 | 42 | 45 | Fisher’s | |
| 3–4 | 2 | 10 | 0.061 | ||
| pN | 0–1 | 27 | 39 | ||
| 2–4 | 16 | 14 | 0.257 | ||
| pM | 0 | 40 | 49 | Fisher’s | |
| 1–3 | 3 | 4 | 1 | ||
| Grade | 1–2 | 20 | 28 | ||
| 3–4 | 23 | 25 | 0.538 | ||
| Lower T Stage | Fibulin 2 vs. Fibronectin | ||
|---|---|---|---|
| ADC + SCC | ADC | SCC | |
| Sample number | 49 | 23 | 26 |
| Correlation coefficient (r Value) | −0.204 | 0.016 | −0.391 |
| Sig. (2-tailed) p | 0.16 | 0.944 | 0.048 |
| Fibulin 2 vs. E-Cadherin | Fibulin 2 vs. Vimentin | Fibulin 2 vs. Cytokeratin | |||||||
|---|---|---|---|---|---|---|---|---|---|
| ADC + SCC | ADC | SCC | ADC + SCC | ADC | SCC | ADC + SCC | ADC | SCC | |
| Sample number | 81 | 45 | 36 | 79 | 44 | 35 | 78 | 44 | 34 |
| Correlation coefficient (r Value) | −0.031 | −0.206 | 0.358 | −0.105 | −0.056 | −0.207 | 0.144 | −0.047 | 0.422 |
| Sig. (2-tailed) p | 0.781 | 0.175 | 0.032 | 0.357 | 0.716 | 0.233 | 0.208 | 0.764 | 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Nenkov, M.; Schröder, D.C.; Abubrig, M.; Gassler, N.; Chen, Y. Fibulin 2 Is Hypermethylated and Suppresses Tumor Cell Proliferation through Inhibition of Cell Adhesion and Extracellular Matrix Genes in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 11834. https://doi.org/10.3390/ijms222111834
Ma Y, Nenkov M, Schröder DC, Abubrig M, Gassler N, Chen Y. Fibulin 2 Is Hypermethylated and Suppresses Tumor Cell Proliferation through Inhibition of Cell Adhesion and Extracellular Matrix Genes in Non-Small Cell Lung Cancer. International Journal of Molecular Sciences. 2021; 22(21):11834. https://doi.org/10.3390/ijms222111834
Chicago/Turabian StyleMa, Yunxia, Miljana Nenkov, Desiree Charlotte Schröder, Mohamed Abubrig, Nikolaus Gassler, and Yuan Chen. 2021. "Fibulin 2 Is Hypermethylated and Suppresses Tumor Cell Proliferation through Inhibition of Cell Adhesion and Extracellular Matrix Genes in Non-Small Cell Lung Cancer" International Journal of Molecular Sciences 22, no. 21: 11834. https://doi.org/10.3390/ijms222111834