
Plastidial Expression of 3β-Hydroxysteroid Dehydrogenase and Progesterone 5β-Reductase Genes Confer Enhanced Salt Tolerance in Tobacco

 , , , , and
, , , , and 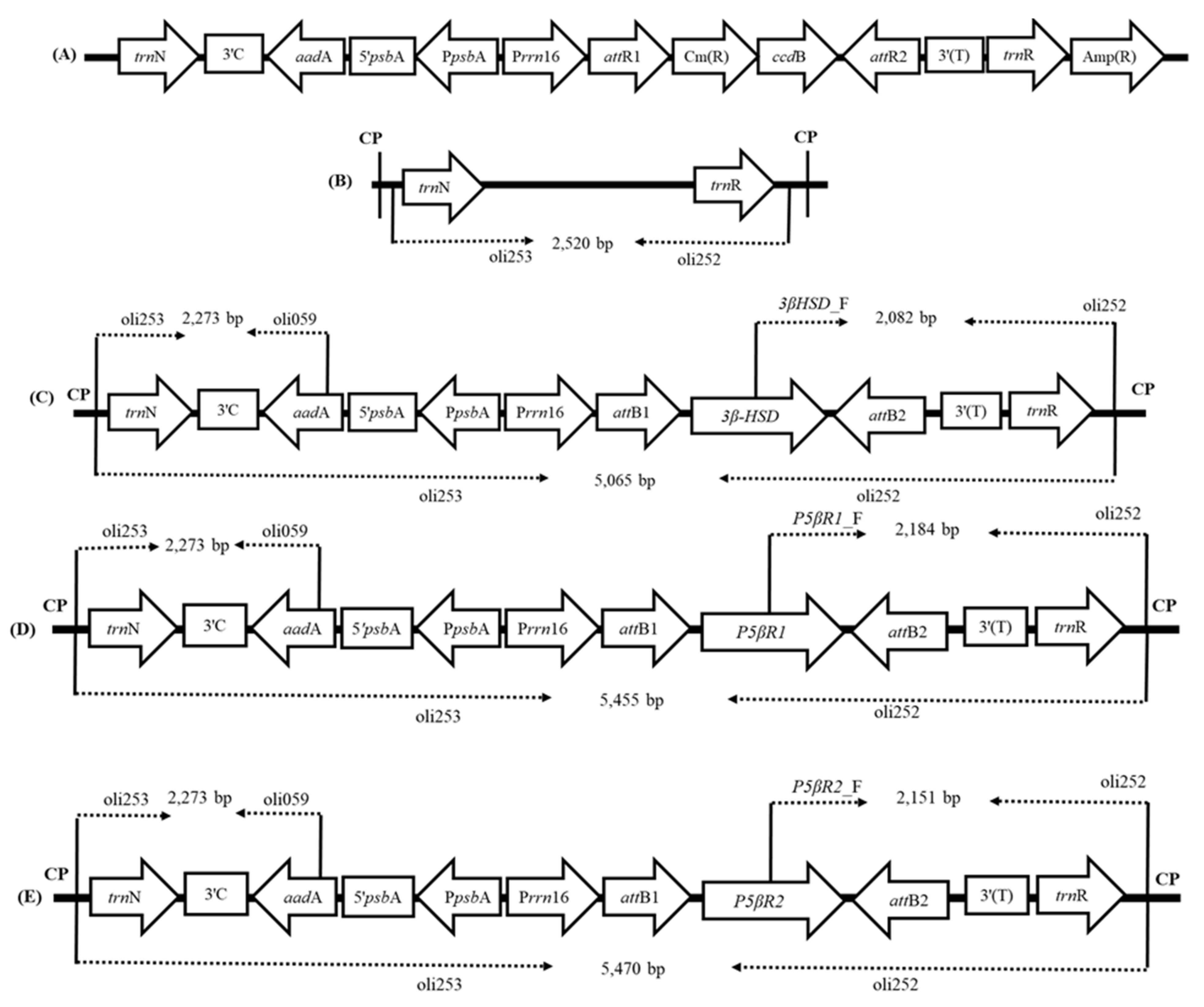{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation of Transplastomic Plants Expressing 3β-HSD, P5βr1 and P5βr2 Genes
2.1.1. Plastid Transformation Vectors and Development of Transplastomic Plants
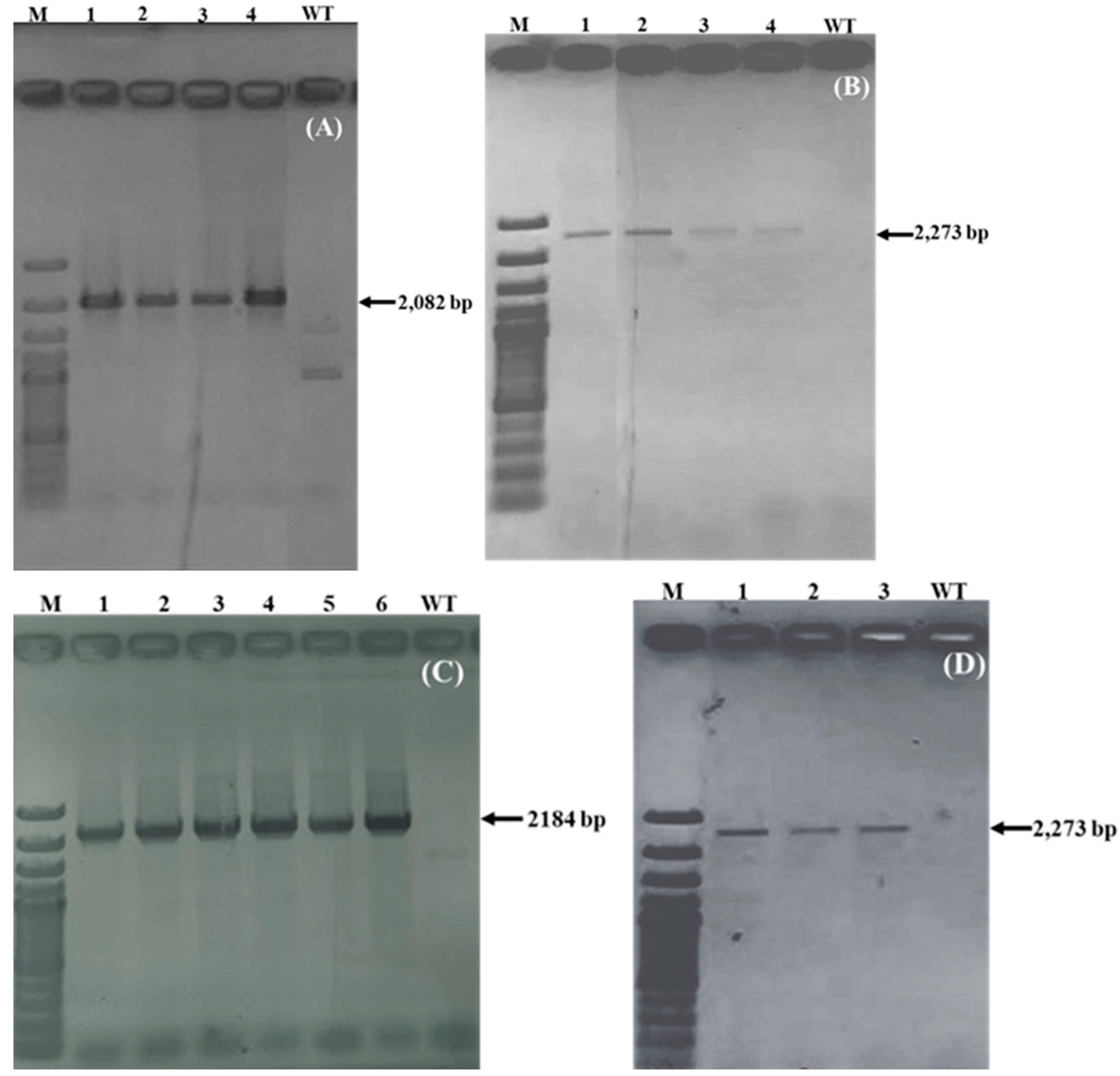
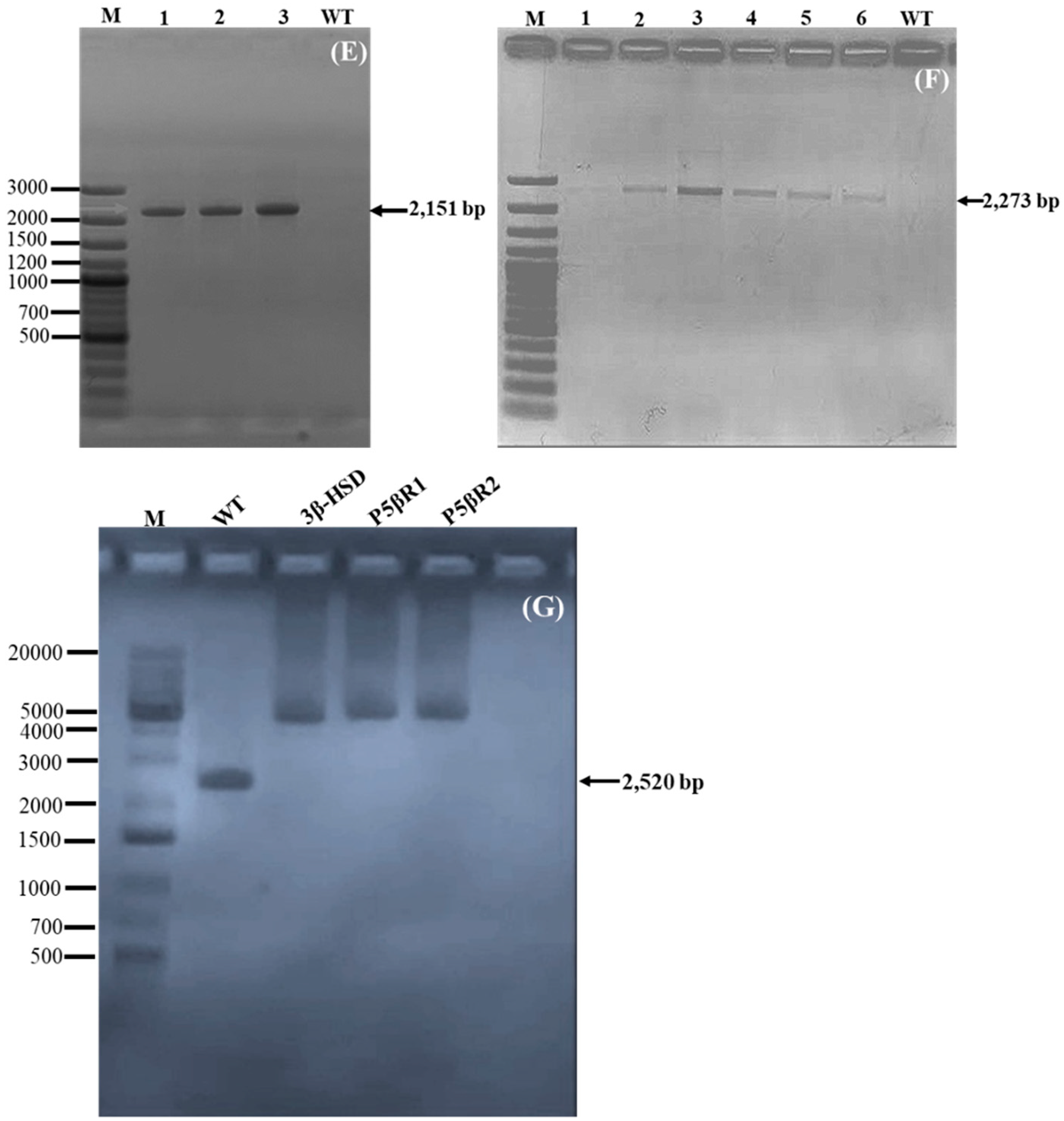
2.1.2. Confirmation of Transgene Integration
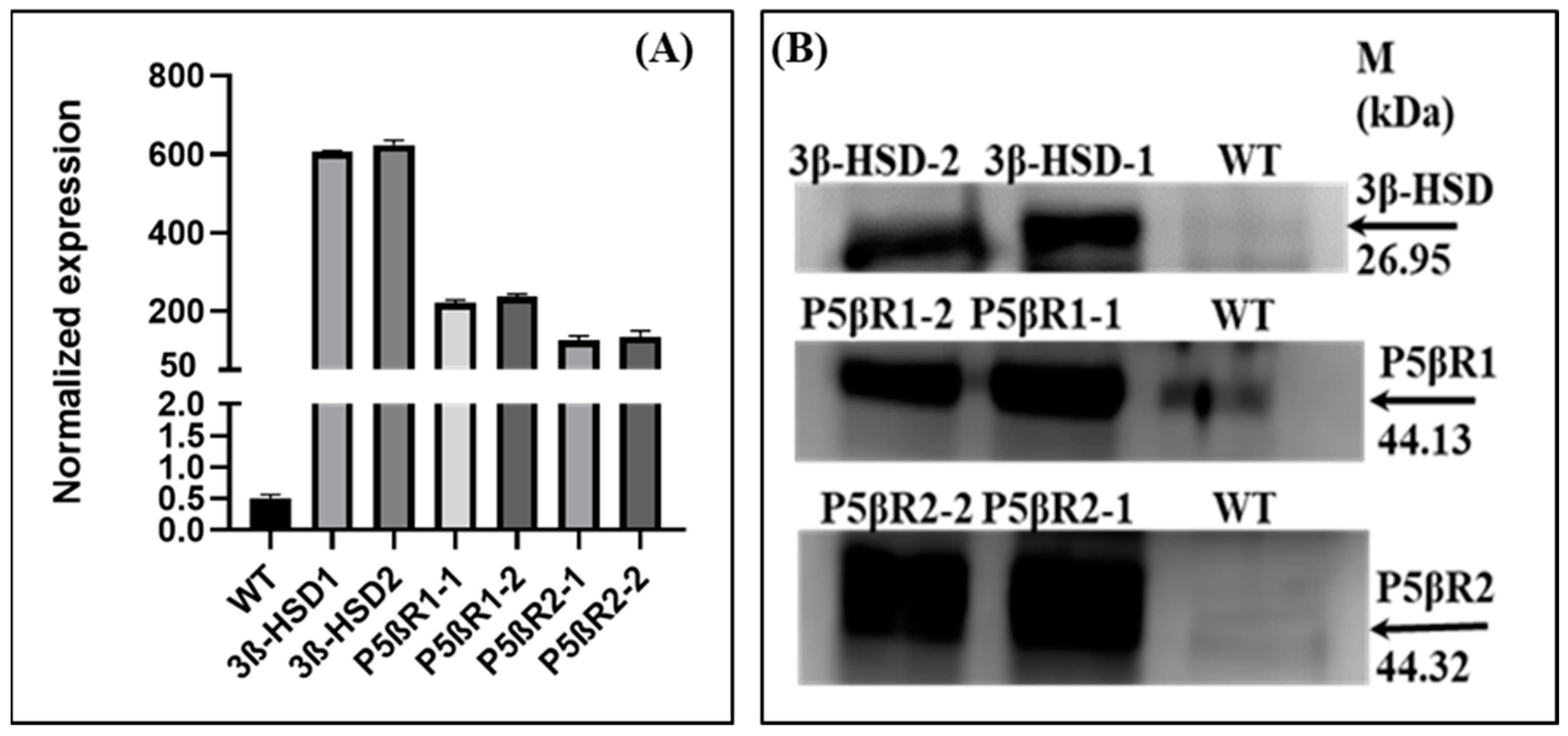
2.1.3. Expression of Transgene RT-qRT-PCR
2.1.4. End-to-End PCR
2.2. Western Blot

2.3. Subcellular Localization of 3β-HSD, P5βR1 and P5βR2
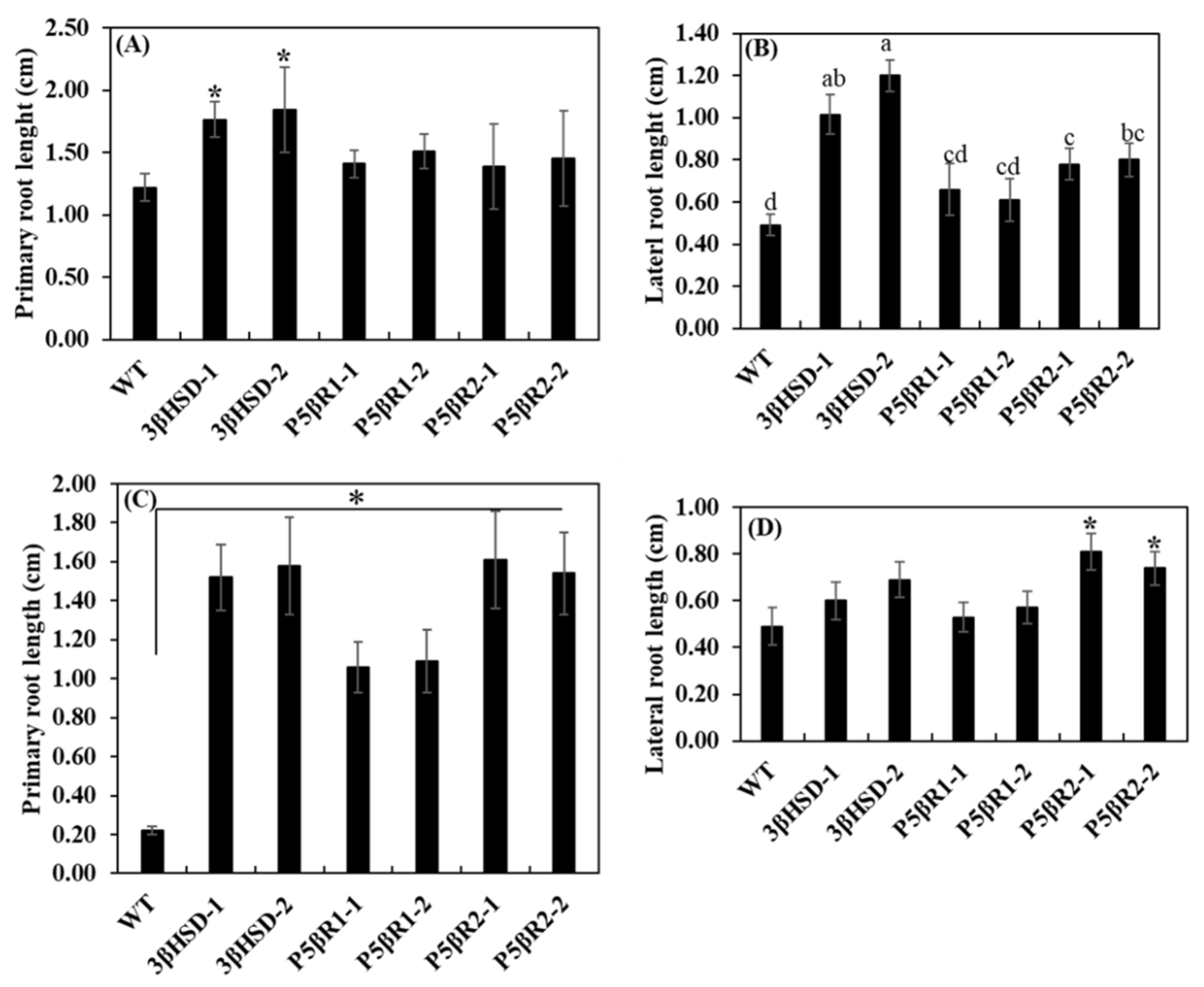
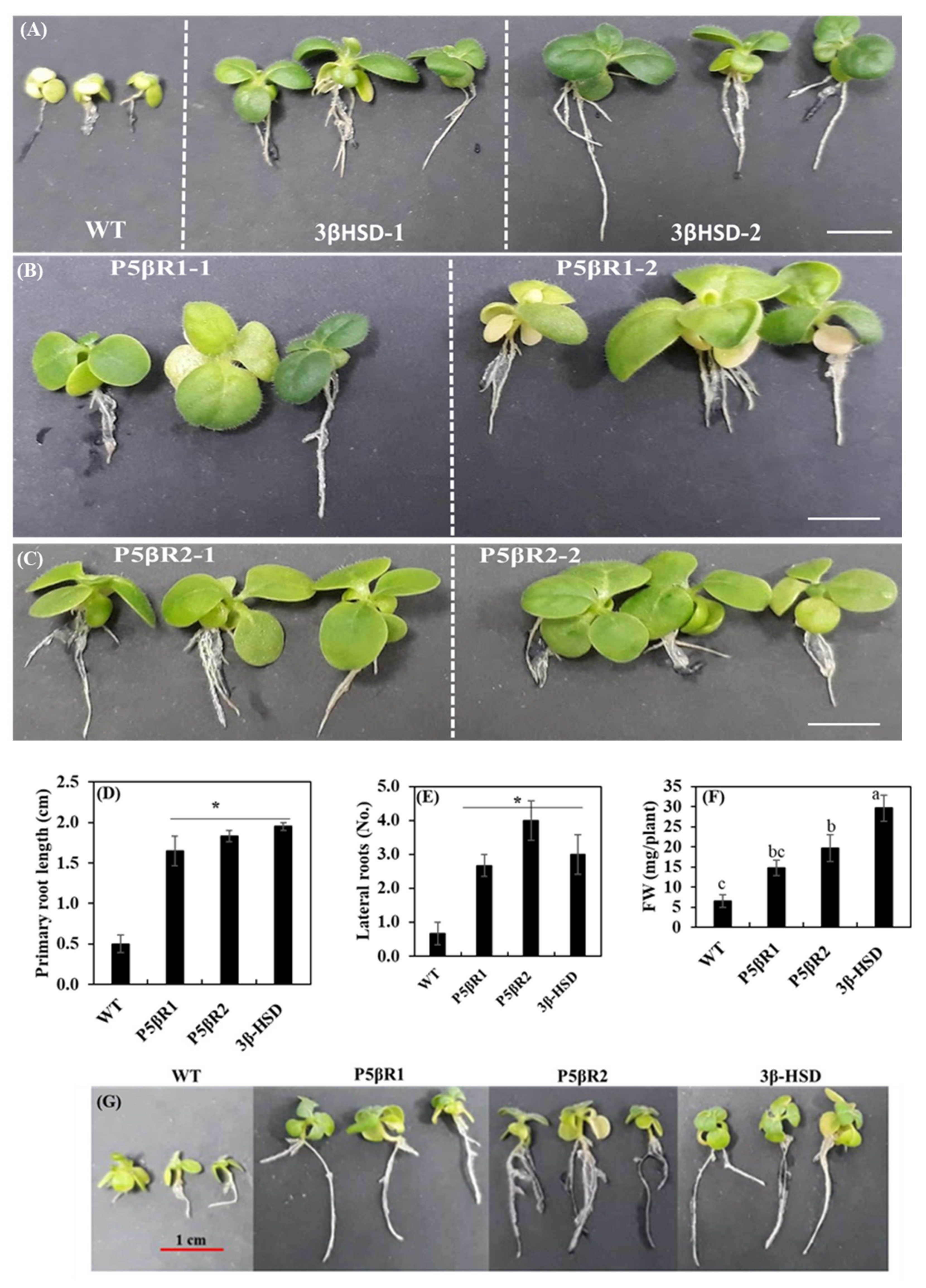
2.4. Effect of Salinity on Plant Growth and Development
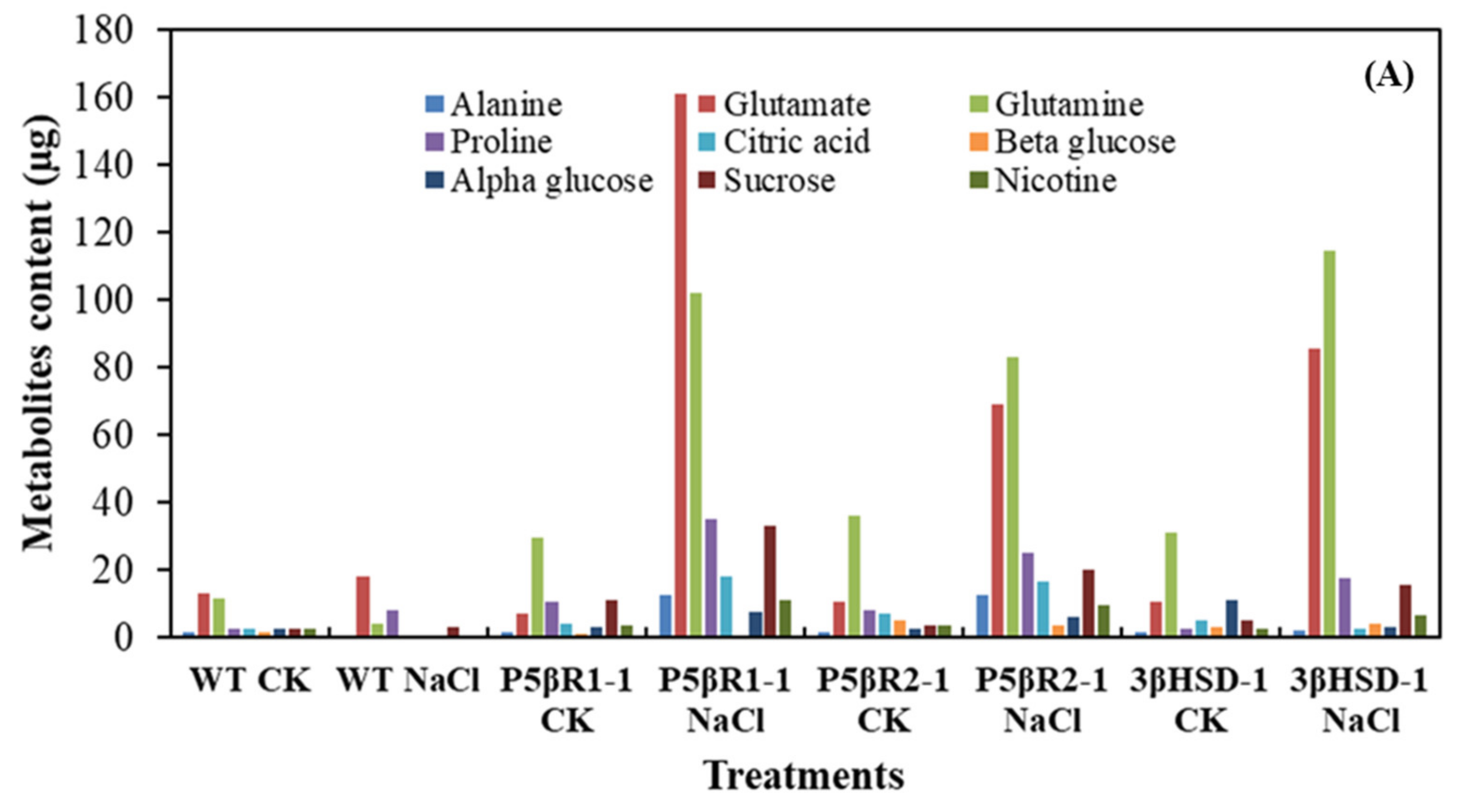
2.5. Metabolite Analysis of the Plants Grown under Salt Stress
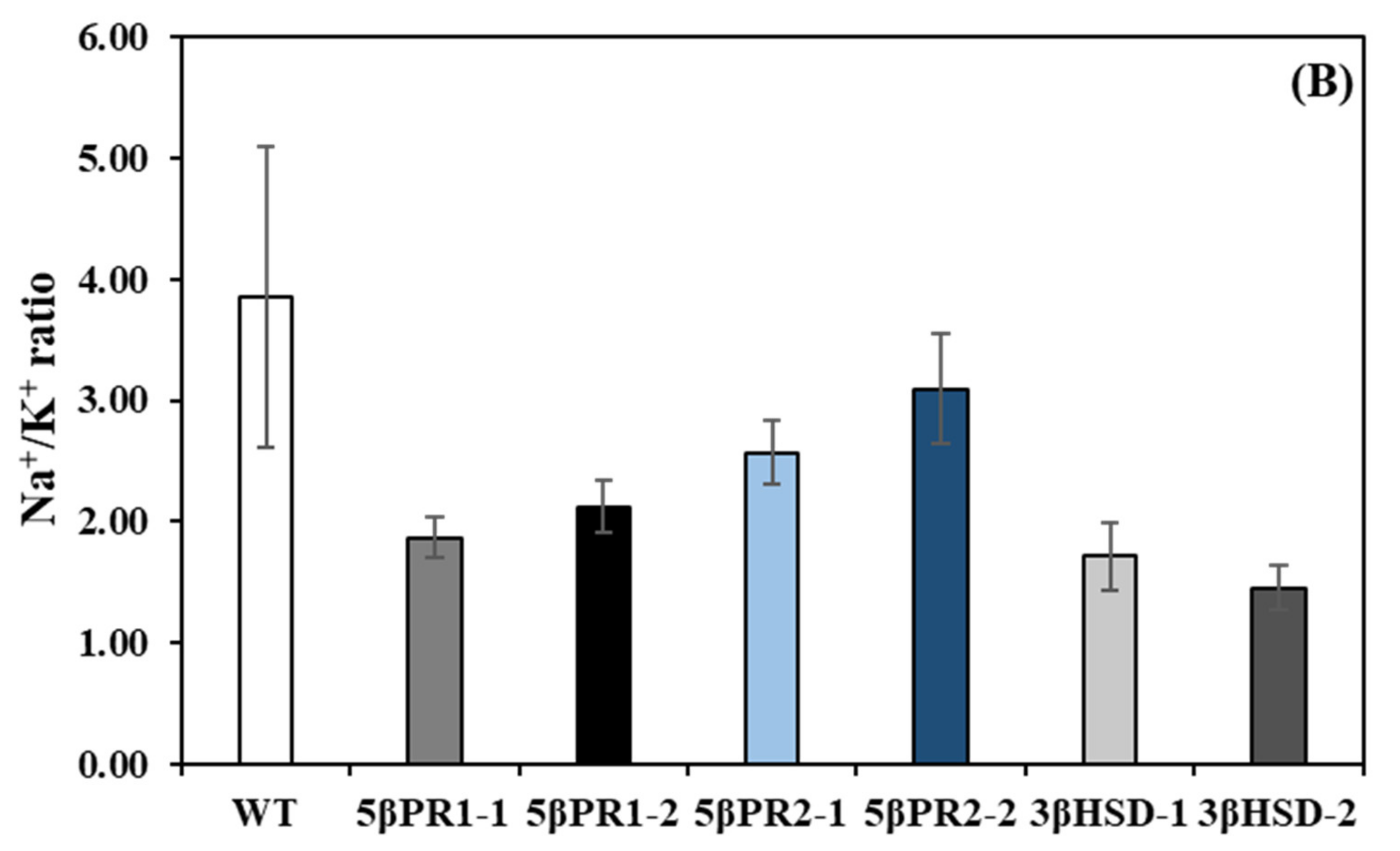
2.6. Transplastomic Expression of the P5βr1, P5βr2 and 3β-HSD Decreases Na+/K+ Ratio under Salt Stress
3. Discussion
3.1. Contribution of 3β-HSD, P5βR1 and P5βR2 Genes towards Plant Growth and Development
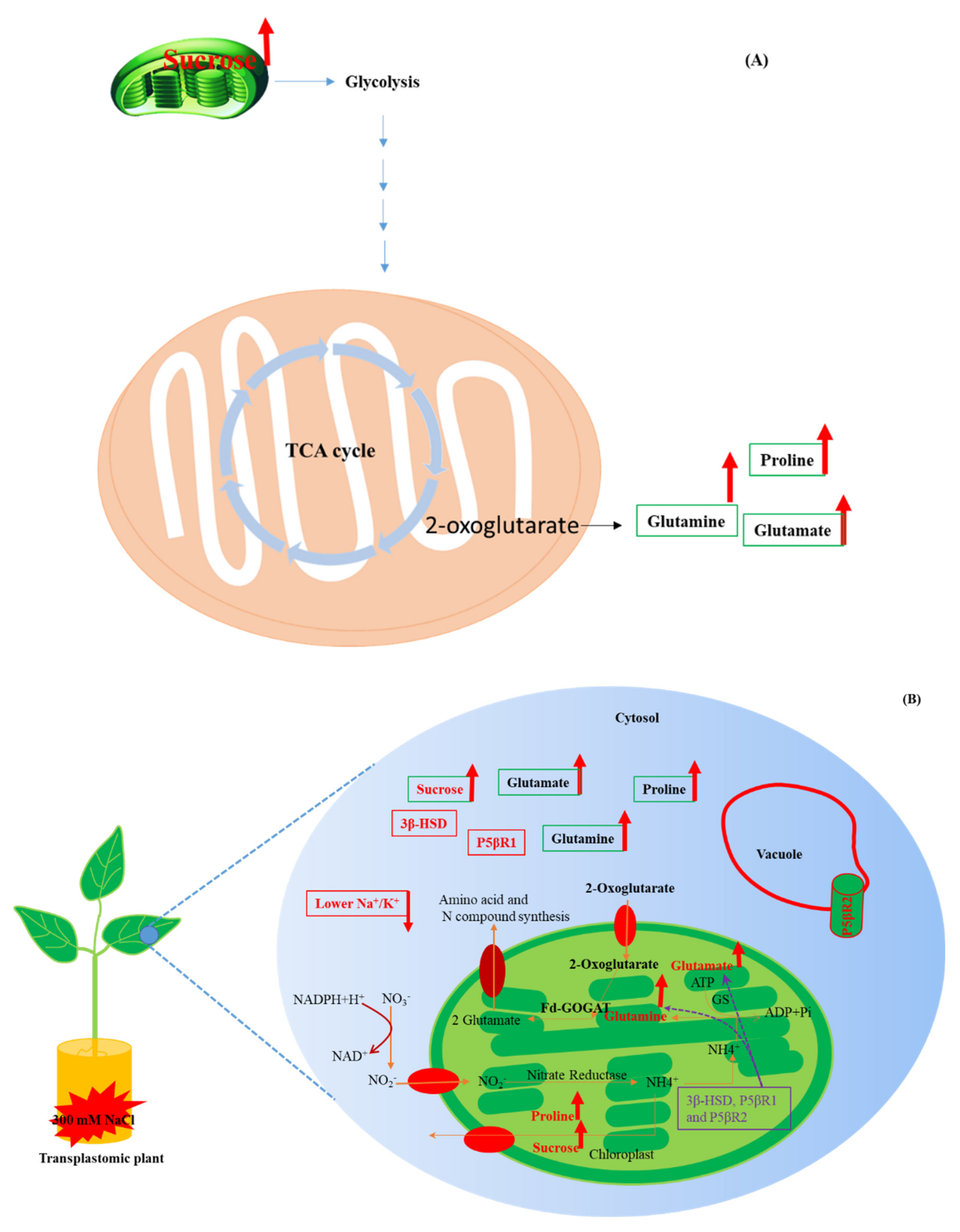
3.2. Enhanced Biosynthesis of Glutamate, Glutamine, Proline and Sucrose in Transplastomic Plants under Salt Stress
3.3. 3β-HSD, P5βR1 and P5βR2 Confer Salt Tolerance
3.4. Protective Role of Proline, Sucrose, Glutamate and Glutamine under Salt Stress
4. Materials and Methods
4.1. Vector Construction
4.2. Plastid Transformation of Tobacco and Regeneration of Transformed Plants
4.3. Confirmation of Transformation and Transgene Expression
4.4. Confirmation of Transgene Expression by Real Time Qrt-PCR
4.5. End-to-End PCR
4.6. Western Blot
4.7. Cloning of the 3βhsd, P5βr1 and P5βr2 and Vector Construction for Subcellular Localization
4.8. Salt Tolerance Assessment and NMR-Based Metabolites Determination
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertol, J.; Rigotto, C.; de Pádua, R.M.; Kreis, W.; Barardi, C.R.M.; Braga, F.; Simões, C.M.O. Antiherpes activity of glucoevatromonoside, a cardenolide isolated from a Brazilian cultivar of Digitalis lanata. Antivir. Res. 2011, 92, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreis, W.; Haug, B.; Yucesan, B. Somaclonal variation of cardenolide content in Heywood’s foxglove, a source for the antiviral cardenolide glucoevatromonoside, regenerated from permanent shoot culture and callus. Vitr. Cell. Dev. Biol. Anim. 2014, 51, 35–41. [Google Scholar] [CrossRef]
- Pérez-Bermúdez, P.; Moya García, A.A.; Tuñón, I.; Gavidia, I. Digitalis purpurea P5βR2, encoding steroid 5β-reductase, is a novel defense-related gene involved in cardenolide biosynthesis. N. Phytol. 2010, 185, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Kreis, W. The Foxgloves (Digitalis) Revisited. Planta Med. 2017, 83, 962–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, M.; de Padua, R.M.; Herl, V.; Muller-Uri, F.; Kreis, W. Expression of 3beta-HSD and P5betaR, genes respectively coding for Delta5-3beta-hydroxysteroid dehydrogenase and progesterone 5beta-reductase, in leaves and cell cultures of Digitalis lanata EHRH. Planta Med. 2010, 76, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Herl, V.; Fischer, G.; Müller-Uri, F.; Kreis, W. Molecular cloning and heterologous expression of progesterone 5β-reductase from Digitalis lanata Ehrh. Phytochemistry 2006, 67, 225–231. [Google Scholar] [CrossRef]
- Herl, V.; Frankenstein, J.; Meitinger, N.; Muller-Uri, F.; Kreis, W. Delta 5-3beta-hydroxysteroid dehydrogenase (3 beta HSD) from Digitalis lanata. Heterologous expression and characterisation of the recombinant enzyme. Planta Med. 2007, 73, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Cacho, M.; Morán, M.; Tárrago, J.F.; Corchete, P. Calcium restriction induces cardenolide accumulation in cell suspension cultures of Digitalis thapsi L. Plant Cell Rep. 1995, 14, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Cingoz, G.S.; Gurel, E. Effects of salicylic acid on thermotolerance and cardenolide accumulation under high temperature stress in Digitalis trojana Ivanina. Plant Physiol. Biochem. 2016, 105, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Paranhos, A.; Fernández-Tárrago, J.; Corchete, P. Relationship between active oxygen species and cardenolide production in cell cultures of Digitalis thapsi: Effect of calcium restriction. N. Phytol. 1999, 141, 51–60. [Google Scholar] [CrossRef]
- Finsterbusch, A.; Lindemann, P.; Grimm, R.; Eckerskorn, C.; Luckner, M. Delta(5)-3beta-hydroxysteroid dehydrogenase from Digitalis lanata Ehrh.—A multifunctional enzyme in steroid metabolism? Planta 1999, 209, 478–486. [Google Scholar] [CrossRef]
- Saito, K.; Yamazaki, M.; Shimomura, K.; Yoshimatsu, K.; Murakoshi, I. Genetic transformation of foxglove (Digitalis purpurea) by chimeric foreign genes and production of cardioactive glycosides. Plant Cell Rep. 1990, 9, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yamazaki, M.; Kaneko, H.; Murakoshi, I.; Fukuda, Y.; Van Montagu, M. Tissue-specific and stress-enhancing expression of the TR promoter for mannopine synthase in transgenic medicinal plants. Planta 1991, 184, 40–46. [Google Scholar] [CrossRef]
- Lehmann, U.; Moldenhauer, D.; Thomar, S.; Diettrich, B.; Luckner, M. Regeneration of plants from Digitalis lanata cells transformed with Agrobacterium tumefaciens carrying bacterial genes encoding neomycin phosphotransferase II and β-glucuronidase. J. Plant Physiol. 1995, 147, 53–57. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Z.; Piao, C.; Lu, K.; Wang, Z.; Cui, M.-L. A stable and efficient Agrobacterium tumefaciens-mediated genetic transformation of the medicinal plant Digitalis purpurea L. Appl. Biochem. Biotechnol. 2014, 172, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alonso, N.; Chong-Pérez, B.; Capote, A.; Pérez, A.; Izquierdo, Y.; Angenon, G.; Jiménez, E. Agrobacterium tumefaciens-mediated genetic transformation of Digitalis purpurea L. Plant Biotechnol. Rep. 2014, 8, 387–397. [Google Scholar] [CrossRef]
- Pradel, H.; Dumke-Lehmann, U.; Diettrich, B.; Luckner, M. Hairy root cultures of Digitalis lanata. Secondary metabolism and plant regeneration. J. Plant Physiol. 1997, 151, 209–215. [Google Scholar] [CrossRef]
- Sales, E.; Segura, J.; Arrillaga, I. Agrobacterium tumefaciens-mediated genetic transformation of the cardenolide-producing plant Digitalis minor L. Planta Med. 2003, 69, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Sales, E.; Munoz-Bertomeu, J.; Arrillaga, I.; Segura, J. Enhancement of cardenolide and phytosterol levels by expression of an N-terminally truncated 3-hydroxy-3-methylglutaryl CoA reductase in Transgenic digitalis minor. Planta Med. 2007, 73, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.P.; Lindemann, P. Expression of recombinant Digitalis lanata EHRH. cardenolide 16′-O-glucohydrolase in Cucumis sativus L. hairy roots. Plant Cell Rep. 2006, 25, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Hagimori, M.; Matsumoto, T.; Obi, Y. Studies on the Production of Digitalis Cardenolides by Plant Tissue Culture: II. Effect of light and plant growth substances on digitoxin formation by undifferentiated cells and shoot-forming cultures of digitalis purpurea l. Grown in liquid media. Plant Physiol. 1982, 69, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Waheed, M.T.; Ismail, H.; Gottschamel, J.; Mirza, B.; Lössl, A.G. Plastids: The green frontiers for vaccine production. Front.plant sci. 2015, 6, 1005. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H. Molecular strategies for gene containment in transgenic crops. Nat. Biotechnol. 2002, 20, 581–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagemann, R. The Sexual Inheritance of Plant Organelles. In Molecular Biology Biotechnol. Plant Organelles: Chloroplasts and Mitochondria; Daniell, H., Chase, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 93–113. [Google Scholar]
- Munns, R. Genes and salt tolerance: Bringing them together. N. Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Gurel, E.; Karvar, S.; Yucesan, B.; Eker, I.; Sameeullah, M. An overview of cardenolides in digitalis-more than a cardiotonic compound. Curr. Pharm. Des. 2017, 23, 5104–5114. [Google Scholar] [CrossRef] [PubMed]
- Svab, Z.; Hajdukiewicz, P.; Maliga, P. Stable transformation of plastids in higher plants. Proc. Natl. Acad. Sci. USA 1990, 87, 8526–8530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangol, S.; Singh, R.; Chen, Y.; Jwa, N.-S. Visualization of multicolored in vivo organelle markers for co-localization studies in Oryza sativa. Mol. Cells 2017, 40, 828. [Google Scholar]
- Klein, J.; Horn, E.; Ernst, M.; Leykauf, T.; Leupold, T.; Dorfner, M.; Wolf, L.; Ignatova, A.; Kreis, W.; Munkert, J. RNAi-mediated gene knockdown of progesterone 5β-reductases in Digitalis lanata reduces 5β-cardenolide content. Plant Cell Rep. 2021, 40, 1631–1646. [Google Scholar] [CrossRef]
- Malcolm, S.B.; Zalucki, M.P. Milkweed latex and cardenolide induction may resolve the lethal plant defence paradox. Èntomol. Exp. Appl. 1996, 80, 193–196. [Google Scholar] [CrossRef]
- Pumirat, P.; Boonyuen, U.; Vanaporn, M.; Pinweha, P.; Tandhavanant, S.; Korbsrisate, S.; Chantratita, N. The role of short-chain dehydrogenase/oxidoreductase, induced by salt stress, on host interaction of B. pseudomallei. BMC Microbiol. 2014, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Sun, Q.; Wu, J.; Zhao, P.; Sun, Y.; Guo, Z. Genome-Wide Identification and Characterization of Short-Chain Dehydrogenase/Reductase (SDR) Gene Family in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 9498. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Chen, A.; Xiao, L.; Muller, H.M.; Ache, P.; Haberer, G.; Zhang, M.; Jia, W.; Deng, P.; Huang, R.; et al. A high-quality genome assembly of quinoa provides insights into the molecular basis of salt bladder-based salinity tolerance and the exceptional nutritional value. Cell Res. 2017, 27, 1327–1340. [Google Scholar] [CrossRef]
- Eker, İ.; Yücesan, B.; Sameeullah, M.; Welβ, W.; Müller-Uri, F.; Gürel, E.; Kreis, W. Phylogeny of Anatolian (Turkey) species in the Digitalis sect. Globiflorae (Plantaginaceae). Phytotaxa 2016, 244, 263–282. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 2013, 65, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S. Regulation of Potassium Transport in Leaves: From Molecular to Tissue Level. Ann. Bot. 2003, 92, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.H.; Endo, A.; Zhou, L.; Penney, J.; Chen, H.C.; Arroyo, A.; Leon, P.; Nambara, E.; Asami, T.; Seo, M.; et al. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 2002, 14, 2723–2743. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Asami, T.; Wu, X.; Tsang, E.W.T.; Cutler, A.J. A Putative Hydroxysteroid Dehydrogenase Involved in Regulating Plant Growth and Development. Plant Physiol. 2007, 145, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, C.; Xu, K.; Cui, J.; Zhang, Z.A.; Ling, F.; An, J.; Wu, Z. Effects of salt stress on photosynthetic characteristics and some physiological traits of rice varieties at different nitrogen levels. J. South China Agric. Univ. 2015, 36, 6–12. [Google Scholar]
- Nasraoui, H.A.; Bouthour, D.; Hfaidh, R.; Gouia, H.; Pageau, K.; Chaffei, H.C. The role of nitrogen availability for the salt-tolerance of two different varieties of durum wheat. Bull. Environ. Contam. Toxicol. 2013, 91, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, Z.; Zhou, Y.; Han, J.; Shi, D. Effects of salt stress on ion balance and nitrogen metabolism in rice. Plant Soil Environ. 2012, 58, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Uddin, M.; Ismail, M.R.; Ashrafuzzaman, M. Responses of glutamine synthetase-glutamate synthase cycle enzymes in tomato leaves under salinity stress. Int. J. Agricul. Biol. 2012, 14, 509–515. [Google Scholar]
- Meng, S.; Su, L.; Li, Y.; Wang, Y.; Zhang, C.; Zhao, Z. Nitrate and Ammonium Contribute to the Distinct Nitrogen Metabolism of Populus simonii during Moderate Salt Stress. PLoS ONE 2016, 11, e0150354. [Google Scholar] [CrossRef]
- Zhonghua, T.; Yanju, L.; Xiaorui, G.; Yuangang, Z. The combined effects of salinity and nitrogen forms on Catharanthus roseus: The role of internal ammonium and free amino acids during salt stress. J. Plant Nutr. Soil Sci. 2011, 174, 135–144. [Google Scholar] [CrossRef]
- Hessini, K.; Hamed, K.B.; Gandour, M.; Mejri, M.; Abdelly, C.; Cruz, C. Ammonium nutrition in the halophyte Spartina alterniflora under salt stress: Evidence for a priming effect of ammonium? Plant Soil 2013, 370, 163–173. [Google Scholar] [CrossRef]
- De Souza Miranda, R.; Gomes-Filho, E.; Prisco, J.T.; Alvarez-Pizarro, J.C. Ammonium improves tolerance to salinity stress in Sorghum bicolor plants. Plant Growth Regul. 2016, 78, 121–131. [Google Scholar] [CrossRef]
- Ding, F.; Wang, R.; Chen, B. Effect of exogenous ammonium gluconate on growth, ion flux and antioxidant enzymes of maize (Zea Mays, L.) seedlings under NaCl stress. Plant Biol. 2019, 21, 643–651. [Google Scholar] [CrossRef]
- Bezerra, M.A.F.; Pereira, W.E.; Bezerra, F.T.C.; Cavalcante, L.F.; da Silva Medeiros, S.A. Nitrogen as a mitigator of salt stress in yellow passion fruit seedlingss. Semina Ciências Agrárias 2019, 40, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.M.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S.; Arif, M.S.; Kausar, R. Nitrogen nutrition and adaptation of glycophytes to saline environment: A review. Arch. Agron. Soil Sci. 2018, 64, 1181–1206. [Google Scholar] [CrossRef]
- Stitt, M.; Krapp, A. The interaction between elevated carbon dioxide and nitrogen nutrition: The physiological and molecular background. Plant Cell Environ. 1999, 22, 583–621. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.M.; Habash, D.Z. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. N. Phytol. 2009, 182, 608–620. [Google Scholar] [CrossRef]
- Renau-Morata, B.; Molina, R.V.; Carrillo, L.; Cebolla-Cornejo, J.; Sánchez-Perales, M.; Pollmann, S.; Domínguez-Figueroa, J.; Corrales, A.R.; Flexas, J.; Vicente-Carbajosa, J.; et al. Ectopic Expression of CDF3 Genes in Tomato Enhances Biomass Production and Yield under Salinity Stress Conditions. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berteli, F.; Corrales, E.; Guerrero, C.; Ariza, M.J.; Pliego, F.; Valpuesta, V. Salt stress increases ferredoxin-dependent glutamate synthase activity and protein level in the leaves of tomato. Physiol. Plant. 1995, 93, 259–264. [Google Scholar] [CrossRef]
- Debouba, M.; Gouia, H.; Suzuki, A.; Ghorbel, M.H. NaCl stress effects on enzymes involved in nitrogen assimilation pathway in tomato “Lycopersicon esculentum” seedlings. J. Plant Physiol. 2006, 163, 1247–1258. [Google Scholar] [CrossRef]
- Saito, T.; Matsukura, C.; Sugiyama, M.; Watahiki, A.; Ohshima, I.; Iijima, Y.; Konishi, C.; Fujii, T.; Inai, S.; Fukuda, N. Screening for γ-aminobutyric acid (GABA)-rich tomato varieties. J. Jpn. Soc. Hortic. Sci. 2008, 77, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Flores, P.; Botella, M.Á.; Cerdá, A.; Martínez, V. Influence of nitrate level on nitrate assimilation in tomato (Lycopersicon esculentum) plants under saline stress. Can. J. Bot. 2004, 82, 207–213. [Google Scholar] [CrossRef]
- Debouba, M.; Maâroufi-Dghimi, H.; Suzuki, A.; Ghorbel, M.H.; Gouia, H. Changes in growth and activity of enzymes involved in nitrate reduction and ammonium assimilation in tomato seedlings in response to NaCl stress. Ann. Bot. 2007, 99, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Parvanova, D.; Ivanov, S.; Konstantinova, T.; Karanov, E.; Atanassov, A.; Tsvetkov, T.; Alexieva, V.; Djilianov, D. Transgenic tobacco plants accumulating osmolytes show reduced oxidative damage under freezing stress. Plant Physiol. Biochem. 2004, 42, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Verma, D.P.S.; Zhang, C. Regulation of proline and arginine biosynthesis in plants. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.K., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 249–265. [Google Scholar]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl. S1), S165–S183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Jiang, Z.; Song, G.; Shan, X.; Wei, Z.; Liu, Y.; Jiang, C.; Jiang, Y.; Jin, F.; Li, Y. Association Analysis and Identification of ZmHKT1;5 Variation With Salt-Stress Tolerance. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Luo, Q.; Wang, Q.; Zhang, X.; Qi, Z.; Xu, F.; Lei, X.; Cao, Y.; Chow, W.S.; Sun, G. Physiological and proteomic responses to salt stress in chloroplasts of diploid and tetraploid black locust (Robinia pseudoacacia L.). Sci. Rep. 2016, 6, 23098. [Google Scholar] [CrossRef] [Green Version]
- Hong, Z.; Lakkineni, K.; Zhang, Z.; Verma, D.P.S. Removal of Feedback Inhibition of Δ1-Pyrroline-5-Carboxylate Synthetase Results in Increased Proline Accumulation and Protection of Plants from Osmotic Stress1. Plant Physiol. 2000, 122, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhu, J.K. Proline Accumulation and Salt-Stress-Induced Gene Expression in a Salt-Hypersensitive Mutant of Arabidopsis. Plant Physiol. 1997, 114, 591–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]
- Ali, Q.; Habib-ur-Rehman Athar, M.Z.; Haider, S.S.; Aslam, N.; Shehzad, F.; Naseem, J.; Ashraf, R.; Ali, A.; Hussain, S.M. Role of Amino Acids in Improving Abiotic Stress Tolerance to Plants. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 175–204. [Google Scholar]
- Slocum, R.; Weinstein, L. Osmotic stress-induced putrescine accumulation as a mechanism of ammonia detoxification in oat leaves. Plant Physiol. Supp. 1990, 93, 24. [Google Scholar]
- El-Bassiouny, H.M.; Bekheta, M. Effect of salt stress on relative water content, lipid peroxidation, polyamines, amino acids and ethylene of two wheat cultivars. Int. J. Agricul. Biol. 2005, 7, 363–368. [Google Scholar]
- Gottschamel, J.; Waheed, M.T.; Clarke, J.L.; Lossl, A.G. A novel chloroplast transformation vector compatible with the Gateway((R)) recombination cloning technology. Transgenic Res. 2013, 22, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Hajdukiewicz, P.T.; Allison, L.A.; Maliga, P. The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J. 1997, 16, 4041–4048. [Google Scholar] [CrossRef] [Green Version]
- Svab, Z.; Maliga, P. High-frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proc. Natl. Acad. Sci. USA 1993, 90, 913–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waheed, M.T.; Thones, N.; Muller, M.; Hassan, S.W.; Gottschamel, J.; Lossl, E.; Kaul, H.P.; Lossl, A.G. Plastid expression of a double-pentameric vaccine candidate containing human papillomavirus-16 L1 antigen fused with LTB as adjuvant: Transplastomic plants show pleiotropic phenotypes. Plant Biotechnol. J. 2011, 9, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Sameeullah, M.; Sasaki, T.; Yamamoto, Y. Sucrose transporter NtSUT1 confers aluminum tolerance on cultured cells of tobacco (Nicotiana tabacum L.). Soil Sci. Plant Nutr. 2013, 59, 756–770. [Google Scholar] [CrossRef]
- Fuentes, P.; Zhou, F.; Erban, A.; Karcher, D.; Kopka, J.; Bock, R. A new synthetic biology approach allows transfer of an entire metabolic pathway from a medicinal plant to a biomass crop. eLife 2016, 5, e13664. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; Kannangara, R.; Jørgensen, K.; Fuglsang, A.T. Analysis of peptide PSY1 responding transcripts in the two Arabidopsis plant lines: Wild type and psy1r receptor mutant. BMC Genom. 2014, 15, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales-Mendoza, S.; Monreal-Escalante, E.; González-Ortega, O.; Hernández, M.; Fragoso, G.; Garate, T.; Sciutto, E. Transplastomic plants yield a multicomponent vaccine against cysticercosis. J. Biotechnol. 2018, 266, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Latif, S.; Gottschamel, J.; Syed, T.; Younus, I.; Gull, K.; Sameeullah, M.; Batool, N.; Lössl, A.G.; Mariz, F.; Müller, M. Inducible expression of human papillomavirus-16 L1 capsomeres in the plastomes of Nicotiana tabacum: Transplastomic plants develop normal flowers and pollen. Biotechnol. Appl. Biochem. 2021. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakagawa, T.; Kurose, T.; Hino, T.; Tanaka, K.; Kawamukai, M.; Niwa, Y.; Toyooka, K.; Matsuoka, K.; Jinbo, T.; Kimura, T. Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 2007, 104, 34–41. [Google Scholar] [CrossRef]
- Xu, K.; Huang, X.; Wu, M.; Wang, Y.; Chang, Y.; Liu, K.; Zhang, J.; Zhang, Y.; Zhang, F.; Yi, L. A rapid, highly efficient and economical method of Agrobacterium-mediated in planta transient transformation in living onion epidermis. PLoS ONE 2014, 9, e83556. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Bayramoğlu Karşı, M.B.; Yenisoy-Karakaş, S.; Karakaş, D. Investigation of washout and rainout processes in sequential rain samples. Atmos. Environ. 2018, 190, 53–64. [Google Scholar] [CrossRef]
- Ziaf, K.; Loukehaich, R.; Gong, P.; Liu, H.; Han, Q.; Wang, T.; Li, H.; Ye, Z. A Multiple Stress-Responsive Gene ERD15 from Solanum pennellii Confers Stress Tolerance in Tobacco. Plant Cell Physiol. 2011, 52, 1055–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sameeullah, M.; Yildirim, M.; Aslam, N.; Baloğlu, M.C.; Yucesan, B.; Lössl, A.G.; Saba, K.; Waheed, M.T.; Gurel, E. Plastidial Expression of 3β-Hydroxysteroid Dehydrogenase and Progesterone 5β-Reductase Genes Confer Enhanced Salt Tolerance in Tobacco. Int. J. Mol. Sci. 2021, 22, 11736. https://doi.org/10.3390/ijms222111736
Sameeullah M, Yildirim M, Aslam N, Baloğlu MC, Yucesan B, Lössl AG, Saba K, Waheed MT, Gurel E. Plastidial Expression of 3β-Hydroxysteroid Dehydrogenase and Progesterone 5β-Reductase Genes Confer Enhanced Salt Tolerance in Tobacco. International Journal of Molecular Sciences. 2021; 22(21):11736. https://doi.org/10.3390/ijms222111736
Chicago/Turabian StyleSameeullah, Muhammad, Muhammet Yildirim, Noreen Aslam, Mehmet Cengiz Baloğlu, Buhara Yucesan, Andreas G. Lössl, Kiran Saba, Mohammad Tahir Waheed, and Ekrem Gurel. 2021. "Plastidial Expression of 3β-Hydroxysteroid Dehydrogenase and Progesterone 5β-Reductase Genes Confer Enhanced Salt Tolerance in Tobacco" International Journal of Molecular Sciences 22, no. 21: 11736. https://doi.org/10.3390/ijms222111736