
Deficiency of AMPKα1 Exacerbates Intestinal Injury and Remote Acute Lung Injury in Mesenteric Ischemia and Reperfusion in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
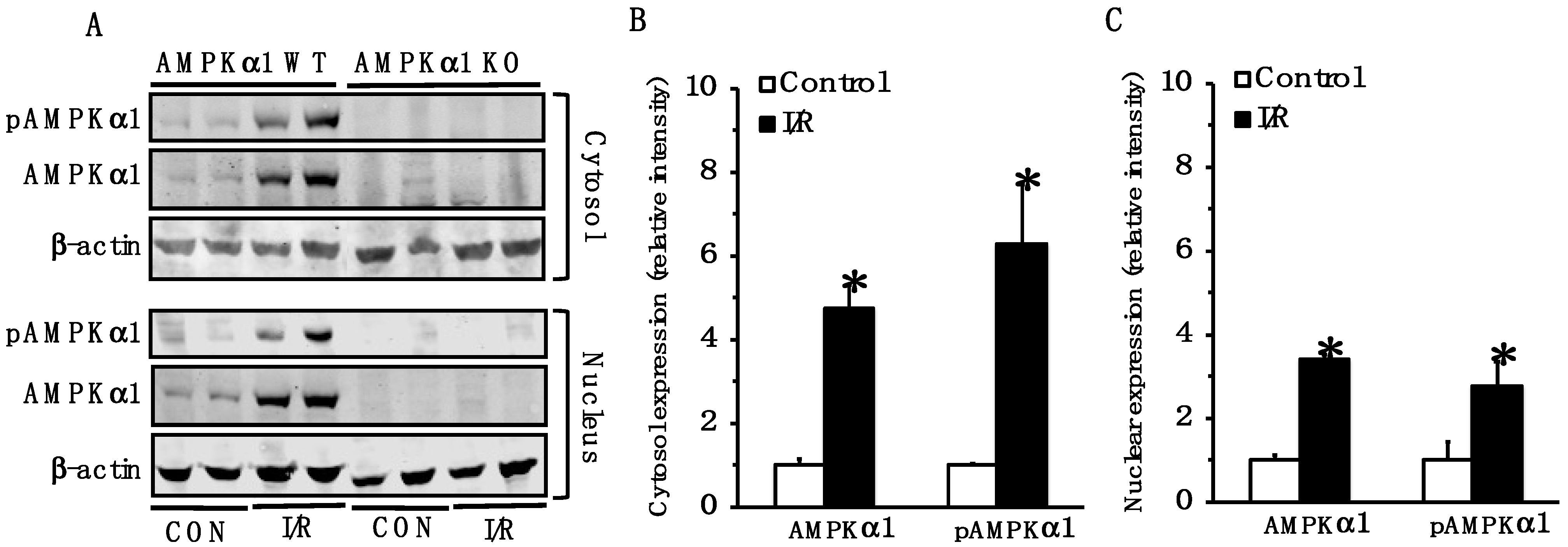
2.1. AMPK Is Activated in Small Intestine Following Mesenteric I/R Injury
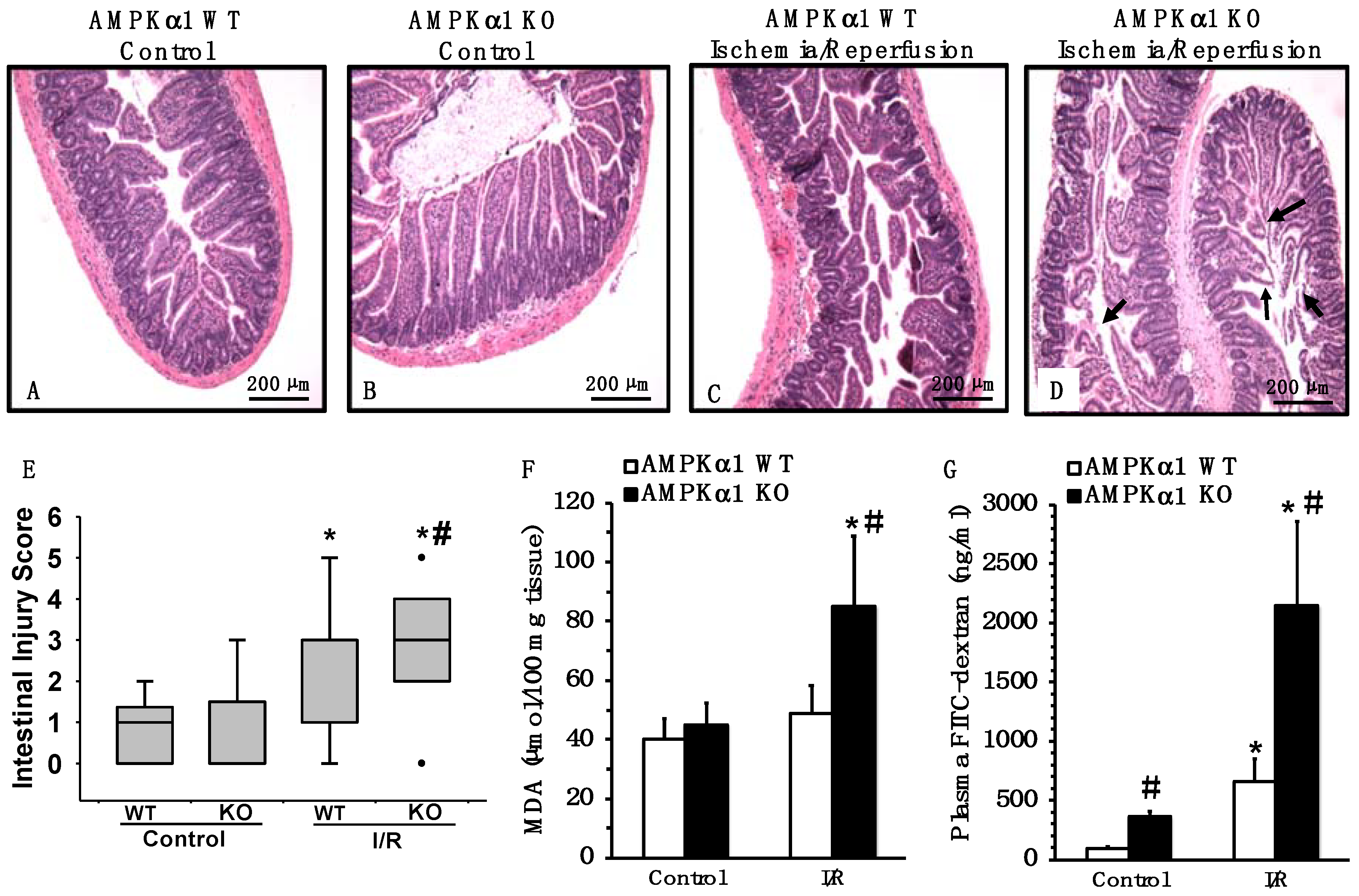
2.2. Genetic Deficiency of AMPKα1 Exacerbates Mesenteric I/R Injury
2.3. Genetic Deficiency of AMPKα1 Is Associated with Increased Epithelial Permeability after Mesenteric I/R
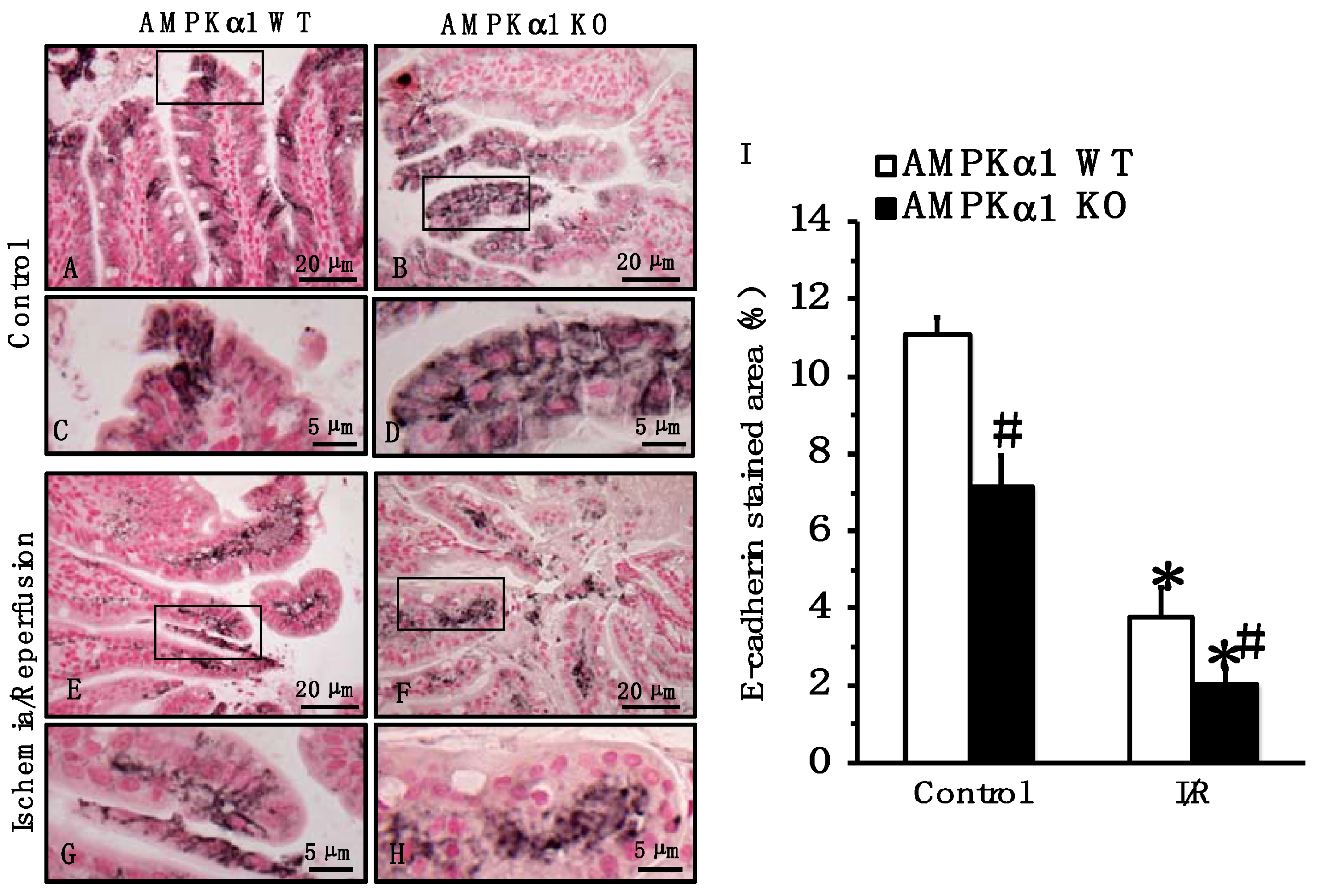
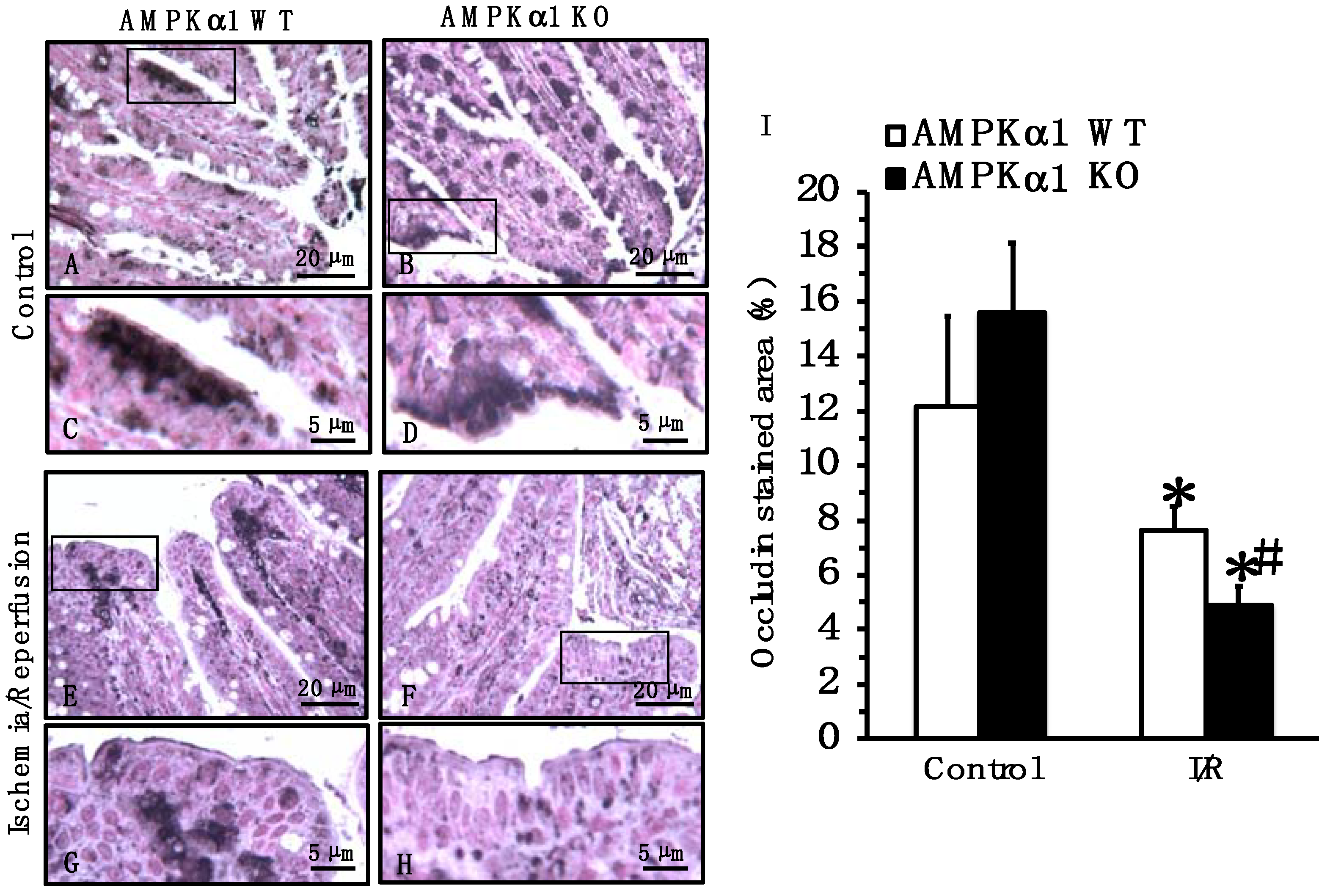
2.4. Genetic Deficiency of AMPKα1 Causes Reduction in Occludin and E-Cadherin Expression after Mesenteric I/R
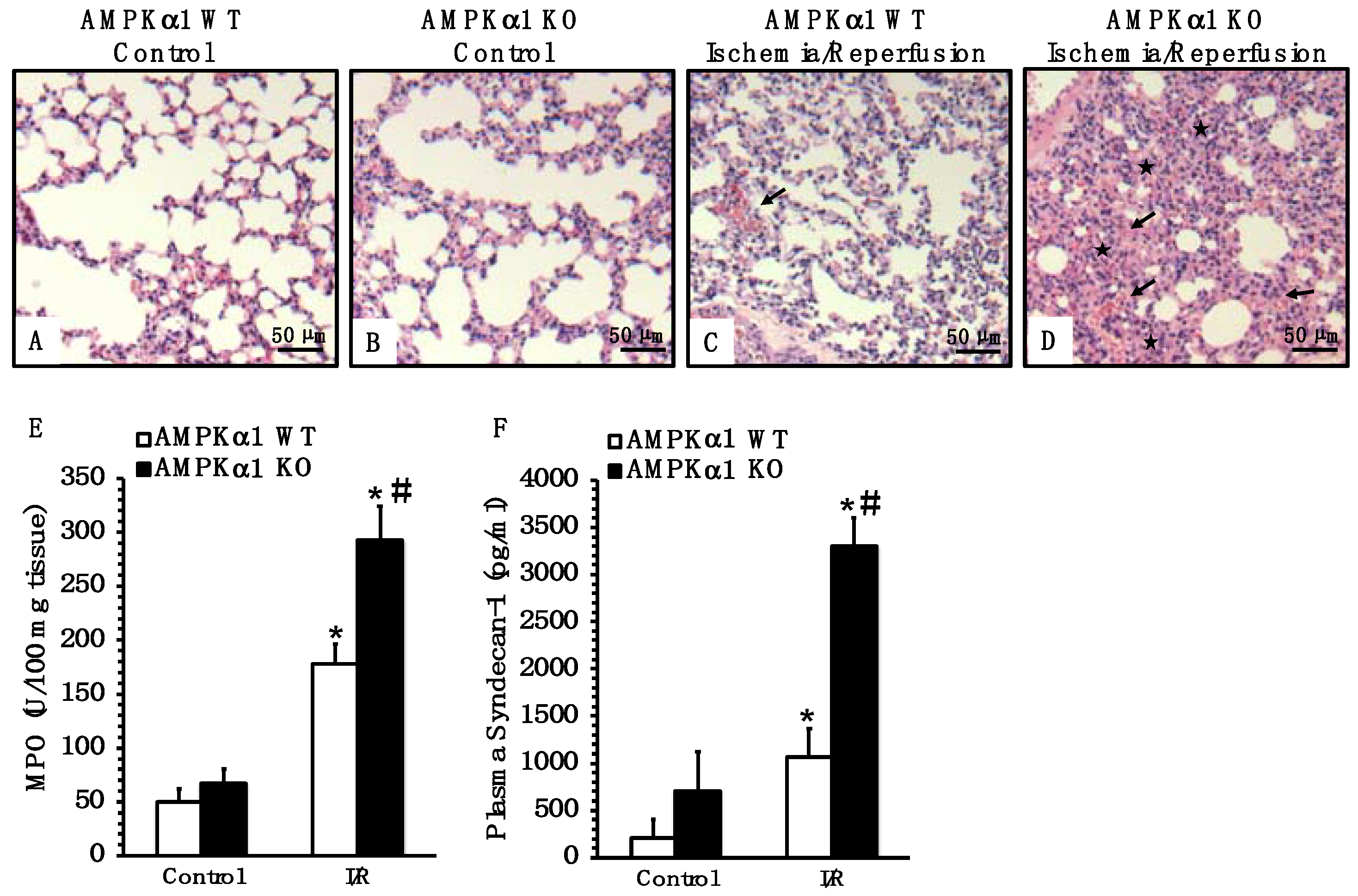
2.5. Genetic Deficiency of AMPKα1 Exacerbates Lung Injury after Mesenteric I/R
2.6. Genetic Deficiency of AMPKα1 Is Associated with Endothelial Glycocalyx Damage after Mesenteric I/R
3. Discussion
4. Materials and Methods
4.1. Murine Model of Mesenteric I/R Injury
4.2. Cytosol and Nuclear Protein Extraction
4.3. Western Blot Analysis
4.4. Histological Analysis of Intestinal Injury
4.5. In Vivo Measurement of Intestinal Permeability
4.6. Malondialdehyde Assay
4.7. Immunohistochemistry
4.8. Histological Analysis of Lung Injury
4.9. Measurement of Myeloperoxidase Activity
4.10. Plasma Levels of Syndecan-1
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AJ | adherent junction |
| AMP | adenosine monophosphate |
| AMPK | AMP-activated protein kinase |
| ATP | adenosine triphosphate |
| FITC-dextran | fluorescein isothiocyanate-conjugated dextran |
| I/R | ischemia and reperfusion |
| KO | knock-out |
| MDA | malondialdehyde |
| MODS | multiple organ dysfunction syndrome |
| MPO | Myeloperoxidase |
| PCR | polymerase chain reaction |
| TJ | tight junction |
| WT | wild-type |
References
- Guan, Y.; Worrell, R.T.; Pritts, T.A.; Montrose, M.H. Intestinal ischemia-reperfusion injury: Reversible and irreversible damage imaged in vivo. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G187–G196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clair, D.G.; Beach, J.M. Mesenteric Ischemia. N. Engl. J. Med. 2016, 374, 959–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollmar, B.; Menger, M.D. Intestinal ischemia/reperfusion: Microcirculatory pathology and functional consequences. Langenbeck’s Arch. Surg. 2011, 396, 13–29. [Google Scholar] [CrossRef]
- Rubenfeld, G.D.; Caldwell, E.; Peabody, E.; Weaver, J.; Martin, D.P.; Neff, M.; Stern, E.J.; Hudson, L.D. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005, 353, 1685–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.A.; Nelson, W.J.; Chavez, N. Cell-cell junctions organize structural and signaling networks. Cold Spring Harb. Perspect. Biol. 2018, 10, a029181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. 2010, 5, 119–144. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell. Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Carling, D.; Viollet, B. Beyond energy homeostasis: The expanding role of AMP-activated protein kinase in regulating metabolism. Cell Metab. 2015, 21, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Quentin, T.K.J.; Steinmetz, M.; Poppe, A.; Bar, K.; Kratzner, R. Different expression of the catalytic alpha subunits of the AMP activated protein kinase-an immunohistochemical study in human tissue. Histol. Histopath. 2011, 26, 589–596. [Google Scholar]
- Harmel, E.; Grenier, E.; Ben Djoudi Ouadda, A.; El Chebly, M.; Ziv, E.; Beaulieu, J.F.; Sané, A.; Spahis, S.; Laville, M.; Levy, E. AMPK in the small intestine in normal and pathophysiological conditions. Endocrinology 2014, 155, 873–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingbeil, L.R.; Kim, P.; Piraino, G.; O’Connor, M.; Hake, P.W.; Wolfe, V.; Zingarelli, B. Age-dependent changes in AMPK metabolic pathways in the lung in a mouse model of hemorrhagic shock. Am. J. Respir. Cell Mol. Biol. 2017, 56, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsiukevich, D.; Piraino, G.; Klingbeil, L.R.; Hake, P.W.; Wolfe, V.; O’Connor, M.; Zingarelli, B. The AMPK activator AICAR ameliorates age-dependent myocardial injury in murine hemorrhagic shock. Shock 2017, 47, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsiukevich, D.; Piraino, G.; Lahni, P.; Hake, P.W.; Wolfe, V.; O’Connor, M.; James, J.; Zingarelli, B. Metformin ameliorates gender-and age-dependent hemodynamic instability and myocardial injury in murine hemorrhagic shock. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2680–2691. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Piraino, G.; O’Connor, M.; Hake, P.W.; Wolfe, V.; Lahni, P.; Zingarelli, B. Metformin exerts beneficial effects in hemorrhagic shock in an AMPKα1-independent manner. Shock 2018, 49, 277–287. [Google Scholar] [CrossRef]
- Yu, H.; Kalogeris, T.; Korthuis, R.J. Reactive species-induced microvascular dysfunction in ischemia/reperfusion. Free Radic. Biol. Med. 2019, 135, 182–197. [Google Scholar] [CrossRef]
- Dogné, S.; Flamion, B. Endothelial glycocalyx impairment in disease: Focus on hyaluronan shedding. Am. J. Pathol. 2020, 190, 768–780. [Google Scholar] [CrossRef]
- Qi, D.; Young, L.H. AMPK: Energy sensor and survival mechanism in the ischemic heart. Trends Endocrinol. Metab. 2015, 26, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.R., III.; Li, J.; Coven, D.L.; Pypaert, M.; Zechner, C.; Palmeri, M.; Giordano, F.J.; Mu, J.; Birnbaum, M.J.; Young, L.H. AMP-activated protein kinase mediates ischemic glucose uptake and prevents postischemic cardiac dysfunction, apoptosis, and injury. J. Clin. Investig. 2004, 114, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Paiva, M.A.; Gonçalves, L.M.; Providência, L.A.; Davidson, S.M.; Yellon, D.M.; Mocanu, M.M. Transitory activation of AMPK at reperfusion protects the ischaemic-reperfused rat myocardium against infarction. Cardiovasc. Drugs Ther. 2010, 24, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, K.; Zarrinpashneh, E.; Szarszoi, O.; Joubert, F.; Athea, Y.; Mateo, P.; Vaulont, S.; Viollet, B.; Bigard, X.; Bertrand, L.; et al. Dual cardiac contractile effects of the α2-AMPK deletion in low-flow ischemia and reperfusion. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H3136–H3147. [Google Scholar] [CrossRef] [Green Version]
- Lieberthal, W.; Tang, M.; Lusco, M.; Abate, M.; Levine, J.S. Preconditioning mice with activators of AMPK ameliorates ischemic acute kidney injury in vivo. Am. J. Physiol. Renal Physiol. 2016, 311, F731–F739. [Google Scholar] [CrossRef]
- Rodríguez, C.; Contreras, C.; Sáenz-Medina, J.; Muñoz, M.; Corbacho, C.; Carballido, J.; García-Sacristán, A.; Hernandez, M.; López, M.; Rivera, L.; et al. Activation of the AMP-related kinase (AMPK) induces renal vasodilatation and downregulates Nox-derived reactive oxygen species (ROS) generation. Redox Biol. 2020, 34, 101575. [Google Scholar] [CrossRef]
- Song, P.; Zou, M.H. Regulation of NAD(P)H oxidases by AMPK in cardiovascular systems. Free Radic. Biol. Med. 2012, 52, 1607–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, S.L.; Moncada, S. AMPKα1 regulates the antioxidant status of vascular endothelial cells. Biochem. J. 2009, 421, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, D.; Kiyosue, A.; Takahashi, M.; Satonaka, H.; Tanaka, K.; Sata, M.; Nagano, T.; Nagai, R.; Hirata, Y. A new constitutively active mutant of AMP-activated protein kinase inhibits anoxia-induced apoptosis of vascular endothelial cell. Hypertension Res. 2009, 32, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inata, Y.; Kikuchi, S.; Samraj, R.S.; Hake, P.W.; O’Connor, M.; Ledford, J.R.; O’Connor, J.; Lahni, P.; Wolfe, V.; Piraino, G.; et al. Autophagy and mitochondrial biogenesis impairment contributes to age-dependent liver injury in experimental sepsis: Dysregulation of AMP-activated protein kinase pathway. FASEB J. 2018, 32, 728–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serino, M.; Waget, A.; Marsollier, N.; Masseboeuf, M.; Payros, G.; Kabani, C.; Denom, J.; Lacombe, A.; Thiers, J.C.; Negre-Salvayre, A.; et al. Lipid-induced peroxidation in the intestine is involved in glucose homeostasis imbalance in mice. PLoS ONE 2011, 6, e21184. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, M.B.; Dhawan, P.; Baumert, T.F. Tight junction proteins in gastrointestinal and liver disease. Gut 2019, 68, 547–561. [Google Scholar] [CrossRef]
- Takeichi, M. Dynamic contacts: Rearranging adherens junctions to drive epithelial remodelling. Nat. Rev. Mol. Cell Biol. 2014, 15, 397–410. [Google Scholar] [CrossRef]
- Han, X.; Fink, M.P.; Delude, R.L. Proinflammatory cytokines cause NO*-dependent and -independent changes in expression and localization of tight junction proteins in intestinal epithelial cells. Shock 2003, 19, 229–237. [Google Scholar] [CrossRef]
- Bruewer, M.; Utech, M.; Ivanov, A.I.; Hopkins, A.M.; Parkos, C.A.; Nusrat, A. Interferon-gamma induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 2005, 19, 923–933. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Johnson, A.M.; Sladojevic, N.; Keep, R.F.; Andjelkovic, A.V. Endocytosis of tight junction proteins and the regulation of degradation and recycling. Ann. N.Y. Acad. Sci. 2017, 1397, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yang, Q.; Rogers, C.J.; Du, M.; Zhu, M.J. AMPK improves gut epithelial differentiation and barrier function via regulating Cdx2 expression. Cell Death Differ. 2017, 24, 819–831. [Google Scholar] [CrossRef]
- Olivier, S.; Leclerc, J.; Grenier, A.; Foretz, M.; Tamburini, J.; Viollet, B. AMPK activation promotes tight junction assembly in intestinal epithelial Caco-2 cells. Int. J. Mol. Sci. 2019, 20, 5171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, K.T.; Tsukamoto, T.; Nigam, S.K. Selective degradation of E-cadherin and dissolution of E-cadherin-catenin complexes in epithelial ischemia. Am. J. Physiol. Renal Physiol. 2000, 278, F847–F852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Suzuki, T.; Seth, A.; Samak, G.; Rao, R. Protein kinase Czeta phosphorylates occludin and promotes assembly of epithelial tight junctions. Biochem. J. 2011, 437, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Elias, B.C.; Seth, A.; Shen, L.; Turner, J.R.; Giorgianni, F.; Desiderio, D.; Guntaka, R.; Rao, R. PKC eta regulates occludin phosphorylation and epithelial tight junction integrity. Proc. Natl. Acad. Sci. USA 2009, 106, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Park, H.Y.; Kunitake, Y.; Hirasaki, N.; Tanaka, M.; Matsui, T. Theaflavins enhance intestinal barrier of Caco-2 Cell monolayers through the expression of AMP-activated protein kinase-mediated Occludin, Claudin-1, and ZO-1. Biosci. Biotechnol. Biochem. 2015, 79, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. L-glutamine enhances tight junction integrity by activating CaMK kinase 2-AMP-activated protein kinase signaling in intestinal porcine epithelial cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bays, J.L.; Campbell, H.K.; Heidema, C.; Sebbagh, M.; DeMali, K.A. Linking E-cadherin mechanotransduction to cell metabolism through force-mediated activation of AMPK. Nat. Cell. Biol. 2017, 19, 724–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingensmith, N.J.; Coopersmith, C.M. The gut as the motor of multiple organ dysfunction in critical illness. Crit. Care Clin. 2016, 32, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Smart, L.; Bosio, E.; Macdonald, S.P.J.; Dull, R.; Fatovich, D.M.; Neil, C.; Arendts, G. Glycocalyx biomarker syndecan-1 is a stronger predictor of respiratory failure in patients with sepsis due to pneumonia, compared to endocan. J. Crit. Care 2018, 47, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Viollet, B.; Athea, Y.; Mounier, R.; Guigas, B.; Zarrinpashneh, E.; Horman, S.; Lantier, L.; Hebrard, S.; Devin-Leclerc, J.; Beauloye, C.; et al. AMPK: Lessons from transgenic and knockout animals. Front. Biosci. 2009, 14, 19–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Quaedackers, J.S.; Beuk, R.J.; Bennet, L.; Charlton, A.; oude Egbrink, M.G.; Gunn, A.J.; Heineman, E. An evaluation of methods for grading histologic injury following ischemia/reperfusion of the small bowel. Transplant. Proc. 2000, 32, 1307–1310. [Google Scholar] [CrossRef]
- Feinman, R.; Deitch, E.A.; Watkins, A.C.; Abungu, B.; Colorado, I.; Kannan, K.B.; Sheth, S.U.; Caputo, F.J.; Lu, Q.; Ramanathan, M.; et al. HIF-1 mediates pathogenic inflammatory responses to intestinal ischemia-reperfusion injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G833–G843. [Google Scholar] [CrossRef] [Green Version]
- Woting, A.; Blaut, M. Small intestinal permeability and gut-transit time determined with low and high molecular weight Fluorescein Isothiocyanate-Dextrans in C3H Mice. Nutrients 2018, 10, 685. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, H.; Yagi, K. Assay for lipid peroxides in animal tissue by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Mullane, K.M.; Kraemer, R.; Smith, B. Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischemic myocardium. J. Pharmacol. Methods 1985, 14, 157–167. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayes, H.V.; Wolfe, V.; O’Connor, M.; Levinsky, N.C.; Piraino, G.; Zingarelli, B. Deficiency of AMPKα1 Exacerbates Intestinal Injury and Remote Acute Lung Injury in Mesenteric Ischemia and Reperfusion in Mice. Int. J. Mol. Sci. 2021, 22, 9911. https://doi.org/10.3390/ijms22189911
Hayes HV, Wolfe V, O’Connor M, Levinsky NC, Piraino G, Zingarelli B. Deficiency of AMPKα1 Exacerbates Intestinal Injury and Remote Acute Lung Injury in Mesenteric Ischemia and Reperfusion in Mice. International Journal of Molecular Sciences. 2021; 22(18):9911. https://doi.org/10.3390/ijms22189911
Chicago/Turabian StyleHayes, Hannah V., Vivian Wolfe, Michael O’Connor, Nick C. Levinsky, Giovanna Piraino, and Basilia Zingarelli. 2021. "Deficiency of AMPKα1 Exacerbates Intestinal Injury and Remote Acute Lung Injury in Mesenteric Ischemia and Reperfusion in Mice" International Journal of Molecular Sciences 22, no. 18: 9911. https://doi.org/10.3390/ijms22189911