
Neuroprotective Effect of 1,4-Naphthoquinones in an In Vitro Model of Paraquat and 6-OHDA-Induced Neurotoxicity

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Preparation of 1,4-NQ Derivatives
2.2. Determination of the Cytoprotective Properties of 1,4-NQs in An In vitro PD Model Induced by PQ and 6-OHDA
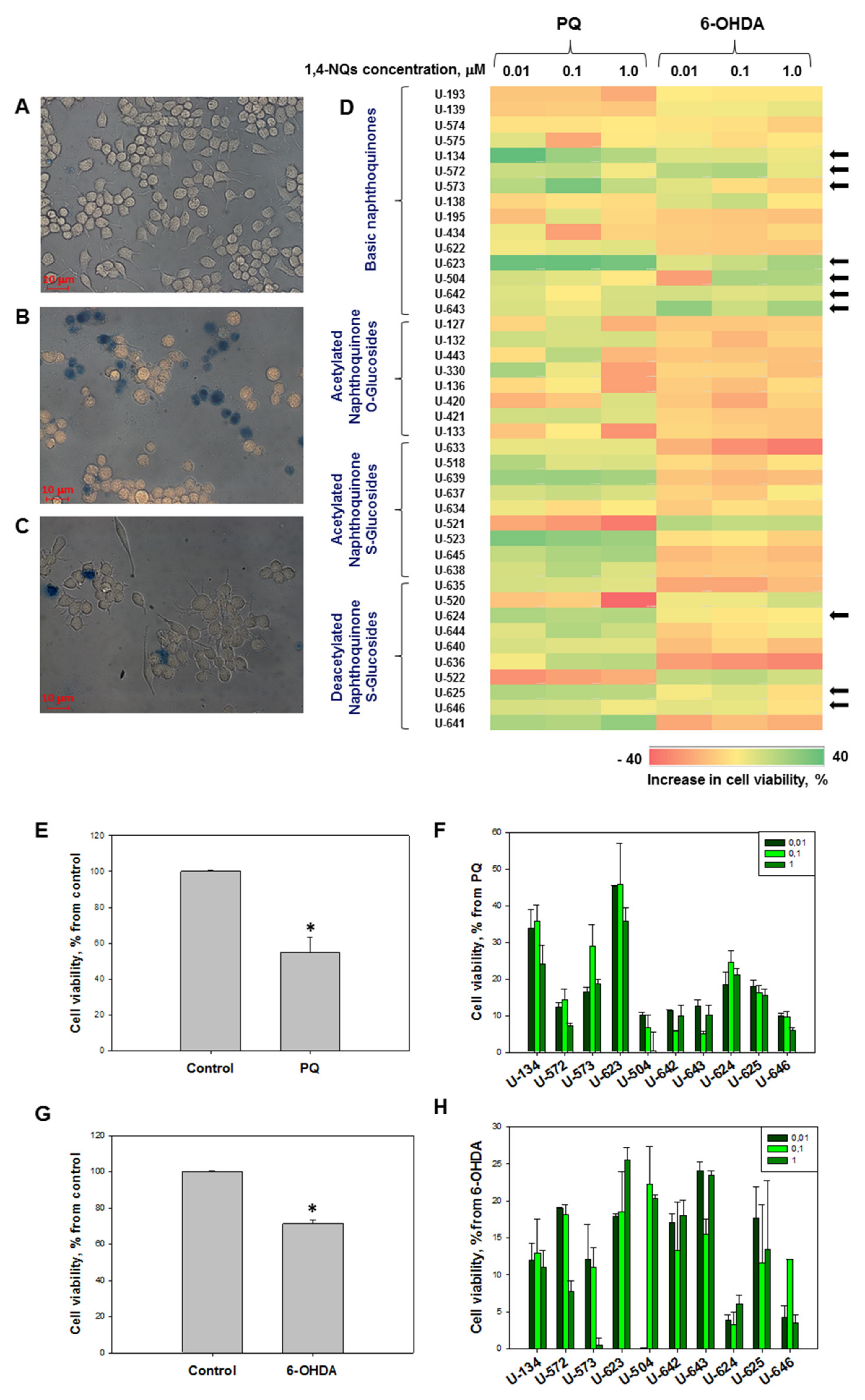
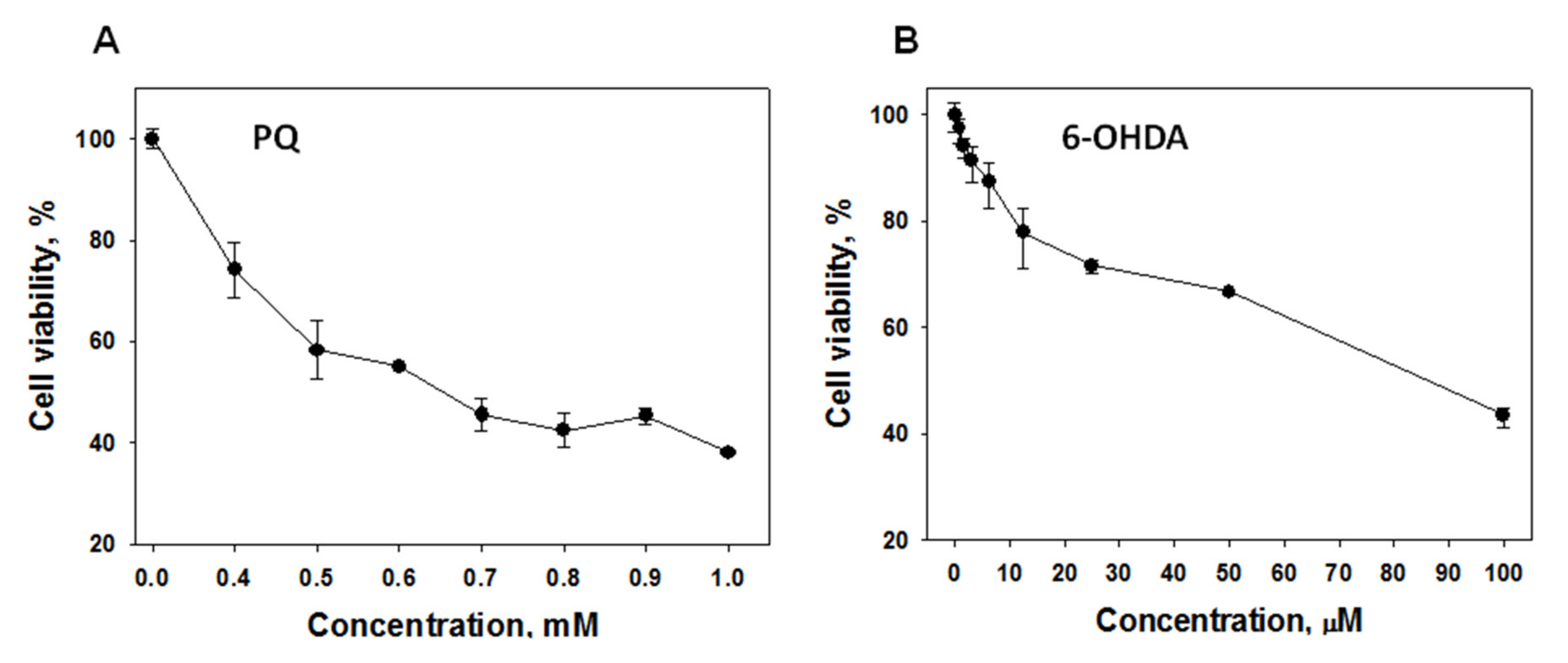
2.2.1. Effect of 1,4-NQs on Cell Viability Using MTT Assay
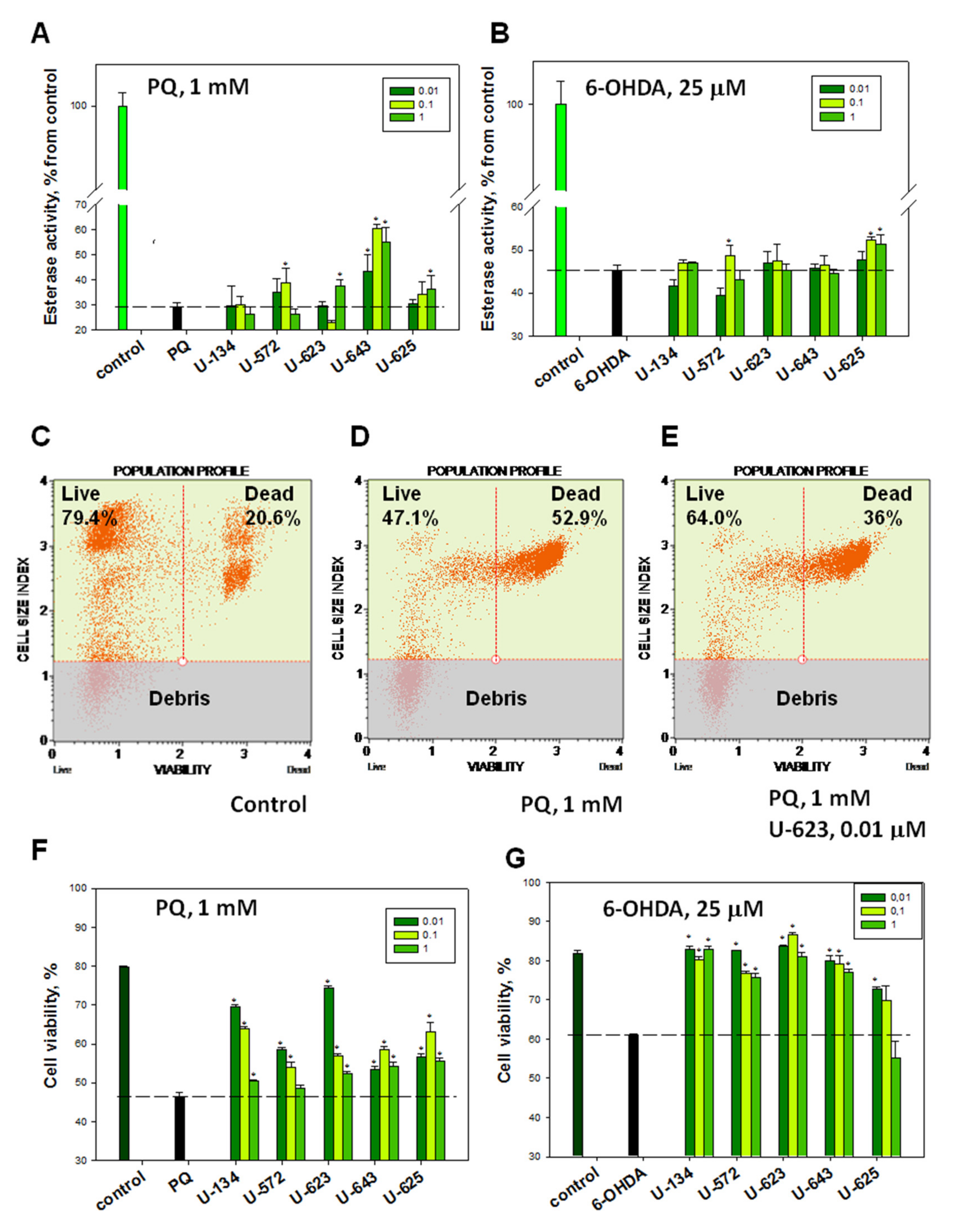
2.2.2. Effects of 1,4-NQs on Cell Viability in FDA and PI Tests
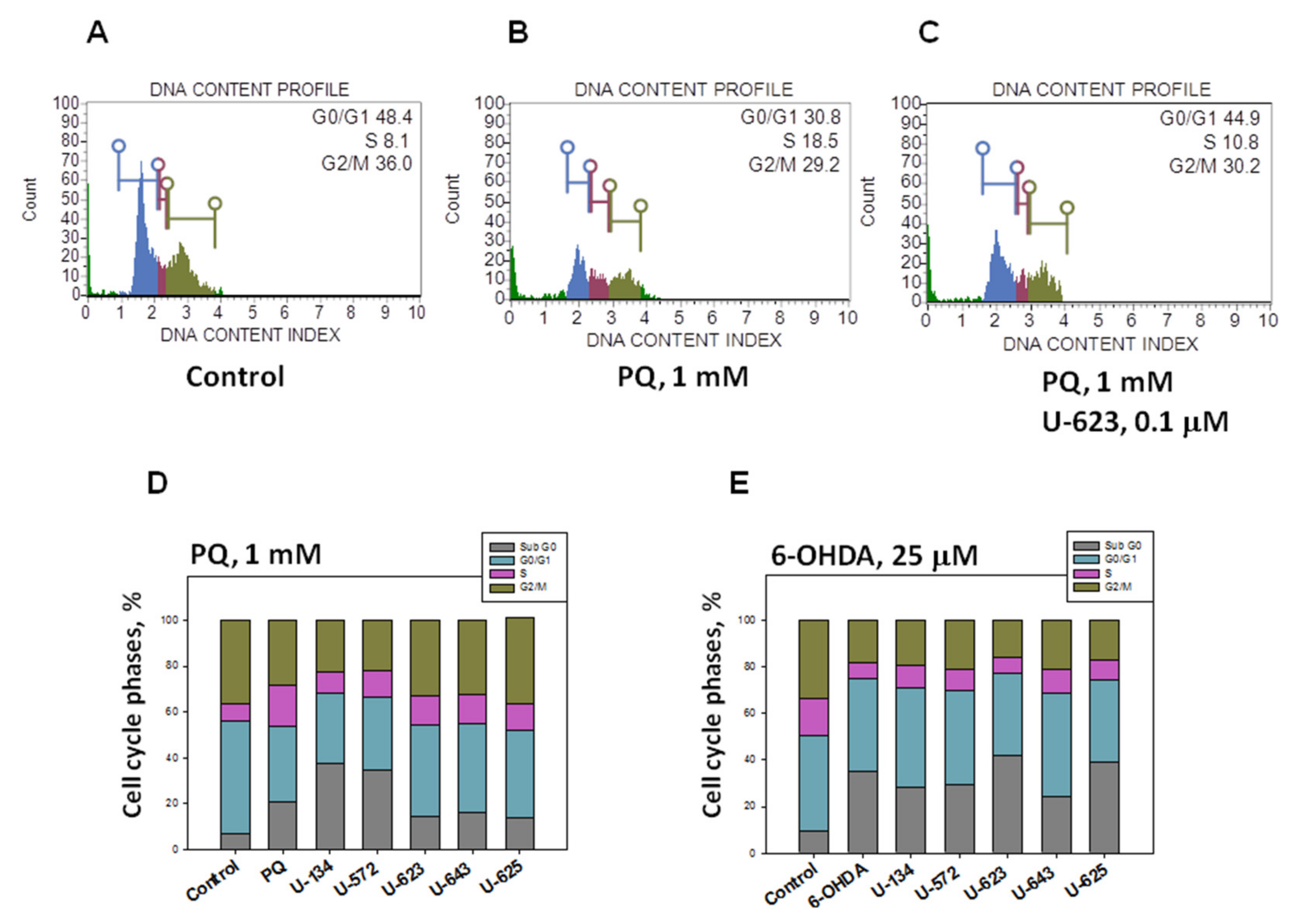
2.3. Influence of 1,4-NQs on Neuro-2a Cell Cycle Disturbance Caused by PQ and 6-OHDA
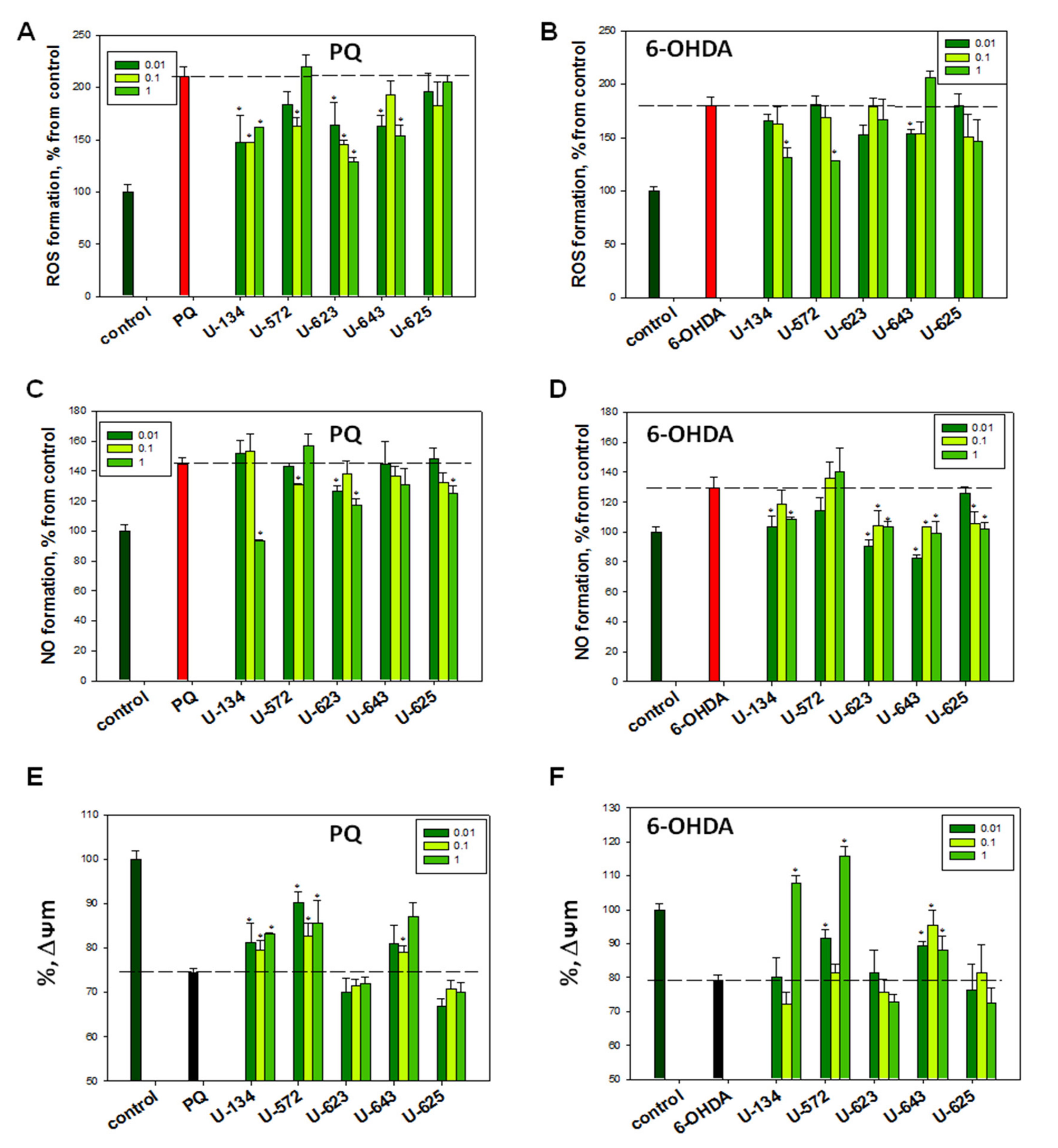
2.4. Evaluation of ROS and NO Production Levels in Neuro-2a Cells Caused by PQ and 6-OHDA in the Presence of 1,4-NQs
2.5. Influence of 1,4-NQs on Mitochondrial Membrane Potential Altered by PQ and 6-OHDA
2.6. Effects of 1,4-NQs on Scavenging DPPH Radical
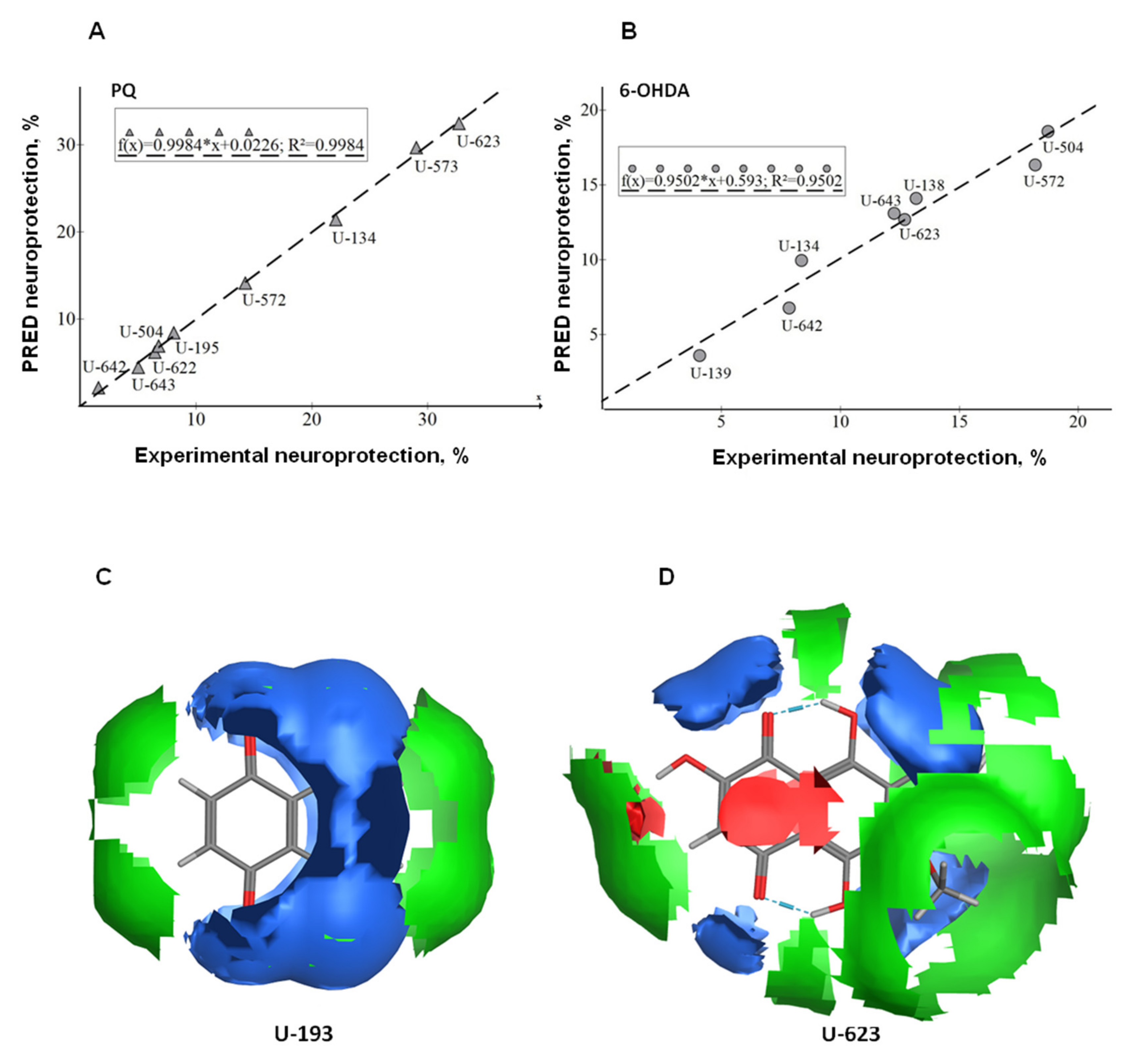
2.7. Generation and Validation of QSAR Models
| PQ = | 6-OHDA = |
| 68.61008 | 44.40310 |
| +2.63075 * SlogP_VSA3 | +0.48187 * SMR_VSA7 |
| +2.63075 * SMR_VSA2 | −0.31397 * SlogP_VSA0 |
| −4.17081 * TPSA | −0.33998 * SlogP_VSA5 |
| −2.29788 * SMR_VSA3 | −0.34822 * SlogP_VSA9 |
| +4.32894 * SlogP_VSA0 | +0.31683 * TPSA |
| +6.35996 * SlogP_VSA6 | −0.10669 * vsurf_D1 |
| +0.82448 * SlogP_VSA5 |
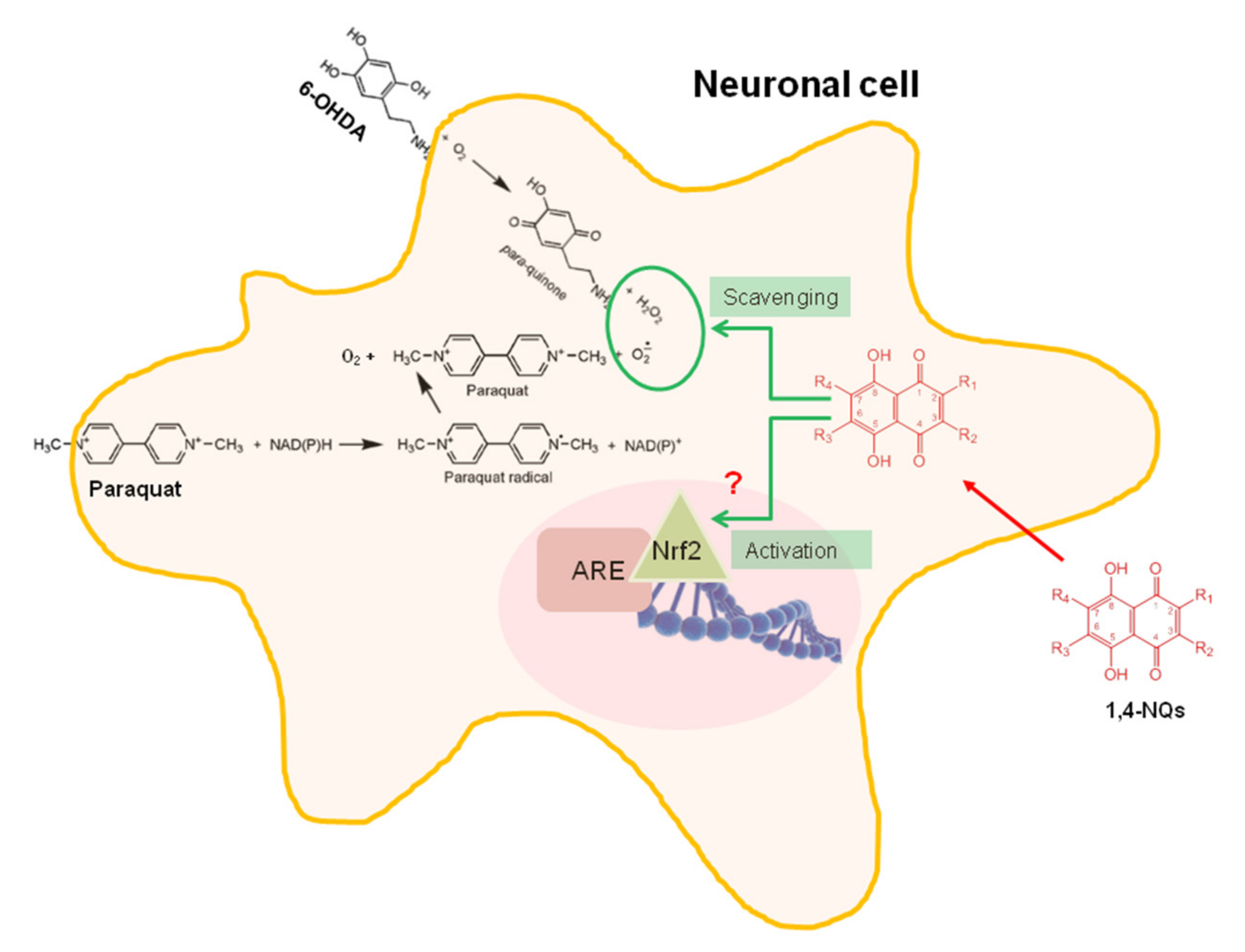
3. Discussion
4. Material and Methods
4.1. Synthesis of 1,4-Naphtoquinones
4.2. Cell Line and Culture Condition
4.3. Paraquat and 6-OHDA Induced In Vitro Model of Neurotoxicity
4.4. Cell Viability Assay
4.4.1. MTT Test
4.4.2. FDA Staining
4.4.3. PI Staining
4.5. Cell Cycle Analysis
4.6. Mitochondrial Membrane Potential (MMP) Detection
4.7. ROS and NO Level Analysis
4.8. Radical Scavenging Assay
4.9. Computer Modeling and Quantitative Structure-Activity Relationship (QSAR) Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1:4-NQs | 1,4-Naphthoquinones |
| 6-OHDA | 6-Hydroxydopamine |
| ARE | Antioxidant responsive element |
| DAF-FM | 4-Amino-5-Methylamino-2′,7′-Difluorofluorescein Diacetate |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethylsulfoxide |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| FDA | Fluorescein diacetate |
| H2DCF-DA | 2,7-Dichlorodihydrofluorescein diacetate |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MAO | Monoamine oxidase |
| MMP | Mitochondrial membrane potential |
| MPTP | 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NO | Nitric oxide |
| Nrf2 | Nuclear factor E2-related factor 2 |
| PBS | Phosphate-buffered saline |
| PD | Parkinson’s disease |
| PI | Propidium iodide |
| PQ | Paraquat or N,N′-dimethyl-4,4′-dipyridylium dichloride |
| QSAR | Quantitative structure-activity relationship |
| ROS | Reactive oxygen species |
| SDS | Sodium dodecyl sulfate |
| TMRM | Tetramethylrhodamine methyl ester perchlorate |
Appendix A

References
- Pringsheim, T.; Jette, N.; Frolkis, A.; Steeves, T.D. The prevalence of Parkinson’s disease: A systematic review and meta-analysis. Mov. Disord. 2014, 29, 1583–1590. [Google Scholar] [CrossRef]
- de Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Bove, J.; Prou, D.; Perier, C.; Przedborski, S. Toxin-induced models of Parkinson’s disease. NeuroRX 2005, 2, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Aminin, D.; Polonik, S. 1,4-Naphthoquinones: Some biological properties and application. Chem. Pharm. Bull. 2020, 68, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.; Zhang, B.-D.; Mohapatra, S.; Li, G.; Li, Y.-M.; Gazit, E.; Segal, D. Novel mannitol-based small molecules for inhibiting aggregation of a-synucleinamyloids in Parkinson’s disease. Front. Mol. Biosci. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnoli, K.; Petzer, J.B.; Steyn, S.J.; Van Der Schyf, C.J. Inhibition of human MAO-A and MAO-B by a compound isolated from flue-cured tobacco leaves and its neuroprotective properties in the MPTP mouse model of neurodegeneration. Inflammopharmacology 2003, 11, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Sari, Y.; Khalil, A. Monoamine oxidase inhibitors extracted from Tobacco Smoke as neuroprotective factors for potential treatment of Parkinson’s disease. CNS. Neurol. Disord. Drug. Targets. 2015, 14, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Mostert, S.; Petzer, A.; Petzer, J.P. The evaluation of 1,4-benzoquinones as inhibitors of human monoamine oxidase. Eur. J. Med. Chem. 2017, 135, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Mostert, S.; Petzer, A.; Petzer, J.P. Evaluation of natural and synthetic 1,4-naphthoquinones as inhibitors of monoamine oxidase. Chem. Biol. Drug Des. 2016, 87, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Vial1, M.L.; Gee1, L.; Davis, R.A.; St. John, J.A.; Ekberg, J.A.K. The plant natural product 2-methoxy-1,4-naphthoquinone stimulates therapeutic neural repair properties of olfactory ensheathing cells. Sci. Rep. 2020, 10, 951. [Google Scholar] [CrossRef] [PubMed]
- Ekimova, I.V.; Plaksina, D.V.; Pastukhov, Y.F.; Lapshina, K.V.; Lazarev, V.F.; Mikhaylova, E.R.; Polonik, S.G.; Pani, B.; Margulis, B.A.; Guzhova, I.V.; et al. New HSF1 inducer as a therapeutic agent in a rodent model of Parkinson’s disease. Exp. Neurol. 2018, 306, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Pronto, J.R.D.; Sarankhuu, B.-E.; Ko, K.S.; Rhee, B.D.; Kim, N.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Han, J. Acetylcholinesterase inhibitory activity of pigment Echinochrome A from sea urchin Scaphechinus mirabilis. Mar. Drugs. 2014, 12, 3560–3573. [Google Scholar] [CrossRef] [Green Version]
- Belan, D.V.; Polonik, S.G.; Ekimova, I.V. Efficiency of preventive therapy with chaperon inducer U133 in the model of preclinical stage of Parkinson’s disease in elderly rats. Russ. J. Physiol. 2020, 106, 1251–1265. [Google Scholar] [CrossRef]
- Polonik, S.; Likhatskaya, G.; Sabutski, Y.; Pelageev, D.; Denisenko, V.; Pislyagin, E.; Chingizova, E.; Menchinskaya, E.; Aminin, D. Synthesis, cytotoxic activity evaluation and Quantitative Structure-Activity Analysis of substituted 5,8-dihydroxy-1,4-naphthoquinones and their O- and S-glycoside derivatives tested against Neuro-2a cancer cells. Mar. Drugs. 2020, 18, 602. [Google Scholar] [CrossRef] [PubMed]
- Ekimova, I.V.; Pazi, M.B.; Belan, D.V.; Polonik, S.G.; Pastukhov, Y.F. The chaperone inducer U-133 eliminates anhedonia and prevents the development of neurodegeneration in monoaminergicemotionogenic structures of the brain in a model of the preclinical stage of Parkinson’s disease in elderly rats. Ros. Fiziol. Zh. 2021, 107. in press. [Google Scholar]
- Przedborski, S.; Ischiropoulos, H. Reactive oxygen and nitrogen species: Weapons of neuronal destruction in models of Parkinson’s disease. Antioxid Redox Signal. 2005, 7, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Ivanova, M.V.; Ruuge, E.K. How do calcium ions induce free radical oxidation of hydroxy-1,4-naphthoquinone? Ca2+ stabilizes the naphthosemiquinone anion-radical of echinochrome A. Arch. Biochem. Biophys. 2003, 413, 191–198. [Google Scholar] [CrossRef]
- Lebedev, A.V.; Ivanova, M.V.; Levitsky, D.O. Echinochrome, a naturally occurringiron chelator and free radical scavenger in artificial and natural membrane systems. Life Sci. 2005, 76, 863–875. [Google Scholar] [CrossRef]
- Lebedev, A.V.; Ivanova, M.V.; Levitsky, D.O. Iron chelators and free radicalscavengers in naturally occurring polyhydroxylated 1,4-naphthoquinones. Hemoglobin 2008, 32, 165–179. [Google Scholar] [CrossRef]
- Elyakov, G.B.; Maximov, O.B.; Mishenko, N.P.; Koltsova, E.A.; Fedoreev, S.A.; Glebko, L.I.; Krasovskaya, N.P.; Artjukov, A.A. Histochrome and Its Therapeutic Use in Acute Myocardial Infarction and Ischemic Heart Disease. U.S. Patent 6410601B2, 25 June 2002. [Google Scholar]
- Glazunov, V.P.; Berdyshev, D.V.; Novikov, V.L. DFT study of mechanisms of the antioxidant effect of natural polyhydroxy-1,4-naphthoquinones. Reactions of echinaminesa and b, metabolites of sea urchin Scaphechinus mirabilis, with hydroperoxyl radical. Rus. Chem. Bul. 2014, 63, 1993–1999. [Google Scholar] [CrossRef]
- Klotz, L.-O.; Hou, X.; Jacob, C. 1,4-Naphthoquinones: From oxidative damage to cellular and inter-cellular signaling. Molecules 2014, 19, 14902–14918. [Google Scholar] [CrossRef] [Green Version]
- Son, T.G.; Camandola, S.; Arumugam, T.V.; Cutler, R.G.; Telljohann, R.S.; Mughal, M.R.; Moore, T.A.; Luo, W.; Yu, Q.S.; Johnson, D.A.; et al. Plumbagin, a novel Nrf2/ARE activator, protects against cerebral ischemia. J. Neurochem. 2009, 112, 1316–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolton, J.L.; Dunlap, T. Formation and Biological Targets of Quinones: Cytotoxic versus Cytoprotective Effects. Chem. Res. Toxicol. 2017, 30, 13–37. [Google Scholar] [CrossRef]
- Crippen, G.M. VRI: 3D QSAR at variable resolution. J. Comput. Chem. 1999, 20, 1577–1585. [Google Scholar] [CrossRef]
- Labute, P. A widely applicable set of descriptors. J. Mol. Graph. Model. 2000, 18, 464–477. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast Calculation of Molecular Polar Surface Area as a Sum of Fragment Based Contributions and Its Application to the Prediction of Drug Transport Properties. J. Med. Chem. 2000, 43, 3714–3717. [Google Scholar] [CrossRef]
- Cruciania, G.; Crivorib, P.; Carruptb, P.-A.; Testab, B. Molecular fields in quantitative structure–permeation relationships: The VolSurf approach. J. Mol. Struct. 2000, 503, 17–30. [Google Scholar] [CrossRef]
- Molecular Operating Environment (MOE). Chemical Computing Group ULC, 1010 Sherbrooke St. West, Suite #910; Montreal, QC, Canada. 2019. H3A 2R7. Available online: http://chemcomp.com (accessed on 10 September 2021).
- Yurchenko, E.A.; Menchinskaya, E.S.; Pislyagin, E.A.; Trinh, P.T.H.; Ivanets, E.V.; Smetanina, O.F.; Yurchenko, A.N. Neuroprotective activity of some marine fungal metabolites in the 6-Hydroxydopamin- and paraquat-induced Parkinson’s disease models. Mar. Drugs. 2018, 16, 457. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Yurchenko, E.A.; Kolesnimova, S.A.; Lyakhova, E.G.; Menchinskaya, E.S.; Pislyagin, E.A.; Chingizova, E.A.; Aminin, D.L. Lanostane Triterpenoid Metabolites from a Penares sp. Marine Sponge Protect Neuro-2a Cells against Paraquat Neurotoxicity. Molecules 2020, 25, 5397. [Google Scholar] [CrossRef] [PubMed]
- Application Note. Precise and Accurate Counts and Viability Measurements Across Multiple Cell Lines Using the Muse™ Cell Count and Viability Assay. Available online: www.luminexcorp.com/wp-content/uploads/2018/11/AN3312EN00EMD.pdf?x80702 (accessed on 10 September 2021).
- Bhoora, S.; Punchoo, R. Policing cancer: Vitamin D arrests the cell cycle. Int. J. Mol. Sci. 2020, 21, 9296. [Google Scholar] [CrossRef] [PubMed]
- Pislyagin, E.; Kozlovskiy, S.; Menchinskaya, E.; Chingizova, E.; Likhatskaya, G.; Gorpenchenko, T.; Sabutski, Y.; Polonik, S.; Aminin, D. Synthetic 1,4-naphthoquinones inhibit P2X7 receptors in murine neuroblastoma cells. Bioorg. Med. Chem. 2021, 31, 115975. [Google Scholar] [CrossRef] [PubMed]
- Leutou, A.S.; Yun, K.; Son, B.W. Induced production of 6,9-dibromoflavasperone, a new radical scavenging naphthopyranone in the marine-mudflat-derived fungus Aspergillus Niger. Arch. Pharm. Res. 2016, 39, 806–810. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | ||||||
| # | Code | R1 | R2 | R3 | R4 | EC50, μM * |
| 1. Basic naphthoquinones | ||||||
| 1 | U-193 | H | H | H | H | >100 |
| 2 | U-139 | HO | H | H | H | >100 |
| 3 | U-574 | HO | Me | H | H | >100 |
| 4 | U-575 | HO | Et | H | H | >100 |
| 5 | U-134 | HO | HO | H | H | >100 |
| 6 | U-572 | HO | HO | Me | H | >100 |
| 7 | U-573 | HO | HO | Et | H | >100 |
| 8 | U-138 | HO | HO | Et | HO | >100 |
| 9 | U-195 | HO | H | Cl | Cl | 23.10 |
| 10 | U-434 | HO | H | Me | Me | 82.75 |
| 11 | U-622 | HO | H | H | MeO | >100 |
| 12 | U-623 | HO | H | MeO | MeO | >100 |
| 13 | U-504 | HO | H | HO | HO | >100 |
| 14 | U-642 |  | >100 | |||
| 15 | U-643 |  | >100 | |||
| 2. Acetylated naphthoquinone O-glucosides | ||||||
| 16 | U-127 | AGO | H | H | H | 16.43 |
| 17 | U-132 | GO | H | H | H | 87.40 |
| 18 | U-444 | AGO | Me | H | H | 5.33 |
| 19 | U-443 | AGO | Et | H | H | 4.46 |
| 20 | U-330 | AGO | H | Me | Me | 9.43 |
| 21 | U-136 | AGO | AGO | H | H | 10.60 |
| 22 | U-420 | AGO | AGO | Me | H | 4.46 |
| 23 | U-421 | AGO | AGO | Et | H | 8.84 |
| 24 | U-133 | AGO | AGO | Et | AGO | 8.45 |
| 3. Acetylated naphthoquinone S-glucosides | ||||||
| 25 | U-633 | HO | AGSCH2 | H | H | >100 |
| 26 | U-519 | HO | AGSCH2 | Me | Me | 84.00 |
| 27 | U-518 | HO | AGSCH2 | Cl | Cl | 32.20 |
| 28 | U-639 | HO | AGSCH2 | H | MeO | >100 |
| 29 | U-637 | HO | AGSCH2 | MeO | MeO | >100 |
| 30 | U-634 | MeO | AGSCH2 | H | H | 2.72 |
| 31 | U-521 | MeO | AGSCH2 | Me | Me | >100 |
| 32 | U-523 | MeO | AGSCH2 | Cl | Cl | 3.14 |
| 33 | U-645 | MeO | AGSCH2 | H | MeO | 11.61 |
| 34 | U-638 | MeO | AGSCH2 | MeO | MeO | 11.05 |
| 4. Deacetylated naphthoquinone S-glucosides | ||||||
| 35 | U-635 | HO | GSCH2 | H | H | >100 |
| 36 | U-520 | HO | GSCH2 | Me | Me | >100 |
| 37 | U-624 | HO | GSCH2 | Cl | Cl | >100 |
| 38 | U-644 | HO | GSCH2 | H | MeO | >100 |
| 39 | U-640 | HO | GSCH2 | MeO | MeO | >100 |
| 40 | U-636 | MeO | GSCH2 | H | H | 19.02 |
| 41 | U-522 | MeO | GSCH2 | Me | Me | 11.47 |
| 42 | U-625 | MeO | GSCH2 | Cl | Cl | 38.02 |
| 43 | U-646 | MeO | GSCH2 | H | MeO | 80.76 |
| 44 | U-641 | MeO | GSCH2 | MeO | MeO | 61.60 |
 | ||||||
| Code | Scavenging of DPPH, % | ||
|---|---|---|---|
| 0.01 µM | 0.1 µM | 1.0 µM | |
| U-134 | 14.55 ± 1.30 * | 16.87 ± 3.57 * | 30.85 ± 3.57 * |
| U-572 | 18.95 ± 0.52 * | 16.13 ± 2.03 * | 28.22 ± 1.16 * |
| U-623 | 16.08 ± 2.79 * | 15.46 ± 1.09 * | 30.34 ± 4.11 * |
| U-643 | 6.52 ± 0.02 * | 14.04 ± 2.73 * | 21.87 ± 1.01 * |
| U-625 | 9.57 ± 0.94 * | 14.29 ± 0.45 * | 30.25 ± 0.55 * |
| Ascorbic acid | 4.23 ± 0.16 | 6.59 ± 0.27 | 47.98 ± 3.48 |
| Class | Code | Description |
|---|---|---|
| 2D | SMR_VSA | Molecular refractivity (including implicit hydrogens). This property is an atomic contribution that assumes the correct protonation state (washed structures). Sum of vi such that Ri is in [0, 0.11]. |
| 2D | SlogP_VSA | Log of the octanol/water partition coefficient (including implicit hydrogens). This property is an atomic contribution model that calculates logP from the given structure; i.e., the correct protonation state). Sum of vi such that Li is in [−0.2, 0]. |
| 2D | TPSA | Topological polar surface area (Å2) calculated using group contributions to approximate the polar surface area from connection table information only. |
| 3D | vsurf_D | Hydrophobic volume. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menchinskaya, E.; Chingizova, E.; Pislyagin, E.; Likhatskaya, G.; Sabutski, Y.; Pelageev, D.; Polonik, S.; Aminin, D. Neuroprotective Effect of 1,4-Naphthoquinones in an In Vitro Model of Paraquat and 6-OHDA-Induced Neurotoxicity. Int. J. Mol. Sci. 2021, 22, 9933. https://doi.org/10.3390/ijms22189933
Menchinskaya E, Chingizova E, Pislyagin E, Likhatskaya G, Sabutski Y, Pelageev D, Polonik S, Aminin D. Neuroprotective Effect of 1,4-Naphthoquinones in an In Vitro Model of Paraquat and 6-OHDA-Induced Neurotoxicity. International Journal of Molecular Sciences. 2021; 22(18):9933. https://doi.org/10.3390/ijms22189933
Chicago/Turabian StyleMenchinskaya, Ekaterina, Ekaterina Chingizova, Evgeny Pislyagin, Galina Likhatskaya, Yuri Sabutski, Dmitry Pelageev, Sergei Polonik, and Dmitry Aminin. 2021. "Neuroprotective Effect of 1,4-Naphthoquinones in an In Vitro Model of Paraquat and 6-OHDA-Induced Neurotoxicity" International Journal of Molecular Sciences 22, no. 18: 9933. https://doi.org/10.3390/ijms22189933