
“Take It or Leave It”—Factors Regulating Competence Development and DNA Uptake in Campylobacter jejuni


Abstract
:1. Introduction
2. Results
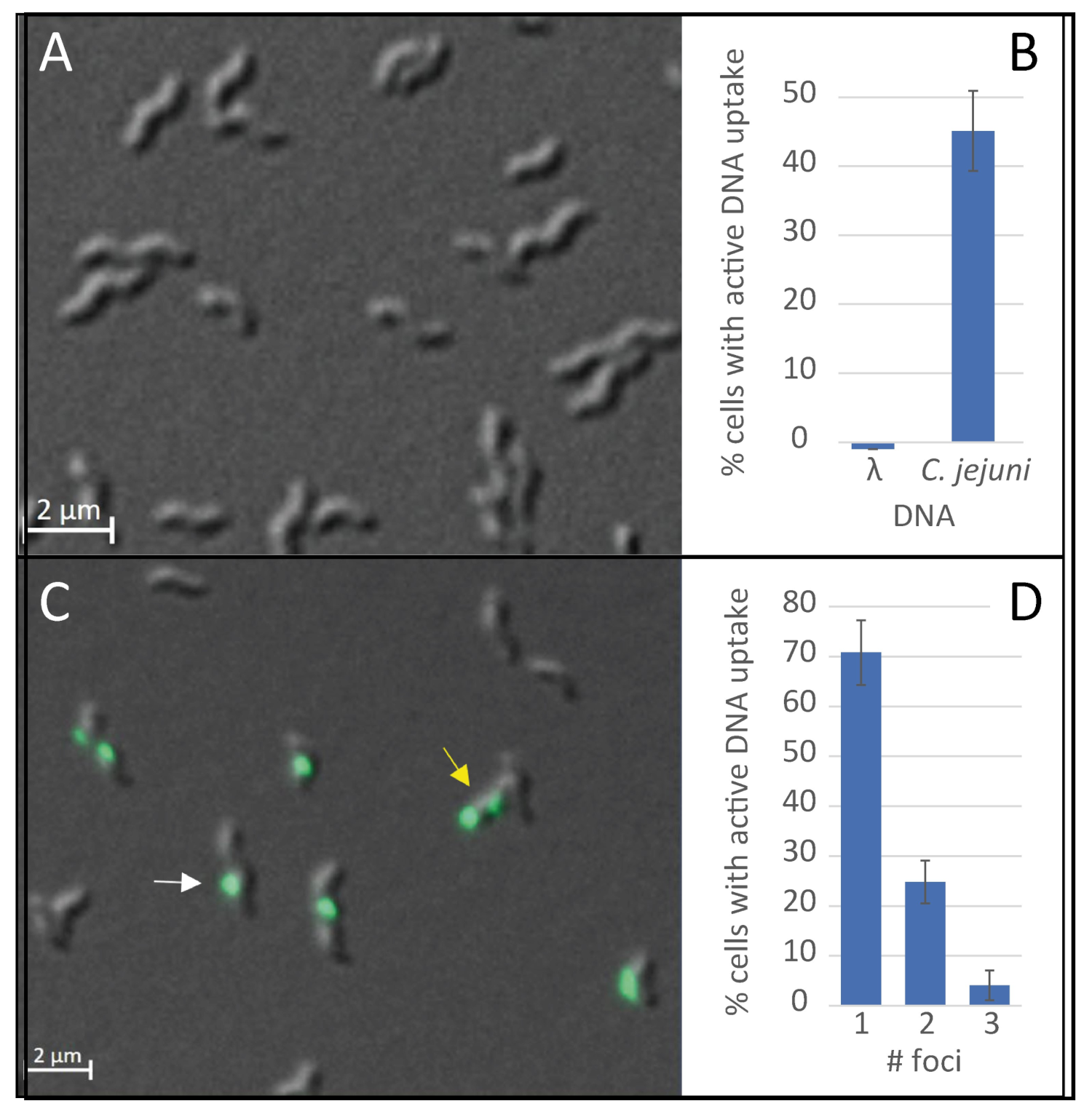
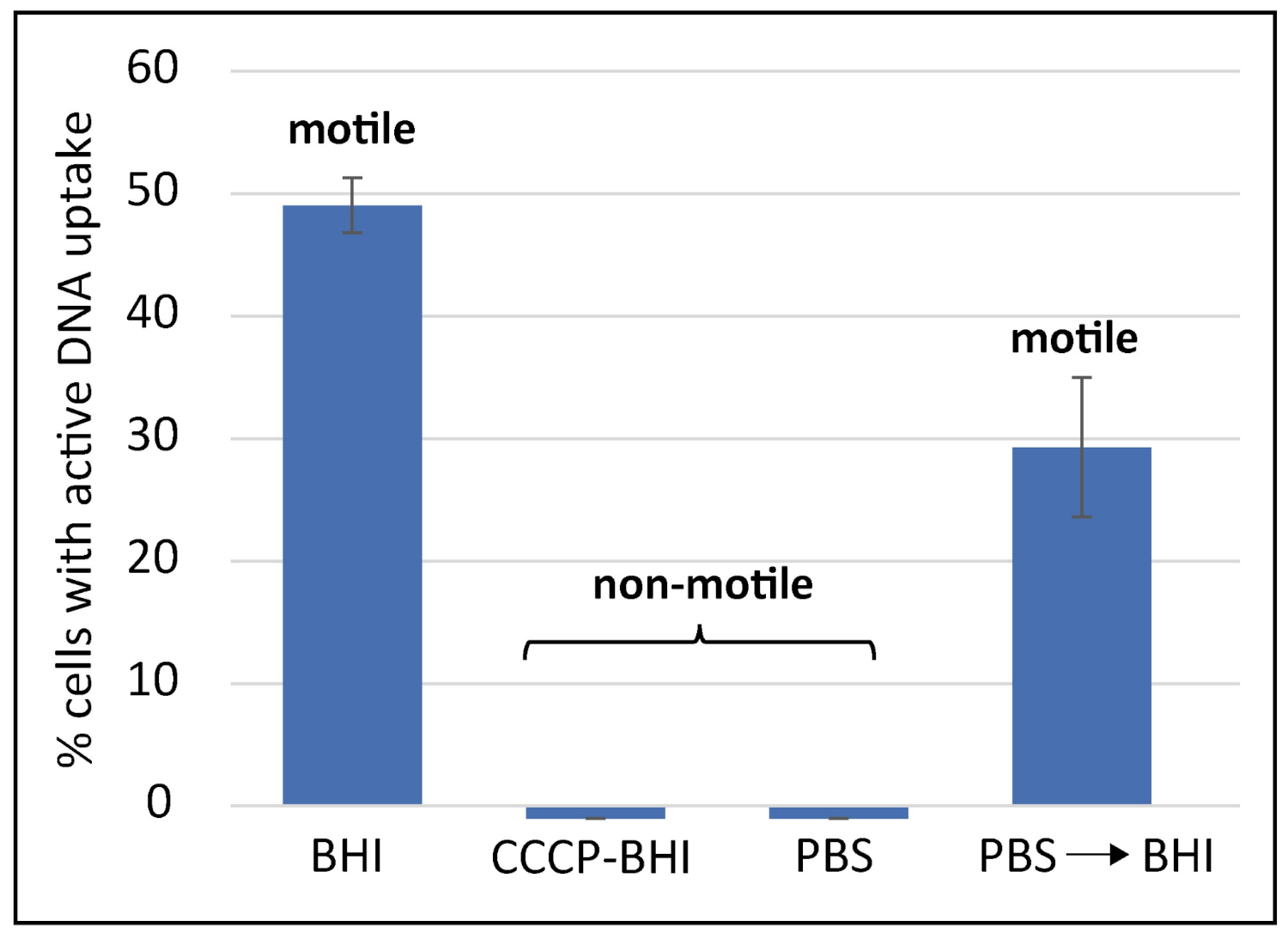
2.1. DNA Uptake in C. jejuni Occurs at Distinct Locations, Is DNA Substrate Specific and Energy-Dependent
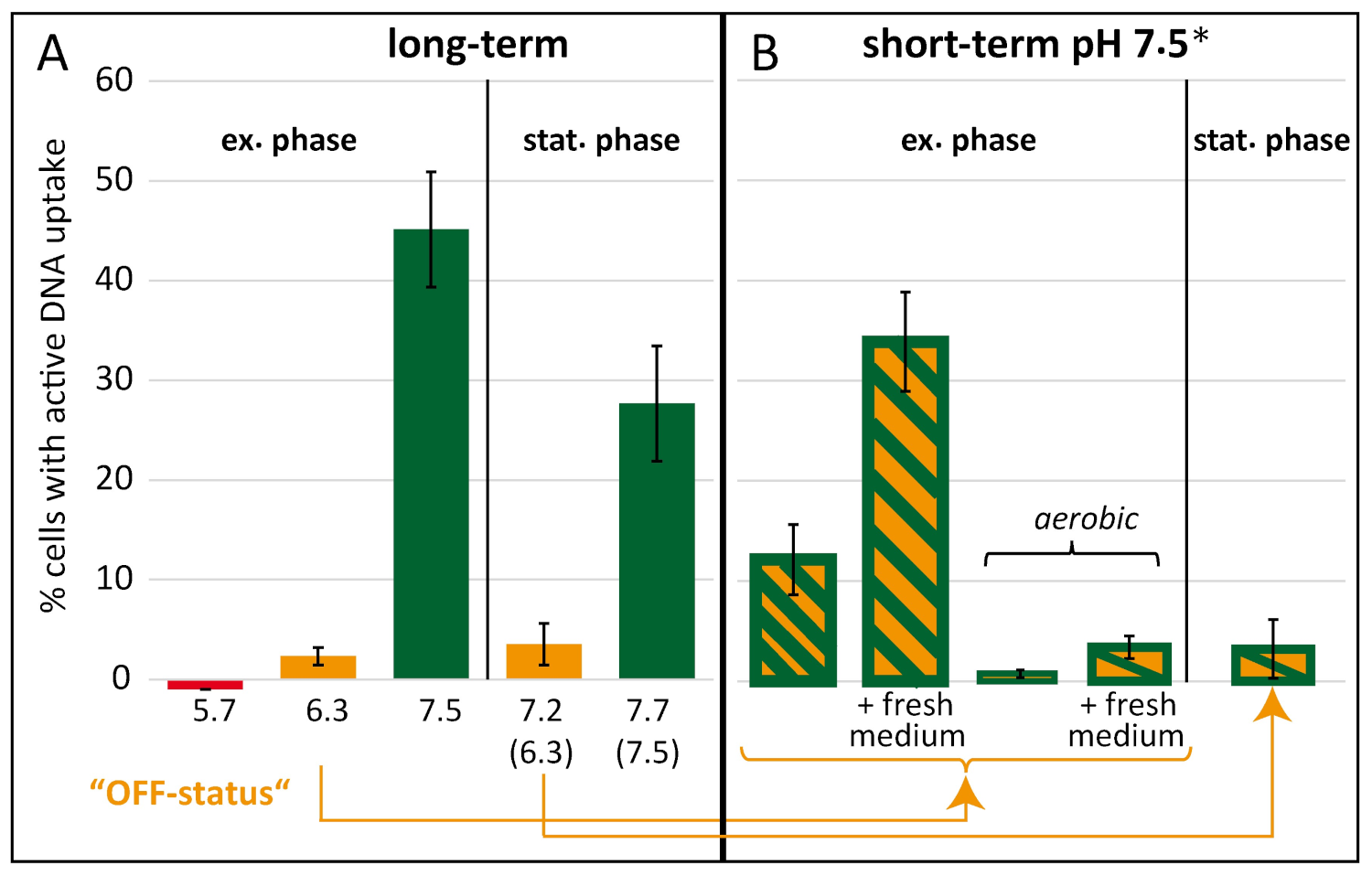
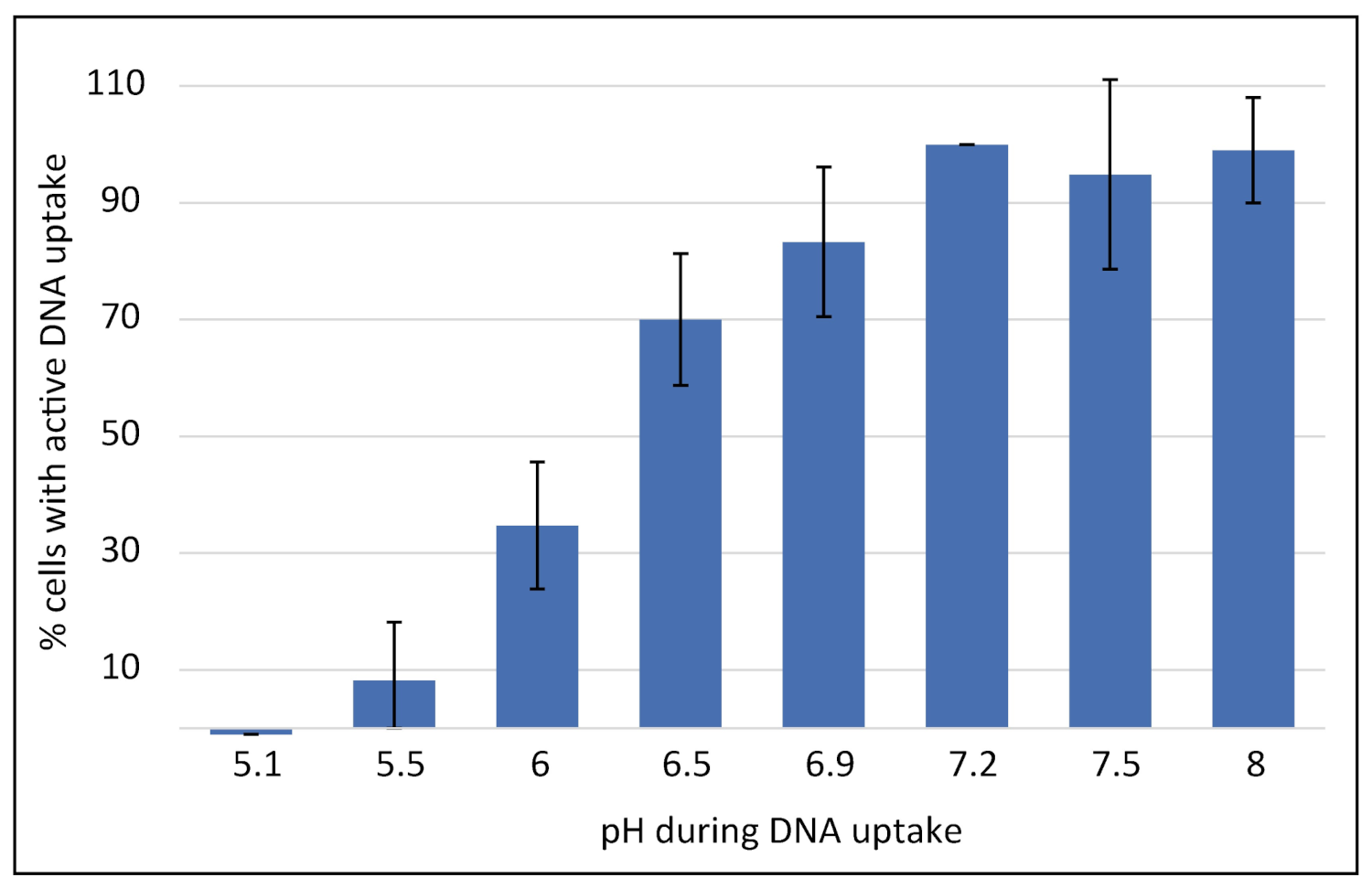
2.2. Competence Development in C. jejuni Is Dependent on External pH and Shut Down under Aerobic Conditions
2.3. Uptake of DNA into a DNase-Resistant State Is a Main Factor for Regulation of Natural Transformation
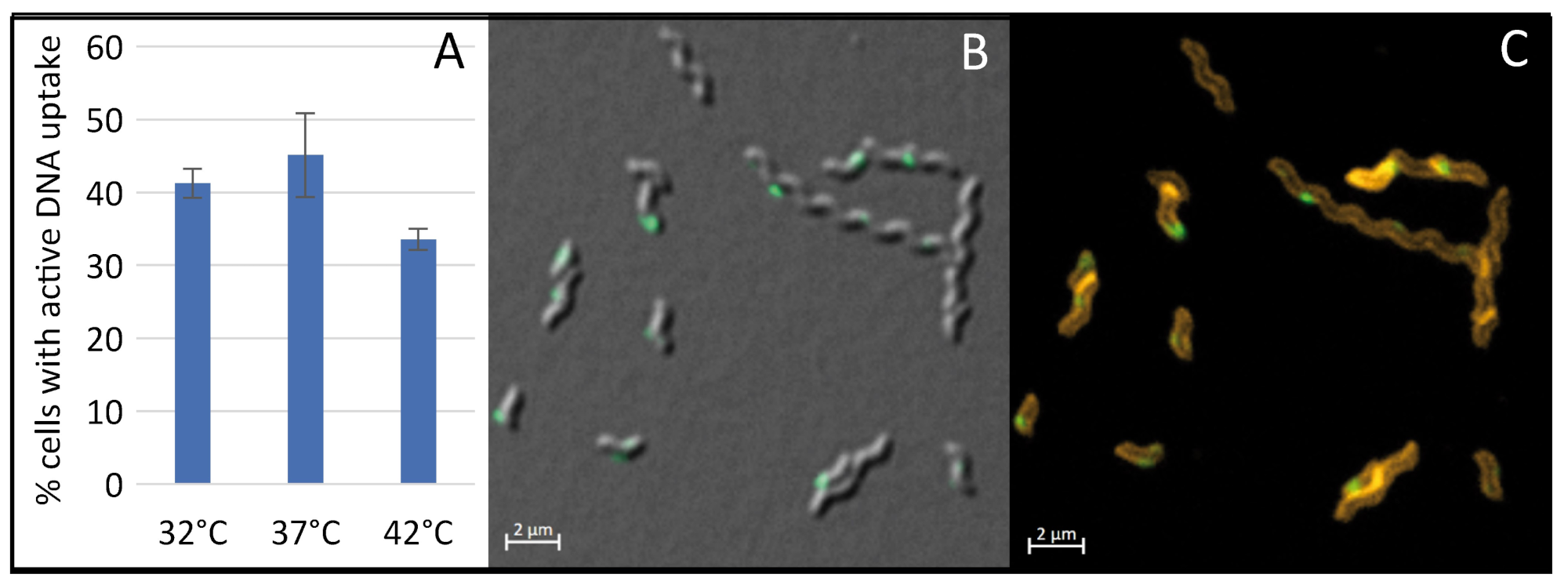
2.4. The Concentration of CO2 or Carbonate and Temperature Do Not Play a Pivotal Role in Competence Regulation
2.5. DNA Uptake Machinery of Competent Cells Was Shut Down at Slightly Acidic pH
3. Discussion
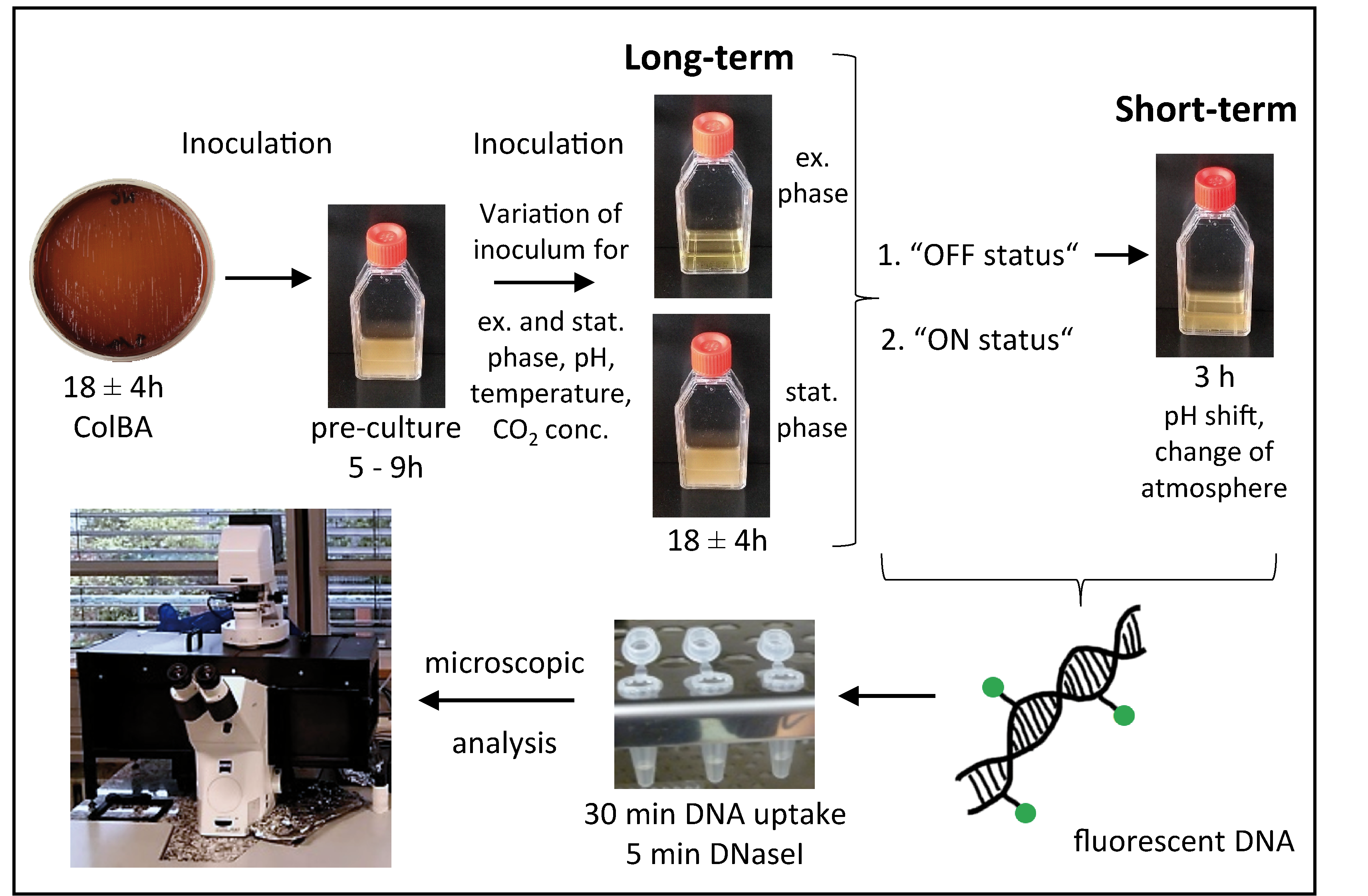
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. DNA Uptake Assay
4.3. Transformation Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar]
- Shange, N.; Gouws, P.; Hoffman, L.C. Campylobacter and Arcobacter species in food-producing animals: Prevalence at primary production and during slaughter. World J. Microbiol. Biotechnol. 2019, 35, 146. [Google Scholar] [CrossRef]
- Whiley, H.; van den Akker, B.; Giglio, S.; Bentham, R. The role of environmental reservoirs in human campylobacteriosis. Int. J. Environ. Res. Public Health 2013, 10, 5886–5907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldenström, J.; Axelsson-Olsson, D.; Olsen, B.; Hasselquist, D.; Griekspoor, P.; Jansson, L.; Teneberg, S.; Svensson, L.; Ellström, P. Campylobacter jejuni colonization in wild birds: Results from an infection experiment. PLoS ONE 2010, 5, e9082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, B.M.; Schielke, A.; Didelot, X.; Kops, F.; Breidenbach, J.; Willrich, N.; Gölz, G.; Alter, T.; Stingl, K.; Josenhans, C.; et al. A combined case-control and molecular source attribution study of human Campylobacter infections in Germany, 2011–2014. Sci. Rep. 2017, 7, 5139. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Panel on Biological Hazards (BIOHAZ). Scientific opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 2105. [Google Scholar] [CrossRef]
- Doyle, M.P.; Roman, D.J. Growth and survival of Campylobacter fetus subsp. jejuni as a function of temperature and pH. J. Food Prot. 1981, 44, 596–601. [Google Scholar] [PubMed]
- Kim, S.H.; Chelliah, R.; Ramakrishnan, S.R.; Perumal, A.S.; Bang, W.S.; Rubab, M.; Daliri, E.B.; Barathikannan, K.; Elahi, F.; Park, E.; et al. Review on stress tolerance in Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2020, 10, 596570. [Google Scholar] [CrossRef] [PubMed]
- Baffone, W.; Casaroli, A.; Citterio, B.; Pierfelici, L.; Campana, R.; Vittoria, E.; Guaglianone, E.; Donelli, G. Campylobacter jejuni loss of culturability in aqueous microcosms and ability to resuscitate in a mouse model. Int. J. Food Microbiol. 2006, 107, 83–91. [Google Scholar] [CrossRef]
- Crofts, A.A.; Poly, F.M.; Ewing, C.P.; Kuroiwa, J.M.; Rimmer, J.E.; Harro, C.; Sack, D.; Talaat, K.R.; Porter, C.K.; Gutierrez, R.L.; et al. Campylobacter jejuni transcriptional and genetic adaptation during human infection. Nat. Microbiol. 2018, 3, 494–502. [Google Scholar] [CrossRef]
- Aksomaitiene, J.; Novoslavskij, A.; Kudirkiene, E.; Gabinaitiene, A.; Malakauskas, M. Whole genome sequence-based prediction of resistance determinants in high-level multidrug-resistant Campylobacter jejuni isolates in Lithuania. Microorganisms 2020, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.Y.; Sprouffske, K.; Wagner, A. Assessing the benefits of horizontal gene transfer by laboratory evolution and genome sequencing. BMC Evol. Biol. 2018, 18, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarth, D.P.; Kwon, Y.M. Horizontal genetic exchange of chromosomally encoded markers between Campylobacter jejuni cells. PLoS ONE 2020, 15, e0241058. [Google Scholar] [CrossRef] [PubMed]
- Golz, J.C.; Epping, L.; Knüver, M.T.; Borowiak, M.; Hartkopf, F.; Deneke, C.; Malorny, B.; Semmler, T.; Stingl, K. Whole genome sequencing reveals extended natural transformation in Campylobacter impacting diagnostics and the pathogens adaptive potential. Sci. Rep. 2020, 10, 3686. [Google Scholar] [CrossRef] [PubMed]
- Golz, J.C.; Stingl, K. Natural competence and horizontal gene transfer in Campylobacter. Curr. Top Microbiol. Immunol. 2021, 431, 265–292. [Google Scholar] [PubMed]
- Attaiech, L.; Boughammoura, A.; Brochier-Armanet, C.; Allatif, O.; Peillard-Fiorente, F.; Edwards, R.A.; Omar, A.R.; MacMillan, A.M.; Glover, M.; Charpentier, X. Silencing of natural transformation by an RNA chaperone and a multitarget small RNA. Proc. Natl. Acad. Sci. USA 2016, 113, 8813–8818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelmoer, D.J.; Donaldson, I.; Rozen, D.E. Conservative sex and the benefits of transformation in Streptococcus pneumoniae. PLoS Pathog. 2013, 9, e1003758. [Google Scholar] [CrossRef] [Green Version]
- Attaiech, L.; Charpentier, X. Silently transformable: The many ways bacteria conceal their built-in capacity of genetic exchange. Curr. Genet. 2017, 63, 451–455. [Google Scholar] [CrossRef]
- Dubnau, D.; Blokesch, M. Mechanisms of DNA uptake by naturally competent bacteria. Annu. Rev. Genet. 2019, 53, 217–237. [Google Scholar] [CrossRef]
- Johnston, C.; Martin, B.; Fichant, G.; Polard, P.; Claverys, J.P. Bacterial transformation: Distribution, shared mechanisms and divergent control. Nat. Rev. Microbiol. 2014, 12, 181–196. [Google Scholar] [CrossRef]
- Corbinais, C.; Mathieu, A.; Damke, P.P.; Kortulewski, T.; Busso, D.; Prado-Acosta, M.; Radicella, J.P.; Marsin, S. ComB proteins expression levels determine Helicobacter pylori competence capacity. Sci. Rep. 2017, 7, 41495. [Google Scholar] [CrossRef] [Green Version]
- Krüger, N.J.; Knüver, M.T.; Zawilak-Pawlik, A.; Appel, B.; Stingl, K. Genetic diversity as consequence of a microaerobic and neutrophilic lifestyle. PLoS Pathog. 2016, 12, e1005626. [Google Scholar] [CrossRef] [Green Version]
- Vegge, C.S.; Brondsted, L.; Ligowska-Marzeta, M.; Ingmer, H. Natural transformation of Campylobacter jejuni occurs beyond limits of growth. PLoS ONE 2012, 7, e45467. [Google Scholar]
- Wilson, D.L.; Bell, J.A.; Young, V.B.; Wilder, S.R.; Mansfield, L.S.; Linz, J.E. Variation of the natural transformation frequency of Campylobacter jejuni in liquid shake culture. Microbiology 2003, 149 Pt 12, 3603–3615. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Taylor, D.E. Natural transformation in Campylobacter species. J. Bacteriol. 1990, 172, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, J.M.; Leveque, R.M.; Dawid, S.; DiRita, V.J. Methylation-dependent DNA discrimination in natural transformation of Campylobacter jejuni. Proc. Natl. Acad. Sci. USA 2017, 114, E8053–E8061. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.; Zhang, Q. Cj0011c, a periplasmic single- and double-stranded DNA-binding protein, contributes to natural transformation in Campylobacter jejuni. J. Bacteriol. 2007, 189, 7399–7407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, B.; Muraoka, W.; Sahin, O.; Zhang, Q. Role of Cj1211 in natural transformation and transfer of antibiotic resistance determinants in Campylobacter jejuni. Antimicrob. Agents Chemother. 2008, 52, 2699–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesner, R.S.; Hendrixson, D.R.; DiRita, V.J. Natural transformation of Campylobacter jejuni requires components of a type II secretion system. J. Bacteriol. 2003, 185, 5408–5418. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, J.M.; Erfurt, R.S.; DiRita, V.J. Characterization and localization of the Campylobacter jejuni transformation system proteins CtsE, CtsP, and CtsX. J. Bacteriol. 2015, 197, 636–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, B.; Chen, I.; Dubnau, D.; Sheetz, M.P. DNA transport into Bacillus subtilis requires proton motive force to generate large molecular forces. Nat. Struct. Mol. Biol. 2004, 11, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stingl, K.; Müller, S.; Scheidgen-Kleyboldt, G.; Clausen, M.; Maier, B. Composite system mediates two-step DNA uptake into Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2010, 107, 1184–1189. [Google Scholar] [CrossRef] [Green Version]
- Cohen, E.J.; Nakane, D.; Kabata, Y.; Hendrixson, D.R.; Nishizaka, T.; Beeby, M. Campylobacter jejuni motility integrates specialized cell shape, flagellar filament, and motor, to coordinate action of its opposed flagella. PLoS Pathog. 2020, 16, e1008620. [Google Scholar] [CrossRef]
- Blair, D.F. Flagellar movement driven by proton translocation. FEBS Lett. 2003, 545, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Hazeleger, W.C.; Wouters, J.A.; Rombouts, F.M.; Abee, T. Physiological activity of Campylobacter jejuni far below the minimal growth temperature. Appl. Environ. Microbiol. 1998, 64, 3917–3922. [Google Scholar] [CrossRef] [Green Version]
- Biesta-Peters, E.G.; Jongenburger, I.; de Boer, E.; Jacobs-Reitsma, W.F. ISO_10272-1:2017, Microbiology of the food chain—Horizontal method for detection and enumeration of Campylobacter spp.—Part 1: Detection method. Int. J. Food Microbiol. 2019, 288, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, J.W.; Kathariou, S. Differential effects of temperature on natural transformation to erythromycin and nalidixic acid resistance in Campylobacter coli. Appl. Environ. Microbiol. 2008, 74, 6121–6125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, N.J.; Buhler, C.; Iwobi, A.N.; Huber, I.; Ellerbroek, L.; Appel, B.; Stingl, K. “Limits of control”—crucial parameters for a reliable quantification of viable Campylobacter by real-time PCR. PLoS ONE 2014, 9, e88108. [Google Scholar]
- Hahn, J.; Maier, B.; Haijema, B.J.; Sheetz, M.; Dubnau, D. Transformation proteins and DNA uptake localize to the cell poles in Bacillus subtilis. Cell 2005, 122, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Kaufenstein, M.; van der Laan, M.; Graumann, P.L. The three-layered DNA uptake machinery at the cell pole in competent Bacillus subtilis cells is a stable complex. J. Bacteriol. 2011, 193, 1633–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svihus, B. Effect of digestive tract conditions, feed processing and ingredients on response to NSP enzymes. In Enzymes in farm animal nutrition, 2nd ed.; Bedford, M.R., Partridge, G.G., Eds.; CAB International: Wallingford, UK, 2011; pp. 129–159. [Google Scholar] [CrossRef]
- Epping, L.; Golz, J.C.; Knüver, M.T.; Huber, C.; Thürmer, A.; Wieler, L.H.; Stingl, K.; Semmler, T. Comparison of different technologies for the decipherment of the whole genome sequence of Campylobacter jejuni BfR-CA-14430. Gut Pathog. 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Transformation Rate ± SD | Fraction Competent Cells ± SD (%) | Change in Transformation Rate Compared to pH 6.3 | Change in Fraction of Competent Cells Relative to pH 6.3 | Ratio Change Transformation Rate vs. Change in Fraction Competent Cells |
|---|---|---|---|---|---|
| pH 5.7 | ≤6.9 × 10−9 ± 6.2 × 10−10 | 0 | ≤0.0053-fold | ∞ | n.a. |
| pH 6.3 | 1.3 × 10−6 ± 2.5 × 10−6 | 2.3 ± 0.9 | 1.0-fold | 1.0-fold | 1.0 |
| pH 6.3 +3 h at pH 7.6 | 1.4 × 10−5 ± 7.8 × 10−6 | 12.2 ± 3.5 | 10.8-fold | 5.3-fold | 2.0 |
| pH 6.3 +3 h with fresh medium | 2.9 × 10−5 ± 2.1 × 10−5 | 33.9 ± 5.0 | 22.3-fold | 14.7-fold | 1.5 |
| pH 6.3 +3 h aerobic at pH 7.9 | 1.3 × 10−6 ± 6.4 × 10−7 | 0.8 ± 0.4 | 1.0-fold | 0.3-fold | 2.9 |
| pH 6.3 +3 h aerobic with fresh medium | 3.7 × 10−6 ± 1.2 × 10−5 | 3.4 ± 1.1 | 2.8-fold | 1.5-fold | 1.9 |
| pH 7.5 | 1.3 × 10−4 ± 2.3 × 10−4 | 45.1 ± 5.8 | 100-fold | 19.6-fold | 5.1 |
| pH 7.5 without DNA | <6.4 × 10−9 ± 2.5 × 10−9 | 0 | <0.0049-fold | ∞ | n. a. |
| pH 7.3 (stat phase) | 6.6 × 10−6 ± 4.3 × 10−6 | 3.5 ± 2.1 | 5.1-fold | 1.5-fold | 3.3 |
| pH 7.3 (stat phase) without DNA | <6.5 × 10−9 ± 8.0 × 10−10 | 0 | <0.005-fold | ∞ | n.a. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golz, J.C.; Stingl, K. “Take It or Leave It”—Factors Regulating Competence Development and DNA Uptake in Campylobacter jejuni. Int. J. Mol. Sci. 2021, 22, 10169. https://doi.org/10.3390/ijms221810169
Golz JC, Stingl K. “Take It or Leave It”—Factors Regulating Competence Development and DNA Uptake in Campylobacter jejuni. International Journal of Molecular Sciences. 2021; 22(18):10169. https://doi.org/10.3390/ijms221810169
Chicago/Turabian StyleGolz, Julia C., and Kerstin Stingl. 2021. "“Take It or Leave It”—Factors Regulating Competence Development and DNA Uptake in Campylobacter jejuni" International Journal of Molecular Sciences 22, no. 18: 10169. https://doi.org/10.3390/ijms221810169